











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
El venado de Coues (Odocoileus virginianus couesi (Coues & Yarrow, 1875) (Ramírez-Pulido et al., 2005) es una de las 14 subespecies de venado cola blanca en México (Ramírez-Pulido et al ., 2005; Villarreal, 1999). Su área de distribución abarca 30.4 % de la república mexicana y es de alto valor cinegético (Galindo-Leal y Weber, 1998). Se localiza desde Arizona, en los Estados Unidos de América, hasta Jalisco en México, a lo largo de la Sierra Madre Occidental (Ezcurra y Gallina, 1981). El estado de Aguascalientes forma parte de su territorio (Galindo-Leal y Weber, 1998), donde la caza furtiva estuvo a punto de extinguirlo durante la década de 1970, época en la que se registró una densidad de 0.65 venados km-2 (Medina-Flores y Medina-Torres, 1989).
Gracias a un programa de vigilancia, instituido en 1975 (Medina-Flores y Medina-Torres, 1989; Villalobos, 1989), sus poblaciones se recuperaron 20 años después, hasta alcanzar cifras de 2.00 venados km-2 (Kobelkowsky, 2000); lo anterior coincidió con el establecimiento de las primeras Unidades de Manejo para la Conservación de la Vida Silvestre (UMA), para su aprovechamiento cinegético.
A partir de 2008, la vigilancia se ha restringido a la Sierra Fría, por lo que en el resto del estado la caza furtiva ha continuado en áreas naturales como la Sierra del Laurel, con presencia de bosques templados de encino y vegetación subtropical, con manchones de áreas perturbadas de pastizal inducido (Medina-Torres, 2008; Medina-Torres et al ., 2008).
Desde 2006 la Comisión Nacional Forestal (Conafor) ha dado un fuerte impulso al establecimiento y operación de UMA extensivas y proyectos de ecoturismo en la Sierra del Laurel, situación que puede poner en peligro la estabilidad poblacional del venado nativo.
La falta de sistemas de seguimiento y evaluación de los estudios poblacionales que realizan los dueños de las UMA constituye un serio problema que impide el funcionamiento como instrumento de conservación, y propicia la sobreexplotación del venado y otras especies de vida silvestre (Urquiza, 2009). Ante este problema, Urquiza (2009) señala la necesidad de que "estos estudios sean realizados por la autoridad o por entes independientes, pues hay el potencial de un conflicto de intereses cuando se deja esta responsabilidad a los dueños de las UMA...". Si bien, ello plantea el dilema de que la autoridad ambiental sea juez y parte, debe considerarse que los técnicos contratados por los dueños de las UMA, pueden verse obligados a alterar los resultados de estudios poblacionales a entera satisfacción de sus empleadores, en consecuencia se sobreestiman las tasas de aprovechamiento. En el caso particular del venado cola blanca, la explicación de su densidad en respuesta a los atributos de su hábitat servirá para pronosticar la variación de su población en tiempo y espacio, si se pretende realizar su aprovechamiento, o bien si ya se está llevando a cabo.
En la Sierra del Laurel, se encontró que la pendiente y altitud del terreno, la distancia al agua, la altura media del estrato arbustivo, además de la densidad por hectárea de Ipomoea murucoides Roem. & Schult y Eysenhardtia polystachya Ortega Sarg. influyeron en la selección del hábitat por el venado de Coues ; es decir, su presencia o ausencia, con lo que se generó un mapa digital de la probabilidad de uso del hábitat por el venado (Medina-Torres et al., 2008). No obstante, aún es necesario analizar la relación entre la densidad de su población y su hábitat, así como determinar los atributos de este que afectan la variabilidad de su densidad. Con ese fin se plantearon los siguientes objetivos: estimar la densidad poblacional del venado cola blanca en la Sierra del Laurel, Aguascalientes; caracterizar el hábitat con base en sus atributos físicos y florístico-estructurales; y explicar la variabilidad de la densidad del venado en respuesta a los atributos del hábitat.
Materiales y Métodos
La investigación se realizó durante 2006 en la Sierra del Laurel, municipio Calvillo, al suroeste del estado de Aguascalientes (Medina-Torres et al., 2008) (Figura 1).
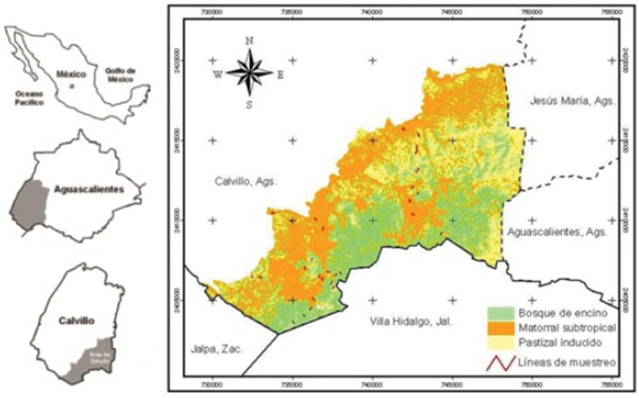
Tiene una superficie de 14 610 ha. Se ubica entre los 21°42' y 21°51' norte, y 102°47' y 102°35' oeste. Pertenece a la subprovincia fisiográfica Sierras y Valles Zacatecanos; tiene una topografía accidentada, con altitudes que varían de 1 700 a 2 760 m. Contiene rocas ígneas extrusivas del Cenozoico; en las partes altas predomina el tipo de suelo Litosol, con profundidades menores a 10 cm, limitado por rocas y tepetate. En las partes bajas sobresalen los suelos Feozem háplico, caracterizados por fertilidad moderada y una capa superficial obscura, suave y rica en materia orgánica y nutrientes (Medina-Torres, 2008; Inegi, 2009). Los cauces drenan hacia el sistema de los ríos Gil-Malpaso-Calvillo, del sistema hidrológico tributario de la subcuenca del río Juchipila, perteneciente a la región hidrológica 12.
En las partes altas, el clima corresponde al templado subhúmedo, con lluvias en verano (C (w)), mientras que hacia las partes bajas del Valle de Huajucar, el clima es semicálido semiseco (BS1h). En la zona oriental, hacia el valle de Aguascalientes, el clima es cálido semiseco (BS1k) (García, 1998; Inegi, 2009). La precipitación promedio anual es de 615 mm, de acuerdo al promedio histórico documentado entre 1960 y 2008 (Medina-Torres et al., 2008).
Se han identificado tres tipos de vegetación: bosque de encino, matorral subtropical y pastizal inducido. El aprovechamiento pecuario es la principal actividad económica, aunque existen algunas áreas agrícolas, y huertos de guayaba, hacia las partes bajas (Medina-Torres et al., 2008).
Densidad poblacional del Odocoileus virginianus couesi
Para obtener una estimación de la densidad poblacional conservadora y evitar sobreestimaciones, se utilizó la tasa de defecación de 25 grupos fecales de venado por día determinada por López-Téllez et al. (2007). Se comparó con estimaciones de trabajos similares realizados en México (Gallina, 1990; Mandujano y Gallina, 1995; Ortíz-Martínez et al., 2005), 15.2 (Zavala, 1992, citado por Ortíz-Martínez et al ., 2005) y 22.0 (Galindo-Leal y Weber, 1998).
El establecimiento de las parcelas (recuento inicial y limpieza subsiguiente) dio inicio en febrero y continuó hasta mayo de 2006, ya que sobre algunas de ellas se realizaron las mediciones de las variables del hábitat. Se tomó en cuenta un tiempo de depósito de 120 días, previo al inicio del establecimiento de las parcelas, lo anterior para cumplir con el supuesto básico del método de Eberhardt y Van Etten (1956): que solo serán contados los grupos fecales depositados después de la caída de las hojas. Al avanzar en el establecimiento de las parcelas, se sumaron los días transcurridos al tiempo de depósito, de tal manera que varío entre 120 y 219 días. La segunda revisión se efectuó durante los meses de junio y julio de 2006, y el tiempo de depósito de los grupos fecales acumulados fue de 55 a 122 días. Dado que las lluvias ese año se retrasaron hasta agosto, se consideró que no hubo pérdida importante de excretas por arrastre pluvial.
La densidad poblacional se estimó mediante el recuento de los grupos fecales en parcelas circulares de 9.3 m2 (Eberhardt y Van Etten, 1956), ya que dicho tamaño representa un equilibrio entre la varianza de muestreo (disminuye) y el sesgo de muestreo (incrementa), en relación con el incremento del tamaño de la parcela (Smith, 1968).
Se tomó como un grupo fecal todo hallazgo de al menos cinco excretas con características semejantes (Gallina, 1990), dispersas no más de 60 cm lineales. El muestreo base se hizo sobre líneas o transectos de 200 m cada uno, para un total de 30 líneas, y en cada una se colocaron seis parcelas, separadas 40 m una de otra para sumar 180 parcelas: 13 en bosque de encino, 10 en matorral subtropical y siete en pastizal inducido. Por parcela se verificó la existencia de grupos fecales en un radio de 1.72 m, a partir de su centro georreferenciado con un error de ubicación de tres metros. Se empleó un modelo de muestreo aleatorio estratificado (Gómez-Aguilar, 1979), para una precisión de 30 % y una confianza de 93.0 %, valores adecuados, dados los recursos humanos y materiales con los que se contó para la investigación. Cabe mencionar que durante la segunda revisión no se localizaron seis parcelas pertenecientes a dos líneas, debido a la pérdida de las marcas utilizadas, por lo que no fueron incluidas en el análisis estadístico.
Con el fin de comprobar si la densidad estimada de venados no varió entre las dos etapas de muestreo, y dado que no hay normalidad en los datos por tratarse de recuentos, se aplicó una prueba de rangos de Wilcoxon . La densidad poblacional se estimó con el modelo:
Donde:
D = Densidad de venados por km2
NP = 107 526.88 parcelas por km2
PG = Promedio de grupos fecales por parcela
TP = Tiempo de depósito de los grupos fecales
TD = Tasa de defecación (grupos fecales por venado por día) (Eberhardt y Van Etten, 1956)
Con base en que los grupos fecales tienen una distribución espacial agregada, se empleó para el cálculo del error estándar el modelo de una distribución binomial negativa (Ezcurra y Gallina, 1981), con la ecuación:

Donde:
x = Media muestral de grupos fecales por parcela
k = Coeficiente de la distribución binomial negativa
n = Número de muestras
El coeficiente se obtuvo mediante la expresión:
Donde:
Caracterización del hábitat
Las variables topográficas, de disponibilidad de agua y florísticoestructurales del hábitat fueron las de predicción en los modelos de regresión logística para modelar la selección de hábitat por el venado de Coues en la Sierra del Laurel (Medina-Torres et al., 2008), y sus valores se promediaron por transecto. Cada comunidad vegetal se caracterizó a partir de ellas.
Las metodologías y procedimientos para obtener las variables topográficas, de disponibilidad de agua, y florístico-estructurales del hábitat se describen a detalle en Medina-Torres et al. (2008).
A partir de los datos obtenidos para cada unidad de muestreo y con el programa Bio-Dap ® (Thomas y Clay, 2005) se estimaron la diversidad, la equidad y la dominancia de especies vegetales. El índice de diversidad de Shannon (H ') expresa la uniformidad de los valores de importancia a través de todos los taxa de plantas en la muestra, bajo el supuesto de que los individuos son seleccionados al azar y que todos los taxones están presentes en la muestra. Adquiere valores entre cero, cuando hay un solo taxon, y el logaritmo de S (número de especies), cuando todas las especies están representadas por el mismo número de individuos (Magurran, 1988). Este índice se obtuvo mediante la ecuación:
Donde:
pi = Proporción de individuos de la especie i respecto al total de individuos (es decir la abundancia relativa de la especie i ;
ni = Número de individuos de la especie i-esima
La equidad se calculó con el índice de Pielou (E), que mide la proporción de la diversidad observada con relación a la máxima diversidad esperada:
Donde:
Su valor va de 0 a 1, de tal forma que el máximo indica que todas las especies son igualmente abundantes (Magurran, 1988).
El predominio se determinó por medio del índice de Simpson (λ), que calcula la probabilidad de que dos individuos tomados al azar de una muestra sean de la misma especie. Este índice está fuertemente influido por la importancia de las especies más dominantes (Peet, 1974; Magurran, 1988). Su valor va de 0 a 1, de tal forma que 1 corresponde a situaciones en las que todas están igualmente representadas (Magurran, 1988):
Donde:
Un análisis multivariado, mediante la técnica del análisis de componentes principales (ACP), permitió reducir las 29 variables del hábitat y verificar los agrupamientos de las líneas por tipo de vegetación. Con ello, se definieron las que contribuyeron a explicar la mayor variación posible en un espacio de tres dimensiones, dado por los primeros tres componentes principales resultantes. Debido a que las variables se midieron en unidades diferentes, se utilizó la matriz de correlaciones (Johnson, 2000).
Relación entre la densidad de venados y los atributos del hábitat
Los atributos del hábitat que explican la densidad del venado, y dado que los recuentos no siguen una distribución normal, se aplicó un modelo lineal generalizado (Pozo-Montuy, 2006) para una distribución binomial negativa con función de enlace logarítmica, cuya variable de respuesta fue el número redondeado de venados km-2, los factores correspondieron a los tipos de vegetación y las covariables a los atributos del hábitat. Este procedimiento se usó para probar la hipótesis de que la densidad de venados no varió entre los tres tipos de vegetación bajo estudio. Los parámetros del modelo se estimaron por el método de Fisher ; y los efectos del factor y covariables del modelo se evaluaron mediante el estadístico Chi cuadrado de Wald . Se analizaron contrastes por parejas, para probar la hipótesis de que los tipos de vegetación no difirieron en la densidad de venado, a un nivel de significancia p = 0.05. La proporción de la variación total de la densidad de venado explicada por el tipo de hábitat, se determinó a partir de la comparación de la devianza del modelo nulo con la D2 = (devianza del modelo nulo - devianza residual) / devianza del modelo nulo × 100.
Se inició con un modelo que incluyó el factor (tipos de vegetación) y todas las covariables, se repitió excluyendo del análisis aquellas no significativas (p > 0.05), hasta obtener el modelo más parsimonioso; es decir, aquel que tuvo el menor número de variables significativas. El criterio de información bayesiano (CIB) permitió seleccionar el mejor modelo, fue preferible el valor más bajo.
Resultados
Grupos fecales por parcela
Se contaron 91 grupos fecales en 96 de 180 unidades de muestreo, de los cuales 63 se registraron durante el establecimiento de las parcelas y 28 en el segundo recuento. La media de grupos fue de 0.35 ± 0.09 y 0.16 ± 0.04, respectivamente. La media total fue de 0.33 ± 0.06 grupos por parcela.
Densidad de venados
El intervalo de confianza a 95 % para la densidad de venados estuvo entre 5.0 y 11.6 venados por km2, con una media estimada estratificada de 8.3 para el área de estudio. La densidad no varió significativamente entre ambas etapas de muestreo, de acuerdo con la prueba de rangos de Wilcoxon (Z = -0.581, p = 0.561), por lo que solo se consideró el promedio de la densidad de venados.
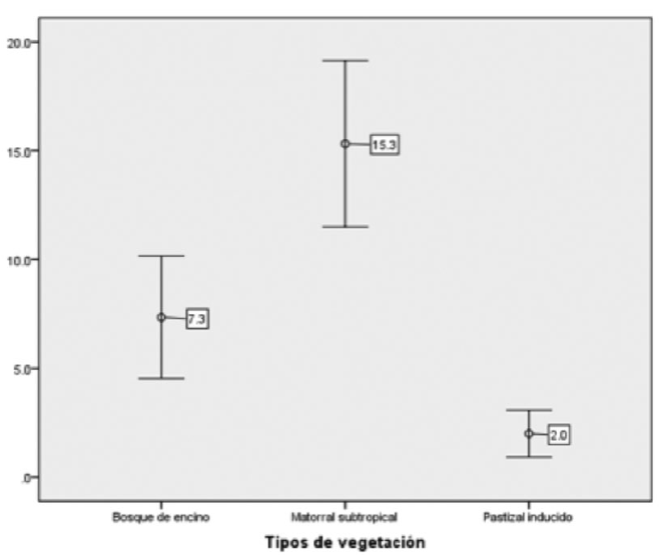
El modelo lineal generalizado demostró diferencias altamente significativas entre los tres hábitats (x2 Wald = 77.642, gl = 2, p = 0.000). El análisis de contrastes confirmó que la densidad varió significativamente entre los tipos de vegetación (Cuadro 1). La mayor densidad se obtuvo en el matorral subtropical, seguida del bosque de Quercus y del pastizal inducido (Figura 2).
Cuadro 1 Contrastes de la diferencia de medias del número promedio de densidad de venado cola blanca por tipo de vegetación.

Caracterización del hábitat
Los tres primeros componentes principales explicaron 66.8 % de la variación total. El primer componente (CP 1) representó 37.1 % del total, y las variables con mayor carga positiva en sus coeficientes fueron la cobertura del dosel arbóreo, la densidad de árboles, la altitud del sitio, el área basal y la altura promedio del arbolado; atributos característicos de estratos arbóreos bien desarrollados del bosque de Quercus . En contraste, los coeficientes con mayor carga negativa para este componente resultaron la cobertura del dosel arbustivo, la densidad de arbustivas, la densidad de árboles y arbustivas por sitio y la densidad de I. murucoides ; es decir, atributos representativos del matorral subtropical.
El segundo componente principal (CP 2) constituyó 20.4 % de la variación total. Las variables con más peso en los coeficientes positivos fueron el índice de Shannon (H' ) y la riqueza florística total, en tanto que la variable con mayor peso negativo fue el índice de predominio de Simpson (λ). El tercer componente principal (CP 3) explicó 9.3 % de la variación total, y en el extremo positivo la variable con mayor peso fue el porcentaje de suelo cubierto con mantillo, mientras que en el extremo negativo correspondió a la rocosidad del suelo (Figura 3).

Relación densidad de venados y atributos del hábitat
Al excluir las variables no significativas del análisis inicial, se generó el modelo más parsimonioso y con mejor ajuste (CIB = 562.24), en el cual siete variables resultaron altamente significativas (p<0.01), explicó poco más de 45 % de la variabilidad de la densidad de venados (D2 = 45.82 %; Cuadro 2). El modelo corresponde a la ecuación 1:
Donde:
DV = Densidad de venados km2 -1
βi = Parámetro de la variable
x1 = Tipo de vegetación
x9 = Índice de equidad de Pielou
x10 = Índice de predominio de Simpson
x13 = Densidad de árboles de I. murucoides por ha
x18 = Riqueza florística del estrato arbóreo
x24 = Altura promedio del estrato arbustivo
x28 = Porcentaje de suelo desnudo
εi = Error
Cuadro 2 Coeficientes de las variables explicativas de la densidad del venado cola blanca en la Sierra del Laurel, Aguascalientes.

Significancia: *** = < 0.001; ** = 0.001; * = 0.01; ns = 0.05.
Devianza del modelo nulo = 836.87, con 179 grados de libertad
Devianza residual = 764.93, con 177 grados de libertad
Criterio de información bayesiano = 562.24
Discusión
Densidad de población del venado
La estimación poblacional comparada con las obtenidas por otros autores en hábitats semejantes a la Sierra del Laurel sugieren que la densidad del venado tiende a ser mayor en lugares con vegetación caducifolia, en contraste con bosques templados, a excepción de La Michilía, Durango (Gallina,1990); aunque de acuerdo con Galindo-Leal y Weber (1998) en esa misma región se registraron densidades de venado más bajas en años posteriores (Cuadro 3).
Cuadro 3 Comparativo de las estimaciones de densidad de venado cola blanca obtenidas por otros autores con las obtenidas en el presente trabajo.
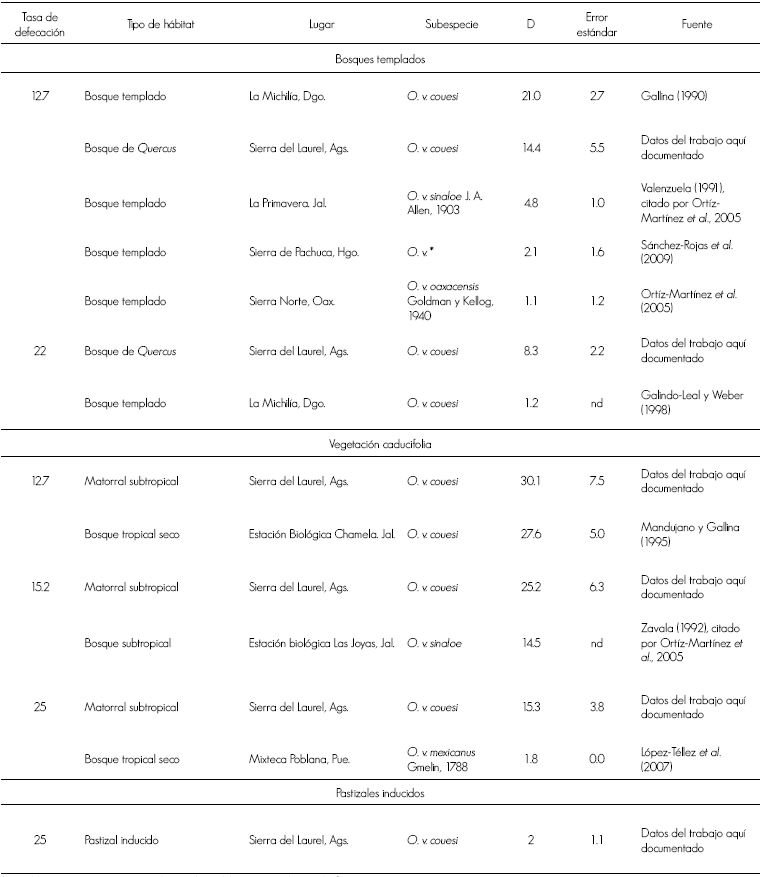
* = Posiblemente O. v. mexicanus ; nd = No disponible; D = Venados por km2.
Otra consideración importante es la aleatorización de las unidades de muestreo, pues si esta condición no se garantiza puede dar lugar a sesgos que ponen en duda o invalidan los resultados; situación que ha originado, incluso, a serias controversias entre investigadores (Galindo-Leal, 1992; Gallina y Ezcurra, 1992).
En el caso de este trabajo, con el establecimiento de las líneas de muestreo se garantiza la validez de los resultados obtenidos. Una ventaja por considerar, es que durante los estudios dasonómicos en los inventarios forestales, es posible obtener datos de las excretas del venado, y, con ello, realizar análisis de selección de hábitat.
Caracterización del hábitat
La utilidad del análisis de componentes principales como procedimiento preliminar de revisión de datos multivariados del hábitat, además de verificar la clasificación de las unidades muestrales, y de identificar datos anómalos permite una primera aproximación a priori , para detectar de manera cualitativa posibles asociaciones entre los agrupamientos configurados (hábitats) y sus respectivas estimaciones de la densidad poblacional del venado cola blanca, en un espacio bi o tri-dimensional de las unidades muestrales, ya sea que se utilicen los dos (Ortíz-Martínez et al., 2005; López-Téllez et al., 2007) o tres primeros componentes principales, es más común el primer caso.
En el presente estudio se observó que la variabilidad de los atributos del hábitat se explicaron por un reducido número de variables relacionadas con los estratos arbóreo, arbustivo y herbáceo, además del grado de equidad en cada comunidad vegetal. También, se determinó qué porcentaje de la variabilidad total del hábitat responde a los primeros tres componentes principales (66.8 %), porcentaje muy cercano al que López-Téllez et al. (2007) registraron en un bosque tropical seco de la Mixteca Poblana (67 %), y superior a lo citado por Ortíz-Martínez et al. (2005) en un bosque templado de Oaxaca (64 %); y por Sánchez-Rojas et al. (2009) en un hábitat similar en la Sierra de Pachuca, Hidalgo (58.6 %).
Relación densidad de venados y atributos del hábitat
Aun cuando algunos autores han señalado la relación aparente entre los agrupamientos generados por un ACP y las densidades de venados registradas en ellos (Ortíz-Martínez et al., 2005; López-Téllez et al., 2007), no es evidencia concluyente para propósitos de gestión y la estadística paramétrica no genera inferencias confiables. Esta investigación demostró que los modelos lineales generalizados ofrecen una solución razonable para entender la variación en la densidad de venados como respuesta a los atributos de su hábitat. Lo anterior es de particular importancia, cuando la densidad poblacional del venado se estima mediante técnicas indirectas, como el recuento de grupos fecales en parcelas, que por su naturaleza no tienen una distribución normal, sino que siguen una distribución agregada binomial negativa (Ezcurra y Gallina, 1981), lo que impide el uso de técnicas estadísticas paramétricas convencionales.
El método utilizado generó un modelo parsimonioso, con solo siete variables de un total de 29 considerados en un principio. Es importante resaltar que dos de las tres variables florístico-estructurales que explicaron la selección del hábitat por el venado en la Sierra del Laurel: número de árboles de I. murucoides ha-1 y altura promedio del estrato arbustivo (Medina-Torres et al., 2008) explicaron, además, la variación en su densidad.
El análisis de los coeficientes de las variables (Cuadro 3) muestra que de los tres tipos de vegetación estudiados, solamente, el pastizal inducido no contribuyó a elucidar la densidad del venado. Asimismo, destaca que el intercepto, que resume el efecto del bosque de encino sobre la densidad del venado, es significativo y negativo, lo que confirma que la densidad en este hábitat es menor en relación al matorral subtropical, resultado semejante a lo citado por otros autores, en bosques templados (Valenzuela, 1991, citado por Ortíz-Martínez et al., 2005; Galindo-Leal y Weber ,1998; Ortíz-Martínez et al ., 2005), o en vegetación caducifolia (Zavala, 1992, citado por Ortíz-Martínez et al ., 2005; Mandujano y Gallina, 1995) (Cuadro 3).
Los coeficientes de las otras variables fueron positivos y altamente significativos, lo cual indica que a medida que dichos atributos se incrementen, la densidad del venado también lo hará. Cabe señalar que los valores más altos son característicos del matorral subtropical y puede considerarse que, en conjunto, explicaron por qué la densidad del venado cola blanca de la Sierra del Laurel es mayor en ese tipo de vegetación.
Por ejemplo, valores altos del índice de equidad corresponderán a sitios donde la repartición vegetal observada estará cercana a la máxima esperada, lo cual implica que el venado será más abundante en hábitats con especies vegetales igualmente representadas. Esto contrasta con el índice de predominio de Simpson , ya que valores altos corresponden a sitios con pocas especies predominantes, que puede ser reflejo del aparente predominio de I. murucoides en los sitios con más densidad de venado. Aun cuando el coeficiente positivo de la densidad de I. murucoides fue uno de los valores más bajos (un posible efecto menos intenso en relación a las otras variables del modelo sobre la densidad del venado), es importante resaltar que su aparición entre las variables explicativas sugiere una dependencia del venado por la densidad de este árbol, tan característico del matorral subtropical, e incluso es posible que su baja densidad en sitios con alta riqueza y diversidad de especies vegetales sea un factor limitativo para la presencia de venado. Se ha documentado que la flor de I. murucoides , tiene 76.25 % de humedad y está disponible durante la floración, entre diciembre a marzo, por lo que sirve como fuente de agua en entornos similares al matorral subtropical de la Sierra del Laurel (Villarreal y Marín-Fuentes, 2005). Es posible que en dicha Sierra, donde son frecuentes las sequías recurrentes y la falta de corrientes de agua permanentes, el venado cubra sus necesidades del vital líquido mediante el consumo de la flor de I. murucoides (Medina-Torres et al., 2008).
Otra de las variables del sotobosque que también contribuyó a explicar la densidad del venado fue el porcentaje de suelo desnudo, pero al parecer no hay un argumento ecológico que respalde esa aparente predilección por suelos desprovistos de vegetación y mantillo. De hecho, puede interpretarse como un sesgo causado por la fácil detección de las excretas en suelos desnudos, por lo que su efecto sobre la densidad del venado debe tomarse con reserva.
Dos atributos más del matorral subtropical fueron altamente significativos en el modelo lineal generalizado: la riqueza florística del estrato arbóreo y la altura promedio del estrato arbustivo, que se relacionan con la disponibilidad de alimento, la cobertura térmica y de escape. Dado que la Sierra de Laurel se distingue por su clima extremoso y sequías prolongadas (Medina-Torres, 2008), el venado requiere de vegetación particularmente densa que le brinde alimento y protección contra las inclemencias térmicas, así como de refugio o vías de escape contra sus depredadores (Galindo-Leal y Weber, 1998; Bello y Equihua, 2001; Ortíz-Martínez et al., 2005). Medina-Torres et al . (2008) consignan que la altura media del estrato arbustivo para una probabilidad de uso superior a 0.5 no debe ser inferior a 2 m.
Ya se ha señalado que la aparente selección del matorral subtropical por el venado pudiese más bien responder a presiones y amenazas a su población. Pese a que no fueron analizados a profundidad, se identificaron otros factores antrópicos que amenazan al venado. Es evidente que el pastizal inducido es producto del desmonte y el sobrepastoreo; su distribución espacial sugiere un proceso de fragmentación preocupante, lo que debe ser objeto de nuevas investigaciones. El matorral subtropical fue el hábitat "seleccionado" por el venado; sin embargo, tal elección, posiblemente es más una respuesta a las amenazas (fragmentación de su hábitat, sobrepastoreo, caza furtiva y presencia de excursionistas), que a la satisfacción de sus necesidades básicas (Medina-Torres, 2008). La caza furtiva es un fenómeno que se ha incrementado en los años posteriores a la realización de esta investigación, y dado lo accidentado del terreno, los cazadores con frecuencia incursionan a pie o a caballo, internándose en la sierra por las mañanas y por las tardes, y con ello obligan a los venados a replegarse hacia los matorrales más densos.
Otros factores que deben considerarse en estudios posteriores, para explicar la densidad del venado en el matorral subtropical son los relacionados con la demografía de la especie. Una mayor proporción de hembras en la población de venados redunda en un incremento en la producción de cervatos, con la consecuente sobrecarga del hábitat. Es frecuente que los migrantes radicados en Estados Unidos de América, quienes arriban durante diciembre y enero al municipio Calvillo a visitar a sus familias, salgan a cazar venados machos.
Sin embargo, aun el matorral subtropical como hábitat que aparentemente satisface los requerimientos del venado, a su vez enfrenta amenazas. Siqueiros-Delgado et al. (2006) mencionan que esa vegetación en la Sierra del Laurel es uno de los tipos más afectados por el desarrollo agropecuario, pues su extensión ha sido mermada por cambios de uso de la tierra para el establecimiento de cultivos de guayaba o de agave azul, o para la ganadería. Se ha documentado que los ganaderos eliminan el matorral subtropical y afectan la calidad del hábitat del venado. González-Alaníz (1997) considera que I. murucoides es una especie tóxica para el ganado, por lo que los productores la erradican con herbicidas. Durante octubre de 2006, la Procuraduría Estatal de Protección al Ambiente del estado de Aguascalientes (Proespa) documentó en el municipio Calvillo que en más de 100 ha se había talado hasta 83 % de los árboles de I. murucoides y eliminado a todos los arbustos de la especie.
Conclusiones
La densidad de venados en la Sierra del Laurel durante la época seca de 2006 varió significativamente entre los tres tipos de vegetación existentes, fue mayor en el matorral subtropical, seguida del bosque de Quercus y del pastizal inducido. Las variables que contribuyeron a explicar la variabilidad de la densidad del venado fueron el tipo de vegetación, el índice de equidad de Pielou , el índice de predominio de Simpson , la densidad de árboles de I. murucoides , la riqueza florística del estrato arbóreo y la altura promedia del estrato arbustivo.
La fragmentación del hábitat por la apertura de tierras a nuevos cultivos, el sobrepastoreo, la caza furtiva, la afluencia de paseantes y la proliferación de infraestructura para proyectos de ecoturismo no se descartan como factores de presión sobre el venado, por lo que se requieren de más estudios para confirmar o descartar que tales factores influyan sobre la densidad de la especie en el matorral subtropical.
Asimismo, se requiere de mayor profundidad en estudios demográficos que permitan conocer la relación de sexos y clases de edad, para evaluar el éxito reproductivo de la especie, lo que pudiera realizarse mediante foto-trampeo.
Conflicto de intereses
Los autores declaran no tener conflicto de intereses.
Contribución para autor
Salvador Martín Medina Torres: selección de unidades de muestreo en campo, documentación de antecedentes, elaboración del manuscrito; Edmundo García Moya: análisis de la relación hábitat-densidad del venado, estructuración y revisión del manuscrito; Marcelo Márquez Olivas: seguimiento del muestreo poblacional, estructuración y revisión del manuscrito; Angélica Romero Manzanares: obtención de indicadores de riqueza, diversidad, equidad y dominancia vegetal y revisión del manuscrito; Humberto Vaquera Huerta: análisis estadístico multivarido y modelo lineal generalizado; Mario Martínez Menes: construcción del sistema de información geográfica y geo-referenciación del marco de muestreo.

