










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista mexicana de ciencias forestales
Print version ISSN 2007-1132
Rev. mex. de cienc. forestales vol.5 n.26 México Nov./Dec. 2014
Artículos
Características anatómicas de la madera de Bursera lancifolia (Schltdl.) Engl. con potencial dendrocronológico
Anatomic characteristics of the wood of Bursera lancifolia (Schltdl.) Engl. with dendrocronological potential
Omar Durán Guerra1, Alejandra Quintanar Isaías1, José Villanueva Díaz2, Ana Teresa Jaramillo-Pérez1 y Julián Cerano Paredes2
1 Universidad Autónoma Metropolitana Unidad Iztapalapa. Correo–e: guerra.duran.omar@gmail.com.
2 Centro Nacional de Investigación Disciplinaria Relación Agua, Suelo, Planta, Atmósfera. Instituto Nacional de Investigaciones Forestales Agrícolas y Pecuarias.
Fecha de recepción: 30 de julio de 2013;
Fecha de aceptación: 6 de agosto de 2014.
Resumen
Algunas especies arbóreas tropicales forman anillos de crecimiento marcados por fibras, bandas de parénquima marginal o terminal, o por el calibre de vasos de madera temprana. Características que se utilizan para interpretar ritmos y tasas de crecimiento, así como edades. En el presente trabajo se hace una evaluación de las propiedades anatómicas de la madera de Bursera lancifolia con valor dendrocronológico. Se usaron 10 núcleos extraídos de tres árboles para caracterizar los tipos celulares que marcan el anillo y para realizar su estudio anatómico. Los núcleos se fecharon en mayo del año 2010, con una correlación de 0.339, mediante el programa COFECHA, y una serie cronológica de 69 años, con una sensibilidad promedio de 0.410; dicha cronología se correlacionó con los datos meteorológicos de precipitación de la zona. Los resultados muestran que Bursera lancifolia es sensible a marcar sus anillos de crecimiento con fibras radialmente comprimidas, septadas y funcionalmente almacenadoras. La madera presenta porosidad difusa con escaso parénquima axial y radios multiseriados. Los parámetros estadísticos indican que B. lancifolia es climáticamente sensible y muestra un comportamiento similar al de las coníferas que se han utilizado para la reconstrucción del clima del centro de México. Se recomienda el uso de la madera de esta especie para estudios dendrocronológicos.
Palabras clave: Anillos de crecimiento, COFECHA, dendrocronológico, especies maderables tropicales, núcleos, xilema secundario.
Abstract
Certain tropical tree species form growth rings marked by fibers, strips of marginal or terminal parenchyma or by the caliber of early wood vessels. These characteristics are used for interpreting growth rates and ages. This paper assesses those anatomical properties of the wood of Bursera lancifolia that have dendrochronological value. 10 cores were obtained from three trees in order to characterize the cell types marking the ring and to carry out their anatomical study. The cores were dated May 2010, with a correlation of 0.339, using the COFECHA software, and a chronological series of 69 years was dated using an average sensitivity of 0.410; this chronology established a correlation with the meteorological precipitation data for the area. The results show that Bursera lancifolia is sensitive to marking growth rings with radially compressed, septated and functionally storing fibers. The wood has a diffuse porosity with a scarce axial parenchyma and multiseriate radii. The statistical parameters show that B. lancifolia is sensitive to climate and has a similar behavior to that of the conifers utilized to reconstruct the climate of central Mexico. The use of the wood of this species is recommended for dendrochronological studies.
Key words: Growth rings, COFECHA, dendrochronological, tropical timber species, cores, secondary xylem.
Introducción
La dendrocronología es una disciplina que ha contribuido a la solución de problemas de índole ecológico, hidrológico, climático o arqueológico. Se fundamenta en la confirmación de la periodicidad del crecimiento del xilema secundario de especies maderables, correspondientes a pulsos anuales, lo que permite definir edades, tiempos históricos o explicar fenómenos de cambio climático (Villanueva, 2000).
En el ámbito mundial se han estudiado pocos taxa que habitan en regiones tropicales y subtropicales (Boninsegna y Villalba, 1996; Villalba, 2000); sin embargo, se ha demostrado que forman anillos de crecimiento, marcados por fibras, bandas de parénquima marginal o terminal, o por el calibre de vasos de madera temprana. Estas características se utilizan para interpretar ritmos y tasas de crecimiento, así como edades en algunos árboles tropicales (Jacoby, 1989; Détienne, 1989; Brienen y Zuidema, 2006; Lisi et al., 2008; Borman y Berlin, 1981; Vetter y Botosso, 1989; Worbes, 1989; Boninsegna et al., 1989). A partir de esa información es posible hacer una prospección de los ecosistemas tropicales en relación con el clima, la hidrología y la fenología.
En Brasil, Venezuela, Bolivia, Perú, India, y la isla de Java se han caracterizado anatómicamente anillos de crecimiento (Devall et al., 1995; Schweinggruber, 1996; Zumaeta, 2008). En particular en Brasil (Détienne y Mariaux, 1977; Vetter y Botosso, 1989; Worbes, 1989; Détienne, 1989; Worbes, 1995; Mattos, 1999; Tomazello y Cardoso, 1999; Tomazello et al., 2001); en Bolivia y en Perú se han reconocido especies tropicales formadoras de crecimiento periódico (Brienem y Zuidemman, 2003; Brienem y Zuidemman, 2005; Rosero, 2009), el cual es un requisito para la selección de especies útiles en la elaboración de series de tiempo para obtener información dendrocronológica. Koriba (1958) describe que en Malasia 15 % de los taxa presentaron un crecimiento continuo, mientras que el resto evidenció algún tipo de crecimiento periódico; en los bosques lluviosos del Amazonas, 43 % de los árboles mostraron anillos de crecimiento (Alvim, 1964) y solo 25 % en la India (Chowdhury, 1964), con un porcentaje similar en el trópico húmedo de Argentina.
En el centro y sur de México existe un reducido número de estudios en dendrocronología tropical, debido a la idea de que las especies tropicales no forman anillos anuales de crecimiento (Stahle et al., 1998), como consecuencia en el país hay pocas series de tiempo dendrocronológicas en zonas con bosques tropicales secos, por lo que aún se desconoce el potencial dendrocronológico de sus taxa (Roig et al., 2012).
En el país, la mayoría de las investigaciones se han realizado en clima templado y frio (Villanueva et al., 2010) y algunas en el trópico. En la Península de Yucatán, Roig et al. (2005, 2012) estudiaron 52 especies, e indican que el porcentaje de plantas con anillos distinguibles, con potencial dendrocronológico, es igual o mayor a los registrados para otras zonas tropicales húmedas.
De acuerdo con Mauseth, (1988) y Raven (2005) el meristemo responsable de la formación de anillos de crecimiento es particularmente sensible a factores de estacionalidad o a factores fenológicos, entre especies, por lo que su actividad puede ser temporalmente asincrónica (Coster, 1927; Coster, 1928; Détienne y Mariaux, 1977; Bormann y Berlyn, 1981; Baas y Vetter, 1989; Jacoby, 1989; Lisi et al., 2008). Los trabajos centrados en la actividad cambial han ayudado a determinar si el crecimiento es continuo o discontinuo en los árboles tropicales, lo que permite seleccionar especies maderables con crecimiento periódico.
Los bosques tropicales caducifolios están ampliamente distribuidos en México. En este tipo de comunidades el género Bursera es un elemento florístico característico; se presentan como individuos de buena talla, la que varía con la disponibilidad del agua. Su distribución se restringe al continente americano, en particular, a la mitad septentrional de su porción intertropical; comprende desde los extremos suroeste y sureste de los Estados Unidos de América, hasta el norte de Perú y de Brasil, las Antillas y las Galápagos (Rzedowski, 2004).
Bursera simaruba (L.) Sarg. en Costa Rica (Enquist y Leffler, 2001) y en un bosque seco de Yucatán (Valdez-Hernández et al., 2010) muestra anillos marcados por fibras, lo que representa un potencial para la ejecución de estudios dendrocronológicos. En B. graveolens (Kunth) Triana & Planch. de Perú, y B. tomentosa (Jacq.) Triana & Planch. procedente de bosques secos en Venezuela (Castillo et al., 2005) se observaron fibras marcando anillos de crecimiento.
En México se ha registrado la formación de anillos por fibras radialmente comprimidas y de mayor densidad, pero difícilmente visibles solo para Bursera simaruba (Roig et al., 2012); esto contrasta con lo señalado por Barajas (1997) y De la Paz y Corral (1980), quienes señalan que la porosidad es difusa sin describir ninguna particularidad asociada a un comportamiento periódico para las fibras.
Bursera lancifolia (Schltdl.) Engl. se localiza en Guerrero, Oaxaca, Querétaro, Morelos y Puebla (Rzedowski, 1992). En los bosques caducifolios del estado de Morelos, los árboles alcanzan hasta 20 m de alto y fustes de 40 cm de diámetro, lo que planteó considerar una evaluación de las características anatómicas del xilema secundario con potencial dendrocronológico.
Materiales y Métodos
El sitio de estudio se caracteriza por presentar un bosque tropical caducifolio (Rzedowski, 1988), en la localidad de Valle de Vázquez, municipio Tlalquiltenango, Morelos. La recolecta del material se realizó en el cerro Puente de Tierra, en una ladera cuyas coordenadas son 18°30'8.1" latitud norte y 99°05'10.9" longitud oeste, a una altitud de 1 019 m.
Para el estudio anatómico y dendrocronológico se seleccionaron, en mayo del año 2010, tres árboles sanos, con el fuste recto, diámetro de 17 hasta 26 cm y una altura de 11 a 17 m. A cada individuo se le extrajeron de tres a cuatro núcleos, para representar la circunferencia, la cual dependió de la posición. Las muestras se obtuvieron con un taladro Pressler, a un DAP de 1.30 m,
Los núcleos se guardaron en bolsas con agua para mantenerlos hidratados. Se montaron en bases de madera acanaladas para su manipulación. Las superficies transversales, tangenciales y radiales se pulieron con una navaja de un filo y, a mano alzada, se realizaron cortes de las tres secciones, las que se fijaron con FAA (formol, agua, ácido acético, alcohol). Posteriormente, se tiñeron con lugol, para resaltar los contenidos ergásticos y las paredes secundarias. Se hicieron montajes semipermanentes para manipular y no perder la tinción. La descripción de los caracteres anatómicos microscópicos de la madera se hizo de acuerdo con los criterios del Comité de la IAWA (1989).
Los núcleos de crecimiento se trataron de acuerdo con técnicas dendrocronológicas para procesarlos en el Laboratorio de Dendrocronología del Centro Nacional de Investigación Disciplinaria Relación Agua, Suelo, Planta, Atmósfera, del Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (CENID- RASPA, INIFAP). Para la medición de la anchura, conteo y fechado anual se usó el sistema trilocular estereoscópico Velmex y las técnicas dendrocronológicas convencionales (Stokes y Smiley, 1968). El fechado se verificó con el programa COFECHA (Holmes, 1983; Grissino–Mayer, 2001), mientras que las tendencias biológicas y geométricas no relacionadas con el clima, se removieron mediante el proceso de estandarización en el programa ARSTAN (Cook y Holmes, 1984).
La respuesta climática de la especie se determinó con un análisis de función de respuesta, entre los índices dendrocronológicos de anillo total y los datos climáticos de la estación "Nexpa" del Servicio Meteorológico Nacional, para ello se usaron los registros de los años 1976 a 2001. La estación climática se eligió en función a la cercanía del sitio de recolecta y la revisión previa de la extensión y la calidad de los datos climáticos disponibles.
La confirmación de la anualidad de los anillos de crecimiento se realizó comparando los valores de los índices de anchura de anillo y la precipitación acumulada anual, lo que permitió determinar si existía asociación significativa entre estos. La información se corroboró con el codatado de los crecimientos anuales y se verificó con el programa COFECHA. El tamaño de muestra utilizado fue de 10 núcleos procedentes de los tres árboles seleccionados.
Resultados
Descripción microscópica de la madera
La madera de Bursera lancifolia presenta anillos de crecimiento distinguibles, formados por una o dos hileras de fibras radialmente comprimidas (Figura 1A, B y E). La porosidad es difusa, con poros solitarios (72.60 %), múltiples radiales de dos a cuatro (21.29 %), múltiples tangenciales de dos a cuatro vasos (5.64 %) y algunos diagonales de dos (0.43 %) (Figura 1A, B y E). Los elementos de vaso tienen placas perforadas, simples y diámetros medianos (100-200 µm). Estos son numerosos (20-40/mm2), con un índice de vulnerabilidad de Carlquist de 4.8. La distribución de los vasos es completamente al azar, por lo que la porosidad no destaca como un carácter que marca los anillos de crecimiento. Las punteaduras intervasculares son areoladas, alternas, de pequeñas a medianas (4-10 µm) (Figura 1D). Las punteaduras de los campos de cruzamiento son de bordes reducidos a aparentemente simples, de medianas a grandes (8–12 µm). Las de mayor tamaño con el eje horizontal y las medianas de forma oval. Destaca que el tamaño de las punteaduras revela una relación de trasporte radio-vaso poco restringido. Los vasos forman tílides con almidones en la albura, y hacia el duramen con gomas y resina. Las tílides aparecen en vasos cavitados.

Figura 1. A, B y E. Secciones transversales mostrando anillos marcados por fibras (af), gomas en vasos (go). C. Sección
tangencial mostrando radios multiseriados (r), canales gomíferos (cg). D. Sección radial mostrando punteaduras
campo de cruzamiento (pc), granos de almidón (a).
Haga clic para agrandar
El parénquima axial paratraqueal es escaso, con cordones uniseriados de tres a siete células. El parénquima de radio es de tipo uniseriado y multiseriado (de 3 a 5 series); presenta canales gomíferos (Figura 1C). El cuerpo está formado por células procumbentes; y por cuadradas en sus extremos, con cristales romboidales. Las fibras son septadas de paredes delgadas con punteaduras simples y en la albura presenta gran cantidad de granos de almidón (Figura 1D).
Análisis dendrocronológico
El análisis y la verificación de COFECHA reveló que los anillos anuales, considerados como potenciales, confirmaron el valor mínimo de significancia (r>0.3281, p<0.01). Resultados que muestran un total de anillos en todas las series de 523, con una serie de correlación de 0.339, una sensibilidad media promedio de 0.410, que abarca una serie maestra de 1940-2009.
Adicionalmente, siete de las 10 muestras superaron el valor de significancia para considerarse adecuadamente fechadas (Cuadro 2). La normalización de la series de anillo total realizada con el programa ARSTAN generó una serie de crecimiento de anillo total de 69 años (1940-2009).
Cuadro 2. Valores de correlación de cada una de las series por segmento de 25 años, derivadas del programa COFECHA.

Haga clic para agrandar
La serie dendrocronológica de anillo total para Bursera lancifolia respondió a la precipitación estacional acumulada para los meses enero-mayo en el período 1976 a 2001 (figuras 2 y 3).
La correlación entre ambas variables (r= 0.49), aunque es inferior a la observada entre variables climáticas y series dendrocronológicas de especies en bosques templados, que algunas veces superan valores de correlación de 0.8, indica que la precipitación es el factor que influye significativamente en el desarrollo de Bursera lancifolia.
Discusión
Bursera lancifolia es caducifolia y presenta una corteza de anchura considerable, lo que revela una estrategia para aislar los tejidos secundarios y evitar la pérdida del agua. Adicionalmente, muestra un sistema radicular muy profundo, que en ocasiones alcanza el manto freático (Rzedowski, 1988). Esto sugiere que tiene la capacidad para explorar fuentes de agua profundas, pero en condiciones desfavorables de baja disponibilidad y atmósferas desecantes recurre a la pérdida de hojas, lo que podría impactar la actividad del cambium vascular y, en consecuencia, al diámetro de los vasos y las fibras, situación que favorecería la formación de anillos de crecimiento distinguibles.
Roig et al. (2012) señalan que esta característica puede considerarse como un rasgo potencial dendrocronológico. En la presente investigación se demostró que los anillos de crecimiento de Bursera lancifolia, se forman por fibras radialmente comprimidas. El análisis dendrocronológico confirmó que los anillos anuales fechados y calculados con el programa COFECHA, y una vez verificadas las correlaciones entre ellos y la precipitación con el programa ARSTAN sugieren una potencialidad para reconstruir una cronología de hasta 69 años. En esa correlación, la formación de los anillos se explica en 49 % por la precipitación (figuras 2 y 3).

Figura 2. Función de respuesta entre los índices de ancho de anillo de Bursera lancifolia (Schltdl.) Engl. y la precipitación
acumulada estacional del período enero-mayo (1976 - 2001).
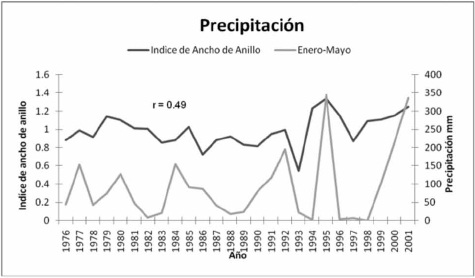
Figura 3. Asociación entre el índice de ancho de anillo total en Bursera lancifolia (Schltdl.) Engl. y precipitación estacional
acumulada del periodo enero-mayo (1976 – 2001).
Por otro lado, se destaca que en algunos núcleos, hacia el margen de la madera tardía, se aprecian vasos de diámetro reducido que aluden una respuesta tardía a la pérdida de hojas (Figura 1E). En general, prevalece la tendencia a la porosidad difusa y se descarta la semicircular.
La porosidad difusa es una estrategia que sugiere una dinámica de abasto regular durante el pulso anual de crecimiento. La distribución al azar y el arreglo de los vasos en solitarios, múltiples radiales, tangenciales y algunos diagonales a lo ancho del anillo provee un sistema accesorio para regular el impacto de la cavitación.
El tamaño de los vasos corresponde a especies de regiones mesófitas, más que del trópico seco; lo que se confirma por el valor de los índices de vulnerabilidad de Carlquist (Carlquist y Hoekman, 1985). Esto insinúa un abasto regular de agua del subsuelo en esta zona geográfica y la poca tolerancia a la atmósfera desecante, es lo que desencadena la caducidad.
Un diagnóstico de carencia de abastecimiento de agua se refleja en la formación de tílides en los vasos que se cavitan cuando el líquido se despresuriza, debido a las condiciones de la atmósfera previamente descritas (Quintanar et al., 2009). Lo que explica los resultados de la correlación entre anillos y precipitación y por qué esta especie presenta porosidad difusa y marca sus anillos solo con fibras. Al respecto, se observa que la respuesta climática de la serie de crecimiento fue al período estacional enero-mayo, y se detectaron respuestas similares en coníferas del norte, centro de México y Tlaxcala (Cardoza, 2010; Cerano et al., 2009). En este sentido, la etapa de crecimiento radial de la especie correspondería al período de invierno-primavera, en el que el agua es almacenada en el subsuelo. La respuesta a marcar únicamente con fibras y no con vasos es un ejemplo de la sensibilidad del cambium vascular y el tipo de sistema radicular de Bursera lancifolia.
La sensibilidad del cambium vascular debe analizarse en función del abasto de agua; es decir, ¿qué determina el calibre de los vasos y qué la respuesta de las fibras a marcar el anillo? Evidentemente, dicha sensibilidad está asociada al tipo de función destino de las iniciales fusiformes, lo que explica por qué el diámetro de los vasos es redundante. Por otro lado, la función destino hacia la formación de fibras con almidones reduce el impacto de más fibras destinadas a marcar el anillo de crecimiento. Destaca el cuerpo de los radios con células mayormente procumbentes, en series de hasta cinco células. Esta expresión morfológica favorece un abasto horizontal rápido para el transporte de agua y la translocación y conducción de metabolitos procedentes del floema, que explicaría por qué la gran mayoría de las fibras de la albura funciona para el almacenamiento; y los radios para la conducción transversal, que se explica por la talla de los individuos.
De acuerdo con Soltis et al. (2008), la familia Burseraceae apareció aproximadamente hace 50 millones de años; cuando prevalecía un clima tropical húmedo, con ausencia de estacionalidad, y ello explicaría un crecimiento radial continuo del xilema, que, a su vez, implicaría que la caducidad es un rasgo adquirido al experimentar atmósferas más desecantes. Cevallos (1998) indica que los anillos de crecimiento aparecieron en las Angiospermas en el Eoceno y en el período reciente del Oligoceno, el clima ya se había modificado con la presencia de ambientes cálidos.
La formación de anillos de crecimiento se ha atribuido principalmente a la madera de especies templadas o de clima frio; postura que ha favorecido la investigación hacia regiones geográficas que presentan claramente estacionalidad, por lo que las especies tropicales se han excluido de los estudios dendrocronológicos (Whitmore,1990). En el presente trabajo se demuestra que Bursera lancifolia es sensible a cambios en la precipitación formando anillos de crecimiento marcados por fibras.
Para investigaciones dendrocronológicas de mayor confiabilidad, se requiere incrementar el tamaño de muestra, ya que los núcleos de crecimiento analizados fue reducido (n<10 incrementos), para especies tropicales se ha sugerido que el tamaño de muestra debe ser superior (n>50) e incluso se recomienda el uso de rodajas de secciones trasversales para dar seguimiento al crecimiento en toda la circunferencia (Stahle et al., 1999).
La generación de cronologías de B. lancifolia más robustas, en términos de tamaño de muestra, puede contribuir a determinar una mejor respuesta climática, para generar datos de sitios donde existe carencia de información climática, o bien es incipiente. La corroboración de especies tropicales con anillos de crecimiento anual es también de importancia para fines de conservación, al definir las tasas de crecimiento y las condiciones climáticas que influyen en su desarrollo (Cerano et al., 2009; Cerano et al., 2011; Roig et al., 2005; Roig et al., 2012; Stahle et al., 1999; Villanueva et al., 2000; Villanueva, 2010).
Conclusión
A partir de las características anatómicas del xilema secundario de Blancifolia Bursera se corroboró la formación de anillos de crecimiento anual.
La cronología generada determinó la respuesta a variables climáticas, por medio de un análisis de función de respuesta a la precipitación acumulada estacional enero-mayo del periodo 1976 a 2001. Se sugiere considerarlos como proxy para extender los registros de la variabilidad climática regional.
El estudio plantea la consideración de especies tropicales con potencial dendrocronológico, cuyas aplicaciones pueden ser en diversos campos científicos y de la conservación de ecosistemas.
Agradecimientos
Agradecemos la hospitalidad y el apoyo de Don Guillebaldo Rondán (que en paz descanse) y a su familia, así como a los biólogos Fortunato Pérez y Luis Yépez por el apoyo durante el trabajo de recolecta y muestreo en el ejido Valle de Vázquez Morelos, México. A la M. C. Rosa Linda Cervantes Martínez y la Ing. Vicenta Constante García por su apoyo en el laboratorio.
Referencias
Alvim, P. 1964. Tree growth periodicity in tropical climates. In: Zimmermann, M. H. The formation of wood in forest trees. Acad. Press. New York, NY, USA. pp. 479-496. [ Links ]
Baas, P. and R. Vetter. 1989. Growth rings in tropical trees. IAWA Bulletin 10:95-174. [ Links ]
Barajas M., J., G. Ángeles A. y P. Solís S. 1997. Anatomía de maderas de México: especies de una selva baja alta perennifolia. UNAM. México, D.F., México. Ediciones Especiales del Instituto de Biología Núm. 16. 126 p. [ Links ]
Boninsegna, J. A., R. Villalba, L. Amarilla and J. Ocampo.1989. Studies on tree rings, growth rates and age-size relationships of tropical tree species in Misiones, Argentina. IAWA Bulletin 10:161-169. [ Links ]
Boninsegna, J. A. and R. Villalba. 1996. Dendroclimatology in the southern hemisphere: review and prospect. In: Dean, J. S., D. M. Meko and T. W. Swetnam. Tree rings. Environment and Humanity. Radiocarbon Tucson. Tucson, AZ, USA. pp. 127-141. [ Links ]
Bormann, F. H. and G. Berlyn.1981. Age and growth rate of tropical trees: New directions for research. School of Forestry & Environmental Studies. Yale University Press. New Haven, CT, USA. 137 p. [ Links ]
Brienen, R. y P. Zuidema. 2003. Anillos de crecimiento de árboles maderables en Bolivia: su potencial para el manejo de bosques y una guía metodológica. PROMAB/ IGEMA. Riberalta, Bolivia. Informe Técnico Núm. 7. 33 p. [ Links ]
Brienen, R. and P. Zuidema. 2005. Relating tree growth to rainfall in Bolivian rainforests: a test for six species using tree ring analysis. Oecologia 146:1-12. [ Links ]
Brienen, R. and P. Zuidema. 2006. Life time growth patterns and ages of Bolivian rain forest trees obtained by tree ring analysis. Journal of Ecology 94:481-493. [ Links ]
Carlquist, S. and D. Hoekman. 1985. Ecological wood anatomy of the woody Southern Flora. IAWA Bulletin 6:319-347. [ Links ]
Cardoza, G. 2010. Reconstrucción de precipitación anual por métodos dendrocronológicos para Santa María de las Cuevas, Tlaxcala, México. Tesis de Maestro en Ciencias. Facultad de Biología. Universidad Juárez del Estado de Durango. Durango, Dgo., México. 109 p. [ Links ]
Castillo, M., M. E. Sanabria e Y. Perozo. 2005. Características anatómicas del tallo de bálsamo de incienso (Bursera tomentosa Jacq. Tr. & Plauch.). Bioagro 17:17-23. [ Links ]
Cerano P., J., J. Villanueva D., Z. P. Fule, J. G. Arreola A., I. Sánchez C. y R. Valdez-Cepeda. 2009. Reconstrucción de 350 años de precipitación para el suroeste de Chihuahua, México. Madera y Bosques 15(2):27-44. [ Links ]
Cerano P., J., J. Villanueva D., R. Valdez C., J. G. Arreola A. y V. Constante G. 2011. El niño oscilación del sur y sus efectos en la precipitación en la parte alta de la cuenca del rio Nazas. Revista de Chapingo Serie de Ciencias Forestales y del Ambiente 17:207-215. [ Links ]
Cevallos, F. y S. Sergio R. 1998. Las plantas con flores. Ciencias 52: 46-57. [ Links ]
Cook, E. R. and R. H. Holmes. 1984. Program ARSTAN and user's manual. Laboratory of Tree-Ring Research. University of Arizona. Tucson, AZ, USA. 15 p. [ Links ]
Coster, C. H. 1927. Zur Anatomie und Physiologie der Zuwachszonen und Jahresringbildung in der Tropen. Annales du Jardin Botanique de Buitenzorg 37:49-161. [ Links ]
Coster, C. H. 1928. ZurAnatomie und Physiologie der Zuwachszonen und Jahresringbildung in den Tropen. Annales du Jardin Botanique de Buitenzorg 38:1-114. [ Links ]
Chowdhury, K. A. 1964. Growth-rings in tropical trees and taxonomy. Journal of the Indian Botanical Society. 43:334-343. [ Links ]
De la Paz P., C. y G. Corral. 1980. Estudio anatómico de la madera de once especies de Angiospermas. Instituto Nacional de Investigaciones Forestales. Boletín Técnico Núm. 64. México D. F., México. 79 p. [ Links ]
Détienne, P. et A. Mariaux.1977. Nature et périodicite des cernes dans le boisrouges de Méliacées africaines. Bois et Forést des Tropiques 175:53-61. [ Links ]
Détienne, P. 1989. Appearance and periodicity of growth rings in some tropical woods. IAWA Journal 10(2):123-132. [ Links ]
Devall, S., R. Parresol B. and S. J. Wright D. 1995. Dendroecological analysis of Cordia alliodora, Pseudobombax septenatum and Annona spraguei in Central Panama. IAWA Journal 16(4):411-424. [ Links ]
Enquist, B. J. and J. Leffler A. 2001. Long-Term tree ring chronologies from sympatric tropical dry-forest trees: individualistic responses to climatic variation. Journal of Tropical Ecology 17:41-60. [ Links ]
Holmes, R. L. 1983. Computer-assisted quality control in tree-ring dating and measurement. Tree-Ring Bulletin 43:69-78. [ Links ]
International Association of Wood Anatomists (IAWA). 1989. IAWA list of microscopic features for hardwood identification. IAWA Bulletin 10(3):219-332. [ Links ]
Jacoby, G. C. 1989. Overview of tree-ring analysis in tropical regions. IAWA Journal 10(2):99-108. [ Links ]
Grissino-Mayer, H. D. 2001. Assesing crossdating accuracy: a manual and tutorial for the computer program COFECHA. Tree-Ring Research 57: 67-83. [ Links ]
Koriba, K. 1958. On the periodicity of tree growth in the tropics, with reference to the mode of branching, the leaf fall and the formation of the resting bud. Gardens Straits Settlement Bulletin Singapore 17:11-81. [ Links ]
Lisi, C. S., M. F. Tomazello, P. C. Botosso, F. A. Roig, R. B. Maria, L. F. Fedele and R. A. Voigt. 2008. Tree-ring formation, radial increment periodicity, and phenology of tree species from a seasonal semi-deciduous forest in southeast Brazil. IAWA Journal 29(2):189-207. [ Links ]
Mauseth, J. D. 1988. Plant Anatomy. Addison Wesley/Benjamin Cummings. San Francisco, CA, USA. pp. 297. [ Links ]
Mattos, P. P. 1999. Identificacao de aneis anuais de crescimiento e estimative de idade e incrementannual em diametro de species nativas do pantanal da Necholandia, MS. Tesis de Doctorado. Setor de Ciencias Agrarias, Universida de Federal do Paranà, UFPR. Curitiba, Brasil. 116 p. [ Links ]
Quintanar I., A., G. Ángeles y J. A. Zavala H. 2009. Anatomía, índices físicos e hidráulicos de la madera de Gliricidiasepium (Jacq.) Steud. Maderas y Bosques 15 (2):71-91. [ Links ]
Raven, H. P., F. Evert, F. Ray and S. Eichhorn E. 2005. Biology of plants. W. H. Freeman and Company Publishers. New York, NY, USA. 370 p. [ Links ]
Roig, F. A., O. Jiménez J., J. Villanueva D., B. Luckman, H. Tiessen, A. Medina and E. J. Noellemeyer. 2005. Anatomy of growth rings at the Yucatán Peninsula. Dendrochronology 22:187-193. [ Links ]
Roig, F. A., J. Villanueva D., J. Jiménez O., W. J. Hayden, J. Barajas M. and B. H. Luckman. 2012. Anatomía de maderas en comunidades rurales de Yucatán. INIFAP CENID RASPA. Gómez Palacio, Dgo., México. Libro Técnico Núm. 4. 242 p. [ Links ]
Rosero, J. 2009. Dendrocronología de àrvores de mogno Swietenia macrophylla King., Meliaceae, ocorrentes na floresta tropical Amazonica do Departamento de Madre de Dios. Perú, Piracicaba. 129 p. [ Links ]
Rzedowski, J. 1988. Vegetación de México. Editorial Limusa, México D.F., México. 189 p. [ Links ]
Rzedowski, J. y F. Guevara F. 1992. Burseraceae. Flora del Bajío y de regiones adyacentes. Fasciculo 3. 46 p. [ Links ]
Rzedowski, J., L. Medina R. y G. Calderón. 2004. Las especies de Bursera (Burseraceae) en la cuenca superior del Rio Papaloapan (México). Acta Botánica Mexicana 66: 23-151. [ Links ]
Soltis, D. E., D. Bell, K. Sangtae and P. S. Soltis. 2008. Origin and early Evolution of Angiosperms. Annals of the New York Academy of Sciences 1133:3-25. [ Links ]
Schweinggruber, F. H. 1996. Tree ring and environment. Paul Haupt. Federal Institute for Forest. Snow and Landscape. Berne, Switzerland. 609 p. [ Links ]
Stokes, M. A. and T. L. Smiley. 1968. An Introduction to Tree Ring Dating. University of Chicago Press. Chicago, IL, USA. 33 p. [ Links ]
Stahle, D. W., M. K. Cleaveland, M. D. Therrell, D. A. Gay, R. D. D'Arrigo, P. J. Krusic, E. R. Cook, R. J. Allan, J. E. Cole, R. B. Dunbar, M. D. Moore, M. A. Stokes, B. T. Burns, J. Villanueva-Diaz and L. G. Thompson. 1998. Experimental Dendroclimatic Reconstruction of the Southern Oscillation. Bulletin of the American Meteorological Society 79 (10):2137-2152. [ Links ]
Stahle, D. W., P. T. Mushove, M. K. Cleveland, F. Roig and G. A. Haynes. 1999. Management implications of annual growth rings in Pterocarpus angolensis from Zimbabwe. Forest Ecology and Management 124(2-3):217-229. [ Links ]
Tomazello, M. F. and N. Cardoso. 1999. Seasonal variations of the vascular cambium of Teak (Tectona grandis L.) in Brazil. In: Wimmer, R. and R. E. Vetter. Tree Ring Analysis. CAB International. Oxon, England. pp. 147-154. [ Links ]
Tomazello, M. F., P. Botosso y C. Lisi. 2001. Anàlise e aplicacao dos aneis de crescimento das àrvores como indicadores ambientais: dendrocronologiae dendroclimatologìa. pp. 117-143. In: Maia, N. B., H. L. Martos y W. Barrella. Indicadores ambientais: conceitos e aplicacoes. EDUC, COMPED, INE. Sao Paulo, Brasil. pp. 32. [ Links ]
Valdez-Hernández, M., J. L. Andrade, P. C. Jackson and M. Rebolledo V. 2010. Phenology of five tree species of a tropical dry forest in Yucatan, México: effects of environmental and physiological factors. Plant Soil 329:155-171. [ Links ]
Vetter, R. E. and P. C. Botosso. 1989. Remarks on age and growth rate determination of Amazonian trees. IAWA Bulletin 10:33-145. [ Links ]
Villalba, R. 2000. Dendroclimatology: a southern hemisphere perspective. In: Smolka P. and W. Volkheimer. Paleo and Neoclimates of the southern hemisphere:the state of the Arts. Springer. Berlin, Germany. pp. 28-57. [ Links ]
Villanueva D., J., D. W. Stahle, M. K. Cleaveland y M. D. Therrell. 2000. Estado actual de la dendrocronología en México. Ciencia Forestal en México 25 (88):3- 34. [ Links ]
Villanueva D., J., J. Cerano, P., D. W. Stahle, V. Consante G., S. Vázquez, J. Estrada A. y J. Benavides S. 2010. Árboles longevos de México. Ciencia Forestal en México 1 (2):3-7. [ Links ]
Whitmore, T. C. 1990. An Introduction to Tropical Rain Forests. Nordic Journal of Botany 11(5):548. [ Links ]
Worbes, M. 1989. Growth rings, increment and age of trees in inundation forests, savannas and a mountain forest in the Neotropics. In: Baas, P. P. and R. E. Vetter Growth Rings in Tropical Woods. IAWA Bulletin 10:109-122. [ Links ]
Worbes, M. 1995. How to measure growth dynamics in tropical trees-A review. IAWA Journal 16(4):337-351. [ Links ]
Zumaeta, L. E. C. 2008. Dendrocronología en árboles de Tornillo, Cedrelinga cateniformis Ducke (Fabaceae). Tesis de Maestría Centro de Investigaciones Jenaro Herrera en el noreste de la Amazonia, Región Loreto-Perú. Universidad Nacional Agraria La Molina. Lima, Perú. 129 p. [ Links ]



