










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de ciencias forestales
versión impresa ISSN 2007-1132
Rev. mex. de cienc. forestales vol.5 no.25 México sep./oct. 2014
Artículos
Rompimiento de la dormancia en semillas y propagación in vitro de Cordia elaeagnoides A. DC.
Breaking of dormancy in seeds and in vitro propagation of Cordia elaeagnoides A. DC.
Fernando Santacruz Ruvalcaba1, José Juvencio Castañeda Nava1, Ana María Gaspar Peralta1, Nancy Núñez Sandoval1 y Antonio Mora Santacruz1
1 Centro Universitario de Ciencias Biológicas y Agropecuarias, Universidad de Guadalajara. Correo-e: srf22191@cucba.udg.mx
Fecha de recepción: 4 de abril de 2014;
Fecha de aceptación: 23 de junio de 2014.
Resumen
Cordia elaeagnoides es una especie maderable de selvas bajas caducifolias, que por la dificultad de sus semillas para germinar, la especie tiene limitaciones para reproducirse. En este contexto, se desarrolló un método para romper la dormancia y propagarla asexualmente por medio de proliferación de yemas axilares in vitro. Se evaluó la presencia de embriones sanos en 1 100 semillas de 11 lotes (100 semillas por lote); se constató que 39.45 % del total corresponden a esta condición. Se probaron 260 ppm de ácido giberélico con inmersión durante 24, 48 y 72 h; la primera alcanzó 96 % de germinación. Para establecer la propagación in vitro, se hizo rescate de embriones en el medio de cultivo Murashige y Skoog (MS) adicionado con 2 mgL-1 de benciladenina; se logró 82 % de germinación. Las plántulas obtenidas se utilizaron para la proliferación de yemas axilares, sobre las cuales se evaluaron los medios MS y McCown's, y los reguladores de crecimiento 2-isopentiladenina, cinetina y benciladenina en dosis de 1, 2 y 3 mgL-1; se obtuvo un promedio de 3.03 brotes por explante tanto en cinetina como benciladenina. Para la rizogénesis in vitro se utilizó ácido 3-indolbutírico a dosis de 2.5 y 5 mgL-1, más adenina en oscuridad o fotoperiodo; los datos revelaron que cualquier dosis del ácido 3-indolbutírico induce la rizogénesis. Se concluye que para la germinación de semillas de Cordia elaeagnoides es conveniente aplicar ácido giberélico para eliminar la dormancia y para la propagación asexual in vitro, cinetina y benciladenina en las proporciones indicadas.
Palabras clave: Ácido giberélico, Cordia elaeagnoides A. DC., germinación, proliferación de yemas axilares, rescate de embriones, rizogénesis.
Abstract
Cordia elaeagnoides A. DC. is a timber species of tropical deciduous forests, which from the difficulty of their seeds to germinate, the species has limitations to reproduce. In this context, a method was developed to break dormancy and asexually propagate it through axillary bud proliferation in vitro. The presence of healthy embryos in 1 100 lots of 11 seeds (100 seeds per lot) were evaluated; it was found that 39.45 % of them correspond to this condition. 260 ppm gibberellic acid were tested with 24, 48 and 72 immersion hours; the first one reached 96 % of germination. To establish in vitro propagation, embryo rescue was made in the Murashige and Skoog (MS) culture medium supplemented with 2 mg L-1 benzyladenine; 82 % germination was achieved. Seedlings were used for axillary bud proliferation, on which the MS and McCown's media growth regulators 2-isopentiladenina, kinetin and benzyladenine at doses of 1, 2 and 3 mgL-1 were evaluated; an average of 3.03 shoots per explant was obtained in both kinetin and benzyladenine. For in vitro rooting 3-indole butyric acid at doses of 2.5 and 5 mg L-1 were used, plus adenine in darkness or photoperiod; data revealed that any 3-indole butyric acid dose induces rooting. It is concluded that for the germination of Cordia elaeagnoides seeds gibberellic acid should be applied to remove dormancy and for its asexual propagation in vitro, kinetin and benzyladenine in the proportions previously indicated must be used.
Key words: Gibberellic acid, Cordia elaeagnoides A. DC., germination, axillary bud proliferation, embryo rescue, rhizogenesis.
Introducción
Las selvas bajas caducifolias reúnen una riqueza de especies considerable; habitan en ellas, por ejemplo, Cordia alliodora (Ruiz et Pav.) Oken, Cordia elaeagnoides A. DC., Tabebuia rosea (Bertol.) DC. y Tabebuia palmeri Rose. C. elaeagnoides es integrante de la familia Boraginaceae, misma que se caracteriza, entre otros rasgos, por presentar incompatibilidad gamética (Gibbs, 1986; Brys et al., 2008; Loha et al., 2009; Mc Mullen, 2012). A esta especie se le denomina de distintas maneras en función de las regiones en las que crece: "cueramo" en Michoacán, "bojote" en Guerrero, "barcino" en Jalisco y "anacahuite" en Oaxaca. Su distribución en México abarca desde Jalisco hasta Chiapas y tiene gran importancia en su ecosistema pues se le considera gregaria del dosel (Van Groenendael et al., 1996); por otro lado, es muy utilizada en la industria de la madera, ya que con ella se elabora una gran diversidad de productos tales como llaveros, joyas, máscaras, cajas, muebles, instrumentos musicales, juegos de salón y artículos de cocina (Pérez, 1993). C. elaeagnoides tiene problemas relacionados con el aborto de embriones, lo cual disminuye la germinación en condiciones naturales, a lo cual habría de agregarse que sus semillas también pueden ser atacadas por roedores generalistas (Van Groenendael et al., 1996).
Las semillas constituyen la unidad móvil y de reproducción sexual de las plantas, y cumplen con la función de multiplicar y perpetuar la especie (Doria, 2010) y permiten que esta llegue a nuevos sitios y microambientes (Taiz y Zeiger, 1998).
Con el fin de que un cultivo sea exitoso es imprescindible considerar tanto la calidad de la semilla (Baskin y Baskin, 2014), como algunos factores ambientales (disponibilidad de agua, temperatura, luz, oxígeno y dióxido de carbono), ya que influyen en el porcentaje y la velocidad de germinación, muchos de los cuales son más o menos específicos para distintos taxa (Rodríguez, 2008).
La viabilidad de la semilla está determinada por características genéticas de la planta progenitora, las condiciones climáticas en distintas etapas (durante la floración, el desarrollo y la maduración del fruto), el grado de madurez de la semilla en la cosecha y el manejo en su colecta (Gallo et al., 2012; Baskin y Baskin, 2014).
Se ha observado que especies de plantas originarias de ambientes en donde se presentan con frecuencia heladas y/o sequías son más propensas a cierto tipo de latencia en comparación con especies de ambientes benignos (Jurado y Flores, 2005).
Otro factor que influye considerablemente en la conservación de la viabilidad de la semilla, es la calidad y cantidad de sustancias químicas contenidas en el embrión (Doria, 2010)
El ácido giberélico (AG3) activa el crecimiento vegetativo del embrión durante la germinación y promueve la producción o secreción de enzimas hidrolíticas principalmente α-amilasa, enzima involucrada en la solubilización de las reservas del endospermo (Salinas et al., 2002). El almidón y las proteínas son desdobladas por varias enzimas; los azúcares, aminoácidos y otros productos son transportados para el crecimiento del embrión (Taíz y Zeiger, 1998).
El cultivo in vitro permite propagar vegetales de manera asexual, y por lo tanto, multiplicar masivamente a las especies. Los reguladores de crecimiento más utilizados en la formación de brotes son benciladenina, cinetina y bencilaminopurina (del grupo de las citocininas) y la inducción puede provenir de callo, nudos o meristemos apicales (Phulwaria y Shekhawat, 2013). Se han documentado casos de rizogénesis dentro de la familia Boraginaceae al aplicar ácido indol-3-acético, ácido indol-3-butírico y ácido naftalenacético, reguladores que varían en cuanto a la eficiencia en función del taxón (Edson et al., 1996; Martin, 2003; Schuler et al., 2005; Phulwaria y Shekhawat, 2013).
Debido a que Cordia elaeagnoides en la región Costa del estado de Jalisco produce muchas semillas con aborto de embriones y presenta dificultad en la germinación de semillas, se planteó como objetivo del presente trabajo, desarrollar una técnica para romper la dormancia y propagar asexualmente la especie por medio de proliferación de yemas axilares in vitro.
Materiales y Métodos
Recolección de semillas
Las semillas de C. elaeagnoides se colectaron en Tomatlán, Jalisco, México (19°56'3'' N y 105°14'8''O), a 50 msnm. Se eligieron veinte ejemplares sanos, de buena altura, fuste limpio y amplia masa foliar, con flores que mantuvieran color café (indicio que los frutos están en el mejor momento); se almacenaron en costales y cada muestra se numeró en función del árbol del que fue tomada (figuras 1a, c). Se trasladó la semilla al Laboratorio de Cultivo de Tejidos Vegetales del Departamento de Producción Agrícola del Centro Universitario de Ciencias Biológicas y Agropecuarias (CUCBA) para su limpieza, pruebas de germinación y cultivo in vitro.

Identificación de embriones sanos
Para seleccionar semillas con embriones sanos se compararon dos métodos: 1) la prueba de corte en 11 lotes con un total 100 semillas evaluadas por lote. Se partió la testa de la semilla con una tijera; se observó el color y la textura del endospermo y se verificó que el embrión estuviera bien desarrollado. Se contabilizó el número de semillas con embriones sanos por lote. 2) la técnica de columna densimétrica, para determinar por medio del peso, si las semillas estaban en buenas condiciones o eran semillas inviables para aplicarla se utilizó una máquina que consta de un cilindro de metacrilato transparente con un separador neumático de semillas, que por medio de inyección de aire, separa las semillas de menor peso (inviables) hacia un recipiente al que suben; y las que tienen un mayor peso quedan en la parte inferior de este cilindro. Se contabilizó el porcentaje de semillas sanas y las inviables, para establecer un comparativo entre estos dos métodos.
Tratamientos de semillas con ácido giberélico
Se preparó una dilución de ácido giberélico (AG3) a 260 ppm en la que se colocaron 30 semillas de cada uno de 20 lotes, con tiempos de inmersión de 24, 48 y 72 h; se contabilizó el número de semillas germinadas de acuerdo al tiempo de inmersión en dichos tratamientos, comparando con un testigo sin tratamiento. Las semillas fueron sembradas en charolas germinadoras, como sustrato se utilizó una mezcla de musgo canadiense y vermicomposta en una proporción de 40:60 respectivamente. Todas las charolas fueron trasladadas a un invernadero a una temperatura media de 30 °C, en donde se regaron diariamente. Las evaluaciones de la germinación se realizaron a los 7, 14, 21 y 28 días.
Proliferación de yemas axilares in vitro
Para el establecimiento in vitro de Cordia eleagnoides fue necesario el rescate de embriones cigóticos. Las semillas se desinfectaron en una solución de hipoclorito de sodio al 3 % por un periodo de 10 min. Se enjuagaron con agua estéril y se dejaron en remojo por 24 h. Para la extracción del embrión se utilizó un microscopio estereoscópico Carl Zeiss Stemi DR 1040; se eliminó la testa y los embriones se colocaron en el medio de cultivo MS (Murashige y Skoog, 1962) adicionado con 2 mgL-1 de benciladenina (BA), 8 gL-1 de agar.
Cuando se obtuvieron los brotes, se desarrolló un experimento con un arreglo trifactorial (3x3x2); las variables fueron los reguladores de crecimiento 2-isopentiladenina (2ip), benciladenina (BA) y cinetina (KIN), cada uno con tres concentraciones (1, 2 y 3 mgL-1) y en dos diferentes medios basales, MS y McCown (Lloyd y McCown, 1980).
La elaboración de los medios de cultivo se realizó de la siguiente manera: los medios basales de los medios referidos fueron suplementados con vitaminas L2 (Phillips y Collins, 1979), 3 % de sacarosa y, para solidificar el medio, se aplicaron 8 gL-1 de Phytagel® (Sigma Chemical Co. Cat. A-1296). Se emplearon frascos de vidrio de alimento infantil de 100 mL de capacidad como contenedores, a los cuales se les adicionaron 25 mL del medio de cultivo respectivo. Se hicieron cinco repeticiones por tratamiento (un frasco con un brote se consideró como una repetición). La evaluación del número de brotes generados se llevó acabo a los 45 días de iniciado el experimento. Ambos sustratos se ajustaron a un pH 5.8, previo a la esterilización en autoclave (121 °C a 1.2 kg cm-2 de presión durante 15 min). La incubación del material vegetal en los experimentos fue a 27 ± 2 °C con un fotoperiodo de 16 h de luz fluorescente a 1 500 lux (25 mmol s-1 m-2).
Inducción de rizogénesis y elongación de tallos
Para la inducción de raíces se estableció un ensayo con un diseño en arreglo trifactorial 3x2x2, en el cual se utilizó el ácido indol3butírico en tres diferentes concentraciones (0, 2.5 y 5 mgL-1), la presencia o ausencia de adenina (40 mgL-1), fotoperiodo (16 h luz/8 h oscuridad) y oscuridad durante las 24 h. Los tratamientos que requerían de oscuridad se cubrieron completamente con papel aluminio. El medio utilizado fue McCown y como gelificante 3 gL-1 de Phytagel®. La incubación del material vegetal en los experimentos se realizó a una temperatura de 27 ± 2 °C. Cada tratamiento contó con cinco repeticiones y fueron evaluados a los 45 días.
Análisis estadístico
Para el análisis estadístico de la propagación asexual mediante el cultivo in vitro de Cordia elaeagnoides se utilizó el software Statgraphics Plus 4.0®. Se realizaron los análisis de varianza para cada uno de los experimentos mediante los cuales se evaluaron las variables de respuesta correspondientes, además de realizar la comparación múltiple de medias mediante la prueba de diferencias mínimas significativas (LSD).
Resultados y Discusión
Identificación de embriones sanos
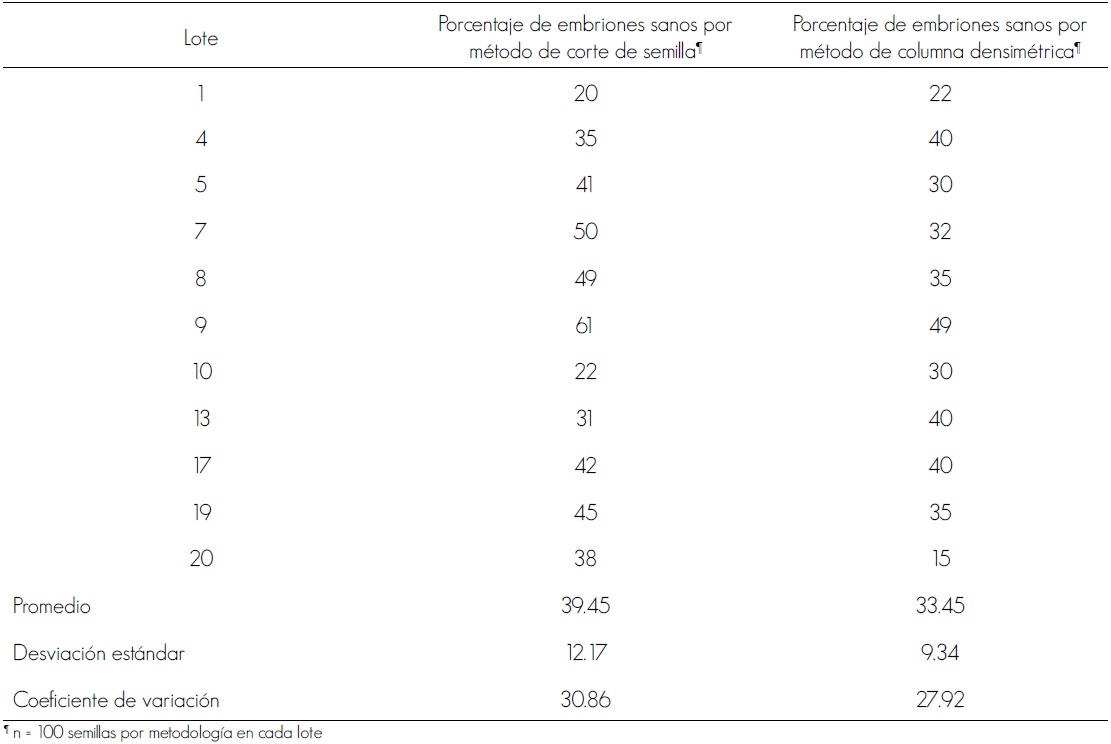
El porcentaje de embriones sanos fue bajo; en la prueba que se realizó por corte en la semilla, fueron 39.45 % en promedio y 60.55 % estaban dañados (Figura 1b); con la separación por columna densimétrica se obtuvieron 33.45 % sanos y 66.55 % dañados; no hubo grandes diferencias entre las dos técnicas, sin embargo por el método manual se contabilizó un mayor número de embriones sanos (Cuadro 1), lo que puede atribuirse a un error de observación, ya que es difícil determinar si están enteros y completamente desarrollados. La técnica de la columna densimétrica se perfila como una prueba rápida y confiable. Los conteos son importantes, ya que permiten observar tendencias en la sanidad de semillas.
Cuadro 1. Comparación de dos métodos para determinar el porcentaje de semillas de Cordia elaeagnoides A. DC. con embriones sanos en 11 lotes.
Determinación de tratamientos de semillas con AG3 para la germinación
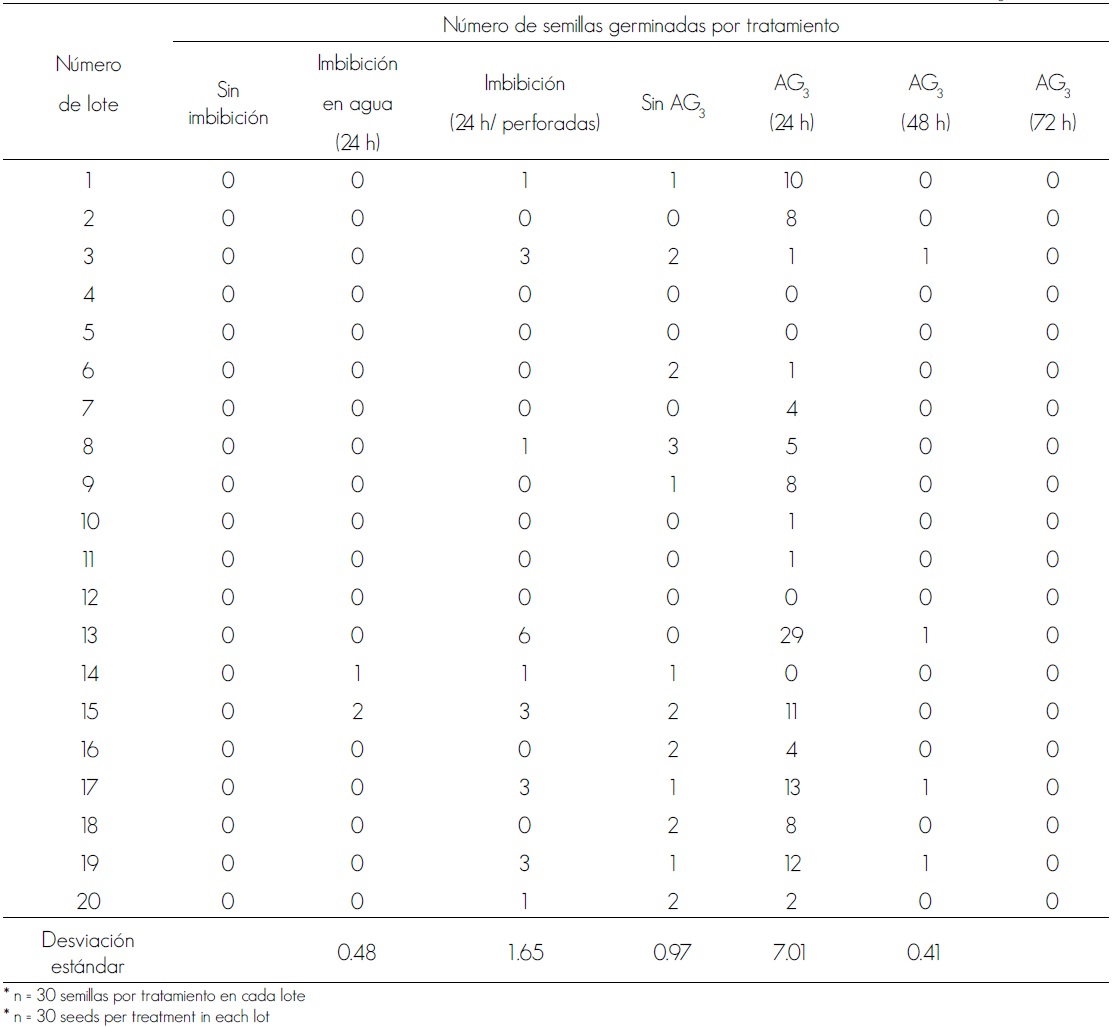
En el tratamiento que consideró 3 tiempos de imbibición y una dosis de 260 ppm de AG3 hubo respuestas de germinación variables; las semillas de los lotes 13, 17 y 19 tuvieron los mejores resultados (Cuadro 2); la germinación C. elaeagnoides fue beneficiada por la ruptura de la dormancia, como efecto del AG3; al comparar los tres tiempos, el óptimo fue el correspondiente al lote 13 (24 h de imbibición en 260 ppm de AG3), ya que de él germinó 96.6 % de las semillas. La activación del embrión generalmente se debe a la estimulación enzimática mediada por el AG3; Salinas et al. (2002) indican que este ácido promueve la síntesis de algunas enzimas, entre las cuales destacan la α-amilasa y la dextrina límite, la α y β glucosidasas, las endo-β gluconasas y las endoxilasas, lo que permite la movilización de hidratos de carbono para el crecimiento y desarrollo del embrión.
Durante las pruebas de germinación de las semillas de Cordia elaeagnoides se observó en la mayor parte de los lotes estudiados que no tuvieron AG3 no se estimuló dicha respuesta. Los lotes de semillas 17, 19 y 13 (Cuadro 2) con tratamiento de imbibición de 24 h y 260 ppm de AG3 fueron los que alcanzaron más altos porcentajes de germinación y de ellos sobresalió el 13. Se reconocen limitaciones en la obtención de semillas completas y la germinación de ellas en especies de la familia Boraginaceae, lo que parece responder a diversos desórdenes morfológicos o fisiológicos, problema que está relacionado con la distilia, que es una reacción de incompatibilidad que según Brys et al. (2008) y Mc Mullen (2012) afecta a muchas especies del género Cordia.
Cuadro 2. Rompimiento de la dormancia en semillas de Cordia elaeagnoides A. DC. mediante tratamientos de inmersión en ácido giberélico (AG3).
Proliferación de yemas axilares en brotes de Cordia elaeagnoides
La propagación in vitro de la especie se inició con el establecimiento a partir del rescate de embriones cigóticos de la semilla; los resultados obtenidos con esta técnica fueron satisfactorios, pues del total de los embriones rescatados se obtuvo 82 % de germinación. A los 2 días posteriores al establecimiento, se inició el proceso de germinación con el despliegue de los cotiledones (Figura 1d). Después de siete días de iniciado el cultivo, el desarrollo de la planta era óptimo, incluso presentaron indicios radiculares. A los 30 días los problemas oxidativos comenzaron a presentarse en la mayoría de las plantas desarrolladas. La técnica de rescate de embriones ha sido utilizada anteriormente exitosamente en Persea americana Mill, en un programa para la multiplicación vegetativa con el fin de llevar a cabo mejoramiento genético. Los embriones maduros e inmaduros fueron extraídos y cultivados en medio MS a la mitad de su concentración y se les adicionó 0.5 mgL-1 de 6-bencil amino purina y la misma cantidad de ácido giberélico (Rodríguez et al., 1999). Pinus maximartinezii Rzedowski también ha sido propagada de esta forma, con el propósito de conservar y expandir a esta especie en peligro de extinción (Ojeda et al., 2006).
Una vez que se corroboró la viabilidad de las semillas y la sanidad de los embriones, se eliminaron los compuestos fenólicos (su abundante producción se observó en la germinación en papel absorbente), lo que se logra mecánicamente, al raspar la testa; este procedimiento ayuda a evitar la oxidación del medio de cultivo y a eliminar la dureza de la cubierta seminal. Se pudo advertir que, a pesar de que la testa se raspara casi en su totalidad, no hubo respuesta germinativa; esto puede indicar que la latencia física y la presencia de compuestos fenólicos no son el único obstáculo que enfrentan las semillas de C. elaeagnoides. Habría de suponerse que existen otros inhibidores de la germinación, como pudiera ser el ácido abscísico (ABA), que protege a la planta en escenarios de estrés al inhibir la síntesis de proteínas y de ácidos nucleicos e inducir la abscisión y la senescencia; además afecta las membranas y mantiene el nivel de otras hormonas y controla el almacenamiento de proteínas de reserva (Zhang et al., 2012); se recomienda realizar un análisis de concentración de ABA en las semillas de C. elaeagnoides para determinar su participación en este proceso.
Después de 45 días no hubo significancia en la producción de brotes por el tipo de medio (p= 0.1714), ni por la cantidad de regulador (p= 0.0714); en el caso del regulador utilizado hubo alta significancia (p=0.0004), al realizar una comparación múltiple de medias con ayuda de la prueba de Diferencias Mínimas Significativas (LSD). La mayor estimulación de brotes pasado este lapso la ejercieron los tratamientos en presencia del regulador de crecimiento KIN y BA, con una eficiencia superior de hasta 1,5 veces más que 2ip (Cuadro 3) (Figura 1f). Con respecto a la concentración, aunque en la prueba de análisis de varianza no se muestra una diferencia, se confirma cierta tendencia positiva al generar un número de brotes más alto y una menor oxidación de los tejidos cuando son utilizadas concentraciones más bajas (1 mgL-1). Esto ha sido mencionado por Balla et al. (2002) quienes explican que la adición de citocininas tiene un efecto represor de la dominancia apical, lo que provoca la proliferación de brotes por medio de la estimulación de las yemas axilares. Los tejidos oxidados fueron muy notorios conforme el trascurso del tiempo, en particular aquellas estructuras con el regulador 2ip y en la mayor concentración 3 mgL-1, además de generar callo pubescente en la parte basal (Figura 1g).
Cuadro 3. Comparación múltiple de medias de Diferencias Mínimas Significativas para el número de brotes de Cordia elaeagnoides A. DC. a los 45 días.

Fue estrictamente necesario adicionar reguladores de crecimiento para inducir los brotes en C. elaeagnoides, lo que difiere con lo consignado por Tacoronte et al. (2004), quienes propagaron otra especie maderable tropical, Swietenia macrophylla King, por medio de yemas axilares y con la mejor respuesta en ausencia de reguladores de crecimiento. En Arnebia hispidissima (Lehm.) A. DC., especie que pertenece también a la familia Boraginaceae, el uso de los reguladores KIN y BAP es favorable para la producción de brotes (Phulwaria y Shekhawat, 2013).
Aunque estadísticamente no se confirmó significancia con respecto a los medios basales, los cultivos en el medio MS comenzaron a generar un mayor número de brotes en un principio, pero en los análisis posteriores, estos brotes se redujeron por el aumento en la oxidación, y mostraron un ligero amarillamiento. En el medio McCown, por el contrario, los cultivos manifestaron un número inferior de brotes, pero la oxidación fue menor y presentan una coloración muy favorable. Cualitativamente se mostró una estrecha relación entre el tipo de medio de cultivo utilizado y el regulador adicionado, de lo que resultó ser más eficiente la utilización de BA para el medio MS y KIN para el medio McCown.
La oxidación puede deberse principalmente por la acumulación de compuestos fenólicos y principalmente al etileno. También a cierta sensibilidad a los reguladores de crecimiento, pues fue más notoria en aquellos que presentaban una mayor concentración de dichos compuestos (Basail et al., 2006).
Schuler et al. (2005) demostraron que Cordia alliodora presenta una tasa de pérdida muy alta en el cultivo de tejidos vegetativos y la catalogaron como una especie recalcitrante para el cultivo in vitro. C. elaeagnoides también podría considerarse dentro de este tipo de especies por la tasa de pérdida ocasionada por la oxidación y acumulación de etileno, lo cual fue difícil controlar y causó detrimento considerable a los brotes.
Inducción de rizogénesis y elongación de tallos
Después de transcurridos 45 días del establecimiento, dos factores tuvieron alta significancia para estimulación de raíces: la concentración del regulador (p=0.0001) y la presencia o ausencia de luz para inducción del número de las mismas (p=0.0000). La utilización de 1500 lux con un fotoperíodo de 16 h luz y 8 h oscuridad fue el mejor tratamiento para generar raíces (1.43 raíces por explante a los 45 días).
Al realizar la comparación múltiple de medias mediante la prueba LSD para la inducción de raíces, se encontró que la mayor estimulación de rizogénesis después de los 45 días (Figura 1e), la ejercían los tratamientos en presencia de ácido indol-3 butírico en concentraciones de 2.5 y 5 mgL-1 (Cuadro 4). Las auxinas están involucradas en casi todos los aspectos del crecimiento y desarrollo de la planta, desde el estado embrionario hasta el estado reproductivo. Una de sus funciones es la elongación de los tallos por medio de la estimulación de la división celular y también la formación del sistema radicular (Phulwaria y Shekhawat, 2013). Por ello su aplicación exógena resulta ser esencial en diferentes especies para inducción de rizogénesis, como sucede con C. elaeagnoides, pues está estrechamente emparentada con Cordia alliodora; Schuler et al. (2005) intentaron su enraizamiento solamente con la adición de sacarosa (15 gL-1 L-1) y carbón activado (2 gL-1), y no lo lograron. Por el contrario, Tacoronte et al. (2004) obtuvieron el enraizamiento de Swietenia macrophylla por medio de un balance entre auxinas y citocininas (8. 9 mgL-1 de ácido indol 3-butírico (AIB), 2.2 mgL-1 de ácido naftalenacético (ANA), 4.0 mgL-1 de KIN).
Cuadro 4. Comparación múltiple de medias de Diferencias Mínimas Significativas para el número de raíces de Cordia elaeagnoides A. DC. estimulados a los 45 días.

Después de transcurrido el periodo indicado se observó una tasa de pérdida de tejidos muy alta, todo debido a la excesiva producción de etileno derivada de la acción del ácido indolacético como otras auxinas sintéticas (Zhao y Hasenstein, 2009). Muchos de los efectos de las auxinas se deben a la coparticipación con el etileno al inhibir su producción; en este caso la actividad de las auxinas se afectó, por lo que se recomienda adicionar agentes antioxidantes al medio de cultivo para el enraizamiento.
La utilización de 1 500 lux fue altamente significativa (P=0.0001) para la elongación de los tallos, lo que aumentó 0.7 cm en promedio, mientras que en etiolación (sin iluminación) sólo aumentaron 0.3 cm. La ausencia de luz prepara a los explantes para el enraizamiento por medio de la estimulación de la producción de auxinas, por lo que la aplicación de técnicas como la etiolación permite iniciar el proceso de rizogénesis (Castro y Sánchez, 2010). La presencia de luz conserva un mayor número de brotes y, por el contrario, la etiolación genera una alta tasa de pérdida de los tejidos, en primera instancia debido a la baja tasa de fotosíntesis, por lo que los tejidos se comienzan a tornar amarillentos y posteriormente mueren. El segundo efecto negativo de la etiolación fue la estimulación de producción de etileno, por lo que la abscisión y muerte de los brotes era cada vez mayor conforme el transcurso del tiempo.
Se recomienda experimentar con diferentes periodos de etiolación, que favorezcan mejores resultados y observar la elongación de los tallos, ya que en el experimento aquí descrito no fue posible por la pérdida de la mayoría de los explantes.
Por lo anteriormente expuesto en el presente trabajo se considera que C. elaeagnoides amerita ser estudiada más a detalle en la biología de la reproducción para tener mayor conocimiento de sus limitaciones en la germinación y viabilidad de la semilla, ya que es un árbol de interés maderable de las selvas bajas caducifolias de la vertiente del Pacífico de México.
Conclusiones
La selección de semilla en C. elaeagnoides con embriones sanos es importante para asegurar un porcentaje alto de germinación, misma que se favorece con períodos de inmersión de la semilla en ácido giberélico (AG3) por 24 h, preferentemente. La propagación in vitro es factible en la especie, a partir del rescate de embriones cigóticos, y la proliferación de brotes a partir de yemas axilares con la adición de las citocininas, cinetina o benciladenina y la inducción de raíces con la presencia de la auxina ácido indol3butírico.
Referencias
Balla, J., J. Blažková, V. Reinnov and S. Prochhváa. 2002. Involvement of auxin and cytokinins in initiation of growth of isolated peabuds. Plant Growth Regulation 38: 149–156. [ Links ]
Basail, M., R. G. Kosky, V. Medero, E. Otero, M. Torres, M. Cabrera, J. López, M. García, A. Santos, A. Rayas, J. Ventura, M. Bauta, M. Álvarez, E. Páz, Y. Beovidez, J. Albert, A. Espinosa y J. García. 2006. Influencia de reguladores e inhibidores del crecimiento en la multiplicación de brotes axilares del cultivar híbrido ‘FHIA-21' (AAAB) en sistema de inmersión temporal. Biotecnología Vegetal 6: 23 – 28. [ Links ]
Baskin, C. C. and J. M. Baskin. 2014. Seeds: ecology, biogeography, and evolution of dormancy and germination. Academic Press. San Diego, CA, USA. 1600 p. [ Links ]
Brys, R., H. Jacquemyn, M. Hermy and T. Beeckman. 2008. Pollen deposition rates and the functioning of distyly in the perennial Pulmonaria officinalis (Boraginaceae). Plant Systematics and Evolution 273:1–12. [ Links ]
Castro, R. D. y G. A. Sánchez R. 2010. Propagación clonal in vitro de Eucalyptus pellita F. Muell a partir de árboles plus. Temas Agrarios 15: 34 - 43. [ Links ]
Doria, J. 2010. Generalidades sobre las semillas: su producción, conservación y almacenamiento. Cultivos Tropicales 31 (1): 74-85. [ Links ]
Edson, J. L., A. D. Leege-Brusven, R. L. Everetl and D. L. Wenny. 1996. Minimizing growth regulators in shoot culture of an endangered plant, Hackelia venusta (Boraginaceae). In vitro Cellular & Developmental Biology 32: 267-271. [ Links ]
Gallo, C., J. M. Enrico, R. Craviotto y M. Arango. 2012. Variabilidad de la viabilidad y vigor de lotes de semillas de soja con presencia de simientes verdes pertenecientes a cultivares de distintos grupos de maduración producidos en dos fechas de siembra. Revista de Investigaciones Agropecuarias 38: 133-140. [ Links ]
Gibbs, P. E. 1986. Do homomorphic and heteromorphic self-incompatibility systems have the same sporophytic mechanism? Plant Systematics and Evolution 154: 285-323. [ Links ]
Jurado, E. and J. Flores. 2005. Is seed dormancy under environmental control or bound to plant traits? Journal of Vegetation Science 16: 559-564. [ Links ]
Lloyd, G. and B. H. McCown. 1980. Commercially-feasible micropropagation of Mountain Laurel (Kalmia latifolia) by shoot tip culture. Procedings International Plant Propagators Society 30: 421-427. [ Links ]
Loha, A., M. Tigabu and A. Fries. 2009. Genetic variation among and within populations of Cordia africana in seed size and germination responses to constant temperatures. Euphytica 165: 189-196. [ Links ]
Martin, K. P. 2003. Rapid in vitro multiplication and ex vitro rooting of Rotula aquatica Lour., a rare rhoeophytic woody medicinal plant. Plant Cell Reports 21: 415–420. [ Links ]
Mc Mullen, C. K. 2012. Pollination of heterostylous Gálapagos native, Cordia lutea (Boraginaceae). Plant Systematics and Evolution 298: 569-579. [ Links ]
Murashige, T. and F. Skoog. 1962. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiologia Plantarum 15: 473-497. [ Links ]
Ojeda Z., M. C., H. L. Olvera, L. M. Ramos, M. J. Star, T. Cepeda, P. Alférez, L. I. Donjuan, E. O. Sáenz, R. Sáenz y E. C. Cerda. 2006. Multiplicación in vitro del Piñón Azul Pinus maximartinezii (Rzedowski). Phyton 75: 109-113. [ Links ]
Orantes G., C., M. A. Pérez F., T. M. Rioja P. y E. R. Garrido R. 2013. Viabilidad y germinación de semillas de tres especies arbóreas nativas de la selva tropical, Chiapas, México. Polibotánica 36: 117-127. [ Links ]
Pérez O., C. P. 1993. Anatomía de la madera de ocho especies con importancia en las artesanías del estado de Michoacán. Acta Botánica Mexicana 23: 103-136. [ Links ]
Phillips, G. C. and G. B. Collins. 1979. In vitro tissue culture of selected legumes and plant regeneration from callus cultures of red clover. Crop Science 19:59-64. [ Links ]
Phulwaria, M. and N. S. Shekhawat. 2013. An efficient in vitro shoot regeneration from immature in florescence and ex vitro rooting of Arnebia hispidissima (Lehm). DC. - A red dye (Alkannin) yielding plant. Physiology Molecular Biology of Plants 19(3): 435–441. [ Links ]
Rodríguez E., M. R. 2008. Influencia de la temperatura en la germinación de semillas de Caesalpinia spinosa (Molina) Kuntze «taya» de cuatro localidades del Departamento La Libertad, Perú. Arnaldoa 15: 87 – 100. [ Links ]
Rodríguez, N. N., M. Capote y V. Zamora. 1999. Cultivo in vitro del aguacatero (Persea americana Mill.). Revista Chapingo Serie Horticultura 5: 231-237. [ Links ]
Salinas, A. R., A. M. Yoldjian, M. L. Dietrich, R. M. Craviotto y V. Bisaro. 2002. Comportamiento de glicinina, β-conglicinina y α-amilasa en semillas de soja deterioradas y no deterioradas. Pesquisa Agropecuária Brasileira 37: 1175-1181. [ Links ]
Schuler G., I., S. Baquero O., D. Gaona T., E. Vega G., J. Rodríguez R., C. Ramírez S., V. Nieto R. y E. Hodson J. 2005. Propagación in vitro de material seleccionado de Tabebuia rosea (Bertol.) DC. (Ocobo) y Cordia alliodora (Ruiz & Pav.) Oken (Nogal Cafetero). Revista Colombiana de Biotecnología 6: 39-50. [ Links ]
Tacoronte, M., M. Vielma, A. Mora y C. Valecillos. 2004. Propagación in vitro de caoba (Swietenia macrophylla King) a partir de yemas axilares. Acta Científica Venezolana 55: 7-12. [ Links ]
Taiz, L. and E. Zeiger. 1998. Plant physiology. Sinauer Associates, Inc. Suderland, MA, USA. 792 p. [ Links ]
Van Groenendael, J. M., S. H. Bullock and L. A. Pérez J. 1996. Aspects of the population biology of the gregarious tree Cordia elaeagnoides in a Mexican tropical deciduous forest. Journal of Tropical Ecology 12: 11-24. [ Links ]
Zhang, L., Y. Hu, S. Yan, H. Li, S. He, M. Huang and L. Li. 2012. ABA-mediated inhibition of seed germination is associated with ribosomal DNA chromatin condensation, decreased transcription, and ribosomal RNA gene hypoacetylation. Plant Molecular Biology 79:285–293. [ Links ]
Zhao, Y. and K. H. Hasenstein. 2009. Primary root growth regulation: the role of auxin and ethylene antagonists. Plant. Growth Regululation 28: 309–320. [ Links ]


