










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de ciencias forestales
versión impresa ISSN 2007-1132
Rev. mex. de cienc. forestales vol.5 no.24 México jul./ago. 2014
Artículo
Mortalidad y reclutamiento de árboles en un bosque templado de pino-encino en Jalisco
Mortality and tree recruitment in a temperate pine-oak forest in Jalisco State
Azucena Gabriela Trigueros Bañuelos1, Raymundo Villavicencio García2 y Ana Luisa Santiago Pérez2
1 Centro Universitario de Ciencias Biológicas y Agropecuarias, Universidad de Guadalajara. Correo-e: azucena_gb@msn.com
2 Departamento de Producción Forestal, Centro Universitario de Ciencias Biológicas y Agropecuarias, Universidad de Guadalajara.
Fecha de recepción: 8 de abril de 2014;
Fecha de aceptación: 8 de julio de 2014.
RESUMEN
En la dinámica forestal y los patrones sucesionales intervienen algunos procesos como la mortandad e incorporación de individuos, cuyo conocimiento fundamenta las acciones de protección y manejo de bosques. Se evaluó la mortalidad y el reclutamiento de la masa arbórea en el área natural protegida Sierra de Quila, Jalisco. En 31 sitios permanentes de monitoreo se realizaron dos mediciones: una en el año 2002 y otra en 2010, que incluyeron todos los árboles con diámetro normal ≥ 7 cm, además de los reclutados. Los códigos de mortalidad registrados fueron: muerto en pie, derribado o roto, cortado y sin rastro aparente. Se estimaron los coeficientes de mortalidad, coeficiente de reclutamiento exponencial y la vida media del bosque. En el primer muestreo se contabilizaron 918 individuos, distribuidos en 17 especies; y en el segundo 879, con 16 especies. Se obtuvo una tasa anual de mortalidad de 1.75 % y un coeficiente de mortalidad exponencial de 1.77 %. La tasa anual de reclutamiento fue 1.08 % y el coeficiente de reclutamiento exponencial 1.07 %. La vida media estimada del bosque varió entre sitios para un promedio de 45.4 años. Las diferencias entre muestras, tanto en individuos como en las tasas demográficas denotaron un desequilibrio en la dinámica poblacional del bosque.
Palabras clave: Dinámica forestal, ecología forestal, inventario, remedición, sitios permanentes, sucesión forestal.
ABSTRACT
Forest dynamics and succession patterns involve certain processes including mortality and incorporation of individuals, the knowledge of which is the basis for forest protection and management actions. The mortality and tree recruitment rates in the natural reserve of Sierra de Quila, Jalisco, were assessed. Two measurements were carried out in 31 permanent monitoring sites: one in the year 2002, and another in the year 2010. These measurements included all the trees with a normal diameter of ≥ 7 cm, apart from the recruited trees. The registered mortality codes were: standing dead, felled, cut down and dead with no apparent traces. The mortality rates, the exponential recruitment rate and the average life span of the forest were estimated. The first sampling involved 918 individuals distributed in 17 species, and the second sampling included 897 individuals distributed in 16 species. An annual mortality rate of 1.75 % and an exponential mortality rate of 1.77 % were calculated. The annual recruitment rate was 1.08 %, and the exponential recruitment rate was 1.07 %. The estimated average life span of the forest varied between sites, but the overall average was 45.4 years. The differences between the samples, for both individuals and demographic rates, indicated an imbalance in the population dynamics of the forest.
Key Words: Forest dynamics, forest ecology, inventory, remeasurement, permanent sites, forest succession.
INTRODUCCIÓN
Los bosques cuando no son expuestos a disturbios de impacto relevante mantienen un relativo equilibrio en sus funciones ecosistémicas; por el contrario, un efecto negativo tiene como consecuencia la fragmentación, deforestación y otras pérdidas intrínsecas del ecosistema. Los bosques, por lo tanto, no son estáticos, sino que están en constante cambio debido a circunstancias tanto naturales como artificiales. La dinámica se refiere a las modificaciones naturales y de eventos atípicos que suceden a través del tiempo en la composición y estructura de las masas forestales (Sheil et al., 2000; Brassard y Chen, 2010); por lo que los disturbios influyen de manera importante en la conservación de los bosques naturales.
Se especula que los sistemas clímax están, relativamente, estables o en equilibrio ecológico, pero son poco frecuentes en la naturaleza, ya que las alteraciones climáticos y las perturbaciones transforman de forma gradual los ecosistemas, lo que interrumpe, apresura o cambia el curso de la sucesión de las especies (Spurr y Barnes, 1982; Woods, 2000). Los incendios forestales, el aumento de la herbivoría, por la ganadería, y el aprovechamiento forestal irracional son algunos de los principales disturbios (Frelich, 2002).
La identificación y análisis de algunos de los procesos ecológicos complejos, sus cambios naturales y antrópicos, así como los sucesos atípicos requieren de la ejecución de investigaciones a largo plazo sobre las comunidades que integran los ecosistemas (Sheil et al., 2000). Desde el punto de vista poblacional inciden sobre la dinámica forestal: la mortalidad y el reclutamiento; razón por la cual son indicadores útiles del funcionamiento de los ecosistemas (Carey et al., 1994). La primera se describe como la cantidad de individuos que perecen en un período, mientras que el segundo alude a la capacidad de aumentar el número de árboles. El balance entre estos elementos demográficos es importante para mantener la densidad, la regeneración natural y la diversidad. La mortandad se genera por variables exógenas (enfermedades, plagas, estrés hídrico y térmico, actividades humanas) y endógenas de los individuos (competencia, decaimiento de los individuos). El reclutamiento está dado por la fecundidad, la adaptación genética y el crecimiento (Bormann y Likens, 1979; Londoño y Jiménez, 1999; Arriaga, 2000; Melo y Vargas, 2003 citado en Quinto et al, 2009).
Una manera de obtener información acerca de la supervivencia, incorporación y muerte de los árboles, necesaria para determinar la mortandad e incorporación, es el monitoreo continuo en sitios permanentes (Päivinen et al, 1994); el cual permite identificar patrones de la sucesión de especies, estado de salud y desarrollo; además, su análisis contribuye a determinar el balance entre el crecimiento, demanda poblacional y los estándares ambientales aceptables (Bellingham et al, 2000; Hernández, 2007).
De manera específica, en áreas protegidas es importante efectuar inventarios forestales continuos que proporcionen información que coadyuve a comprender los procesos de mortandad e incorporación forestal; asimismo las evaluaciones de estas tasas por periodos largos brindan una mayor confiabilidad en su estimación, y en combinación con estudios en lapsos cortos ofrecen detalles sobre causas, intensidad y patrones de muerte en un ecosistema (Kolb et al., 1994). Lo anterior es fundamental tanto para el desarrollo de planes eficientes de ordenamiento forestal, como para llevar a cabo acciones racionales en conservación, restauración y manejo sostenible de espacios protegidos (Kolb et al., 1994; Bellingham et al., 2000; Hernández, 2007). En este sentido, el objetivo del presente trabajo es calcular las tasas anuales de mortalidad y reclutamiento en bosques de pino-encino del Área de Protección de Flora y Fauna Sierra de Quila (APFFSQ), con base en la remedición de sitios permanentes de monitoreo en un intervalo de ocho años (2002-2010).
MATERIALES Y MÉTODOS
La zona de estudio se localiza en el APFFSQ, a 100 km al suroeste de la ciudad de Guadalajara, en el estado de Jalisco. Se sitúa entre 20°14' y 20°22' N, 103°56' y 104°08' O (SARH, 1993) (Figura 1). La mayor parte de la sierra es una meseta cubierta por un bosque de Pino-Encino, mientras que en su flanco sur predomina el bosque de Encino y la Selva Baja Caducifolia. Otras comunidades vegetales asociadas y de extensión reducida son el bosque de Galería, bosque Espinoso y bosque Mesófilo de Montaña (Guerrero y López, 1997). En el bosque de Pino-Encino las especies más abundantes son Quercus resinosa Liebm (41 %), Pinus lumholtzii Rob. & Fern. (15.7 %) y Pinus douglasiana Martínez (11.5 %) (Villavicencio et al, 2006).
De las microcuencas tributarias que se originan en la parte alta del área natural protegida (ANP), destaca la del río Santa Rosa, ya que su afluente es permanente y uno de los principales abastecedores de agua para el poblado de Tecolotlán (Vega et al, 2011); tiene una superficie de 14.9 km2, de la cual 92 % corresponde a un bosque templado de Pino-Encino, Selva Baja Caducifolia y bosque de Galería (Figura 1). Su clima predominante es el templado subhúmedo C (W2)(W), con lluvias en verano y con una temperatura promedio de 18.6 °C (Köppen modificado por García, 1964); la precipitación pluvial media anual es de 876 mm. La microcuenca tiene los siguientes subtipos de suelo, según la clasificación de la FAO/UNESCO: Regosol eútrico (34.2 %), Cambisol húmico (26.5 %), Feozem háplico (25.3 %), Cambisol eútrico (13.3 %) y Leptosol (0.7 %) (Villavicencio et al., 2013).
En el año 2002, Villavicencio (2004) estableció una red sistemática de 93 sitios de monitoreo permanente sobre un área piloto con una orientación de sur a norte en la parte central del APFFSQ; su área de estudio representaba los principales tipos de vegetación, suelo y estratos altitudinales. Esta red se distribuyó con rumbo franco y los centros de los lugares se localizaron a equidistancias de 400 m; en cada sitio circular de 500 m2 se registró todo el arbolado ≥ 7 cm de diámetro normal (DN, a 1.30 m).
Esa red inicial de muestreo se utilizó como la base de datos de referencia para el año 2010. Se seleccionaron 31 sitios ubicados en el bosque templado de la microcuenca del río Santa Rosa, en los cuales se remidió todo el arbolado ≥ 7 cm de DN. En cada sitio se registró el diámetro, la altura, el estado fitosanitario, la posición social de cada árbol en la masa forestal y otros parámetros ambientales. Se cuantificaron los árboles muertos y los reclutados que alcanzaron el diámetro mínimo. De acuerdo a Londoño y Jiménez (1999) se definieron los siguientes códigos para clasificar la condición observada de muerte de un árbol: muerto en pie, derribado o roto, cortado y sin rastro aparente. Con la finalidad de evaluar el proceso demográfico se estimaron las tasas de reclutamiento y mortalidad mediante los modelos matemáticos descritos por Quinto et al. (2009); así como la vida media del bosque y el tiempo de duplicación con las expresiones referidas por Aguilar y Reynel (2011).
Tasa anual de mortalidad. Calcula la mortandad de los árboles por sitio de muestreo:
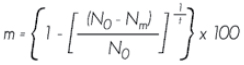
Donde:
m = Tasa anual de mortalidad expresada en porcentaje
N0 = Número de árboles al principio del período
Nm = Número de árboles muertos
t = Tiempo transcurrido entre los dos inventarios
Coeficiente de mortalidad exponencial. Evalúa el decremento poblacional por sitio de muestreo:
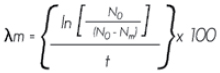
Donde:
λm = Coeficiente de mortalidad exponencial expresada en porcentaje
ln = Logaritmo neperiano
N0 = Número de árboles al principio del período
Nm = Número de árboles muertos
t = Tiempo transcurrido entre los dos inventarios
Tasa anual de reclutamiento. Estima la incorporación de árboles por sitio de muestreo:
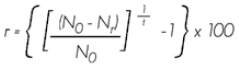
Donde:
r = Tasa anual de reclutamiento expresada en porcentaje
N0 = Número de árboles al principio del período
Nr = Número de árboles muertos
t = Tiempo transcurrido entre los dos inventarios
Coeficiente exponencial de reclutamiento. Deduce el incremento poblacional por sitio de muestreo:

Donde:
λr = Coeficiente de mortalidad exponencial expresada en porcentaje
ln = Logaritmo neperiano
N0 = Número de árboles al principio del período
Nr = Número de árboles reclutados
t = Tiempo transcurrido entre los dos inventarios
Vida media del bosque. Se refiere al tiempo necesario para que la población en la muestra inicial disminuya a la mitad con la tasa actual de mortalidad; la expresión se interpreta en años:
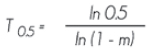
Donde:
T0.5 = Vida media del bosque
ln = Logaritmo neperiano
m = Tasa anual de mortalidad
Tiempo de duplicación. Es el tiempo requerido para que la población se duplique manteniendo la presente tasa de reclutamiento, la expresión se interpreta en años:

Donde:
T2 = Vida media del bosque
ln = Logaritmo neperiano
r = Tasa anual de reclutamiento
Se realizó un análisis de ANOVA de un factor para comparar la varianza entre cada medición (2002 y 2010) de los parámetros diámetro, altura y área basal por grupo de especies. La hipótesis nula probada para los parámetros DN, altura y área basal por grupo de especies se realizó con 90 % (alfa - 0.1) de intervalo de confianza. La hipótesis por aceptar o rechazar fue: Ho: m1=m2, no existe cambio significativo entre los años de muestreo del diámetro, altura y área basal.
RESULTADOS Y DISCUSIÓN
Composición de especies y parámetros dasométricos. En el año 2002 se registraron 918 individuos con un DN ≥ 7 cm, pertenecientes a 17 especies; mientras que en 2010 se midieron 879 árboles distribuidos en 16 taxa. El único taxón no registrado en 2010 fue Crataegus pubescens (Kunth) Steud.; cuya pérdida (3 individuos) se debió a causas fitosanitarias (Cuadro 1). En el transcurso de ocho años se observó un decremento demográfico de 4.2 %, ya que en 2002 la densidad era de 574 árboles ha-1 y en 2010 disminuyó a 549 individuos ha-1. En este período, las especies que redujeron su densidad fueron siete y las tres principales: P. lumholtzii, Quercus coccolobifolia Trel. y Arbutus xalapensis HBK.
Con respecto a la densidad del arbolado, los pinos como P. lumholtzii, P. douglasiana Martínez, P. devoniana Lindl. y P. oocarpa Scheide var. oocarpa contribuyeron con 44.9 % de los individuos en 2010 (Cuadro 1). En particular, los tres primeros son más abundantes debido a que en la zona predomina una geoforma en laderas cóncavas, donde se favorece la humedad en el suelo. En el caso de P. oocarpa suele observarse en sitios con laderas convexas, con menor aporte de humedad. Aspecto que concuerda con lo referido por Jardel et al. (2004) para bosques mixtos de pino-latifoliadas y bosques de pino en la Sierra de Manantlán.
Pinus douglasiana y P. lumholtzii tuvieron valores similares en alturas; no obstante, P. douglasiana obtuvo una mayor ventaja en la competencia debido a su incremento diamétrico, y como consecuencia en el área basal (Cuadro 1). En los encinos, Quercus resinosa Liebm. fue dominante junto con los pinos, seguido por Quercus coccolobifolia. En contraste con la abundancia, Quercus obtusata Bonpl., Quercus castanea Née, Quercus candicans Née y Quercus crassifolia Bonpl. se presentaron en menor proporción (Cuadro 1). Con base en el estudio y mapa de vegetación generado para el área protegida por Villavicencio (2004), Quercus resinosa es un elemento dominante en el bosque de pino-encino (41 %) distribuido en el intervalo altitudinal de 2 000 a 2 300 m. En tanto, Q. coccolobifolia, Q. obtusata y Q. castanea corresponden, en su distribución altitudinal más acotada, al bosque de pino-encino entre 2 000 a 2 200 m; por otro lado, Q. candicans y Q. crassifolia se desarrollan en bosque de pino-encino en cotas superiores (2 200 a 2 450 m) o en cañadas con remanentes similares al bosque mesófilo de montaña (Guerrero y López, 1997).
En el Cuadro 2 se resumen los datos descriptivos en composición y estructura por grupo de especies (pinos, encinos, otras hojosas). Respecto a la abundancia (N ha-1) y dominancia (g m2 ha-1) destacan los pinos con 46.2 y 57.5 %, respectivamente en el inventario de 2002, y con 44.9 y 60.7 % en el 2010.
En el análisis de varianza para el grupo de especies de pinos se concluyó que durante el tiempo transcurrido entre los dos inventarios, el cambio en DN, altura y área basal fue significativo estadísticamente, ya que los valores F de los tres parámetros dasométricos fueron superiores a sus respectivos valores críticos de F, por consiguiente la hipótesis se rechazó (Cuadro 3).
La hipótesis nula de los parámetros dasométricos para los encinos se aceptó, porque no hubo una diferencia significativa; es decir, los valores de F estuvieron por debajo del valor crítico (Cuadro 4). En específico, el análisis para la especie dominante, Quercus resinosa, se ajustó con lo esperado para el conjunto de encinos, en la que los valores de F para el diámetro normal, altura y área basal (0.714, 0.230 y 0.871) no fueron superiores al valor crítico (2.714). Lo anterior se refleja en las diferencias obtenidas entre los muestreos de estos parámetros, en comparación con las estimaciones para el grupo de los pinos.
El cambio en el promedio de DN y altura de los pinos fue de 2.2 cm y 2 m; en contraste, para los encinos fue menor (0.5 cm y 0.3 m). En relación con la densidad del arbolado, Quercus resinosa presentó el mayor número de individuos en ambos inventarios (197, en 2002 y 205 árboles ha-1, en 2010), así como el total del grupo de encinos (283 árboles ha-1), mientras que el grupo de los pinos disminuyó de 274 a 255 individuos ha-1; valores que podrían relacionarse con los diferentes atributos intrínsecos de ambos tipos de árboles.
En el APFF La Primavera, a pesar de los frecuentes incendios forestales, las poblaciones de los encinos mantienen su sobrevivencia; por lo tanto, el daño ocasionado por el fuego y el aumento de la temperatura del suelo estimula en algunas especies su capacidad de rebrotamiento, tal como sucede en Quercus resinosa, lo cual evidencia su gran capacidad adaptativa a este tipo de disturbio (Arias et al., 2009).
En relación con su distribución altitudinal y ecología en la Sierra Madre Occidental (SMOC): Quercus resinosa ha sido clasificada como especie pionera, debido a sus hojas coriáceas, rígidas, gruesas y deciduas, que le confieren resistencia ante la evapotranspiración; asimismo su porte bajo le permite un rápido establecimiento (Flores, 2007). Quercus coccolobifolia, también es pionera, se establece en claros y aunque su distribución altitudinal es inferior a la de Q. resinosa, en la zona de la MRSR, es abundante (55 árboles ha-1 Flores (2007) efectuó un análisis sobre la dinámica de encinos en diferentes sitios de las sierras Madre Oriental y Occidental en diferentes tipos de cobertura, altitud y grado de disturbio; el autor sustentó que en la sucesión natural de Quercus resinosa y Q. coccolobifolia se reemplazan por especies intermedias (Q. rugosa, Q. obtusata y Q. crassifolia), y estas, a su vez, por las tardías (Q. laurina y Q. uxoris); asimismo el patrón de distribución que conllevan depende de la altura. Igualmente Quercus coccolobifolia se clasifica como un taxon pionero por su porte bajo y tamaño pequeño de bellota, el cual le facilita tener una mayor dispersión en las áreas abiertas (Castillo et al., 2008).
Las especies colonizadoras presentan hojas más grandes, gruesas y son más intolerantes a la sombra, que otros taxa de hojas delgadas y de tamaño pequeño (Castillo et al., 2008). En específico para la Sierra de Quila, Guerrero y López (1997) distinguen la distribución de los encinos en dos estratos: el primero, donde los de hojas duras y coriáceas se ubican a menores altitudes y baja humedad (Quercus resinosa y Q. magnoliifolia); el segundo, en el cual los encinos de hojas pequeñas y delgadas se distribuyen en los lugares altos (Q. crassifolia y Q. candicans).
En el área de estudio, el grupo de otras hojosas tampoco tuvo diferencias significativas, por su baja frecuencia en los sitios (5.8-5.2 %) (Cuadro 5). Los taxa pertenecientes a la familia Ericaceas: Arbutus xalepensis Kunth, A. glandulosa M. Martens & Galeotti, son susceptibles a las plantas de la familia Loranthaceae que afecta su crecimiento o causa su muerte (González, 1990). Guerrero y López (1997) especifican que Comarostaphylis glaucescens (Kunth) Zucc. ex Klotzsch suele desarrollarse en claros erosionados o al lado de los caminos.
Mortalidad de árboles y reclutamiento. En el periodo 2002-2010 murieron 121 árboles, con un promedio anual de 15; el reclutamiento en el mismo lapso correspondió a 82, para un promedio de 10 por año. La tasa de mortalidad fue de 1.75 % año-1 y la de reclutamiento 1.08 % año-1; este valor se expresa en la cantidad de individuos por hectárea muertos y reclutados (78 y 53) (Cuadro 6).
La vida media del bosque es de 45.4 años, tiempo para que la población, con la tasa de mortalidad actual, se reduzca a la mitad; el tiempo de duplicación se calculó en 74 años (Cuadro 6). Ramírez et al. (2002) analizaron la mortalidad y reclutamiento de árboles en seis parcelas sin disturbio de un bosque nublado de Venezuela, a pesar de que la estimación de vida media proyectada para cada parcela varió, dedujeron un promedio de 43.21 años. En el parque estatal Fontes do Ipiranga se registran 41.23 años como vida media del bosque y la de duplicación en 20.39 años, esto debido a que las tasas demográficas para el total de la población son de 1.67 % año-1 para la mortalidad y la de reclutamiento 3.47 % año-1 (Gomes et al., 2003). En esa región de Brasil, la sucesión de especies tolerantes a la sombra es rápida y la muerte de árboles dominantes abren espacios del dosel donde otras especies leñosas se regeneran de inmediato (Gomes et al., 2003).
Los valores de los coeficientes de mortalidad y reclutamiento exponencial fueron muy similares a los de las tasas; el primero fue 1.77 % y el segundo 1.07 %; cifras que se compararon con estudios efectuados en ecosistemas forestales con fines de conservación, manejo o bien con algún grado de disturbio. Al respecto, las tasas anuales de mortalidad y reclutamiento estimadas, en un bosque primario tropical maduro, fueron muy similares a las de la presente investigación: 1.39 % año-1 y 1.2 % año-1 (Quinto et al., 2009).
Ramírez et al (2002) obtuvieron, en un bosque nublado de los andes venezolanos, tasas de 1.59 y 1.65 % año -1 de mortandad y reclutamiento, los cuales están dentro de lo normal para bosques maduros expuestos a pequeños disturbios naturales. Sin embargo, el mismo autor señala que para ponderar estas estimaciones es necesario realizar más estudios sobre las causas de mortalidad; si son inducidas o naturales dependerá que la tasa sea alta o baja. En dos parcelas permanentes de monitoreo en un bosque maduro sin afectaciones antrópicas, Londoño y Jiménez (1999) consignan tasas anuales de mortalidad y reclutamiento inferiores a las aquí documentadas.
En bosques productivos, la mortalidad por manejo silvícola aumenta; tal como Aguilar et al. (2006) lo exponen para parcelas manejadas con tasa de mortalidad de 0.72 a 5.02 % año-1 y tasa de reclutamiento de 0.01 a 3.45 % año-1; mientras que aquellas sin manejo, registraron valores de 0.25 a 2.62 % año-1 para la tasa de mortalidad; para la de reclutamiento de 0.62 a 1.98 % año-1. Lugo y Scatena (1996) indican que una tasa anual de mortalidad menor a 5 % se considera como natural o intrínseca por su baja intensidad, si el porcentaje anual es superior, entonces es catastrófica.
La distribución diamétrica de los árboles muertos (121) y reclutados (82) se agrupan en clases de 5 cm de DN (Figura 2). En la clase diamétrica de 10 cm ocurrió la mayor mortalidad, con 53 individuos (43.8 %), lo cual se debe a un proceso natural de aclareo (Carey et al. 1994; Lugo y Scatena 1996); Smith et al. (2005) mencionan que en bosques mixtos de coníferas, la densidad del rodal es una causa de mortalidad.
De los arboles reclutados, 50 % (41 individuos) se incorporó a la clase diamétrica de 10 cm, ya que en 2002 estos árboles estaban próximos a efectuar dicho cambio (Figura 2); por otro lado, la clase correspondiente a 5 cm tuvo un patrón inverso; es decir, el número de individuos muertos fue menor que los reclutados; los primeros fueron latifoliadas como Quercus resinosa, Arbutus glandulosa y A. xalapensis, (muertos en pie); mientras que los árboles reclutados incluyen encinos: Quercus resinosa (15) y Q. coccolobifolia (3); otras hojosas: Clethra hartwegii Britton (4) y Arbutus xalapensis (2); y pinos, Pinus lumholtzii (2) y P. devoniana (1). Lo anterior evidencia una regeneración baja en pinos y mayor en encinos; en particular, la especie más abundante fue Q. resinosa. Sucesión que concuerda con lo que refieren Jardel et al. (2004), quienes consignan que los encinos y los madroños presentan tolerancia intermedia a la sombra, se desarrollan en el sotobosque, alcanzan el dosel y su densidad aumenta cuando no existen disturbios; sin embargo, los pinos que tienen requerimientos más altos de luz, los favorecen los incendios que abren claros en el dosel.
Un factor determinante en la competencia es la tolerancia a la sombra, ya que algunas latifoliadas son más tolerantes que los pinos, por lo que la tendencia, conforme la sucesión avanza en condiciones de mayor cobertura de dosel sería de pino a encino (Oliver y Larson, 1990; Jardel et al, 2004). Esto no sucedería en presencia de disturbios de gran escala, como ocurre en los bosques templados de Sudamérica; que al tener catástrofes naturales, como inundaciones, terremotos, incendios y deslaves las especies intolerantes a la sombra tienden a dominar (Veblen, 2005).
El porcentaje más alto (48.8 %) de los árboles muertos se concentró en tres sitios: 47, 52 y 59, los cuales registraron una tasa anual de mortalidad de 6.3, 7.1 y 10.4 % año-1, respectivamente (Figura 3). El código de mortalidad en esos sitios fueron: árboles muertos cortados, con 64.4 % (MC), sin rastro aparente con 20.3 % (M?); en pie, con 8.5 % (MP); y derribados, con 6.8 % (MD) (Figura 3); en el resto de los sitios la tasa de mortalidad varió de 0 a 2.5 % año-1. Las causas de mortandad en esos sitios fueron MP (50 %), MD (21 %), M? (16.1 %) y MC (12.9 %). Cabe destacar que de excluir los porcentajes obtenidos del cálculo de las tasas demográficas de los sitios de muestreo 47, 52 y 59; el valor para la tasa anual de mortalidad disminuiría en 1.03 % año -1; y para la de reclutamiento sería 0.95 % año -1. Por lo tanto, la incorporación sería superior a la mortandad, como se esperaría en bosques preservados con disturbios naturales, y denotaría un estado de estabilidad en su dinámica. De acuerdo a estos valores demográficos, la vida media del bosque sería de 66.4 años y el tiempo de duplicación de 71.3 años.
Al respecto, los árboles muertos cortados y sin rastro aparente se vincularon con algunas acciones de manejo fitosanitario entre el año 2002 y el 2007. Las áreas de intervención incluyeron el espacio de los sitios de muestreo 47, 52 y 59, los cuales concentraron el porcentaje más alto de árboles cortados. Las cortas de saneamiento se hicieron por la presencia del descortezador Dendroctonus mexicanus Hopkins; cuyos hospederos son Pinus douglasiana, P. oocarpa, P. devoniana y P. lumholtzii, otras especies pudieron haber sido afectadas por daños directos o indirectos asociados a las mismas actividades de manejo (Comité Regional de la Sierra de Quila, 2007).
Las especies de árboles muertos de mayor frecuencia en toda el área de estudio fueron, en orden descendente: Pinus lumholtzii (40.5 %), Quercus resinosa (20.7 %), Q. coccolobifolia (10.7 %), P. douglasiana (9.1 %), Arbutus xalapensis (6.6 %), A. glandulosa (3.3 %), Crataegus pubescens (3.3 %), Q. castanea (2.5 %), así como Clethra hartwegii, P. montezumae, P. oocarpa, Prunus serotina, Q. candicans con 0.8 % para cada una.
El registro de mortandad con porcentaje más alto correspondió al de muerto cortado con 38 (46 árboles), seguido de muertos en pie, con 29.8 % (36 individuos); muerto sin rastro aparente, con 18.2 % (22); y muerto derribado con 14 % (17) (Figura 4).

De acuerdo con Carey et al. (1994), la forma de explicar la mortandad de los arboles es compleja, pues intervienen una extensa combinación de elementos, muchos de ellos difíciles de identificar. Algunos autores como Lugo y Scatena (1996) sugieren que en la mayoría de los árboles de clases diamétricas y alturas superiores ocurre una senescencia producida por factores endógenos, como la genética y la edad.
Carey et al. (1994) coinciden en que los árboles que mueren en pie, y no muestran incremento anterior a su decaimiento responde a variables intrínsecas como debilitamiento por enfermedades, o estrés ambiental (competencia, falta de nutrientes, sequía). Estos individuos de grandes clases también caen con frecuencia, lo que puede derivar en la muerte inmediata de otros adyacentes de dimensiones menores. Por lo anterior, se distinguen dos grupos de mortandad: la ocasionada por acciones de origen humano, y la natural o intrínseca.
Las cortas de saneamiento incluyen el arbolado muerto cortado y muerto sin rastro aparente, categorías en las que sobresalen, por el número de individuos, las especies con mayor cantidad MC: P. lumholtzii y Q. resinosa con 19 y 13, respectivamente; y como M?, P. lumholtzii (13 árboles) y Q. coccolobifolia (cuatro árboles). En general, los árboles cortados y sin rastro aparente tuvieron promedios en diámetros de 20.8 y 16.4 cm y alturas promedio de 12.3 y 10.7 m. En relación con las condiciones cuyo origen se debe a factores naturales o intrínsecos, las especies con un alto número de árboles como muertos derribados fueron P. lumholtzii y P. douglasiana con 9 y 4, respectivamente; y como muertos en pie Q. resinosa y P. lumholtzii, con 13 y 8 individuos. El pino triste (Pinus lumholtzii) registró una mortandad superior, contrario a su regeneración (Cuadro 7).
Las especies reclutadas con frecuencia más alta fueron, en orden descendente: Quercus resinosa (43.9 %), Pinus devoniana (15.9 %), P. douglasiana (11 %), P. lumholtzii (9.8 %), Q. coccolobifolia (6.1 %), Clethra hartwegii (4.9 %) y P. oocarpa (2.4 %). En total se calcularon 53 árboles ha -1 reclutados, con un promedio anual de 7 árboles ha -1, que en su conjunto suman un incremento en el área basal de 0.549 m2 ha -1 (Cuadro 8).
Algunas de las causas por las que las especies de reclutamiento responden a una mayor o menor dominancia han sido citadas por González (1986), quien menciona que al igual que en otras regiones de Jalisco, donde las especies de encino son las dominantes del dosel, en bosques mixtos de pino y encino o encino, Quercus resinosa es una de las más características y extendidas en el estado y puede dominar desde los 1 300 hasta los 2 500 m de altitud, generalmente, en hábitats más secos. De acuerdo a Villavicencio (2004) y Santiago y Villavicencio (2007), Quercus resinosa es dominante tanto en el APFF Sierra de Quila como La Primavera, por su estrategia reproductiva asexual mediante rebrotes en los tocones o bien de raíz, que favorecen su alta frecuencia y densidad de plántulas. Por el contrario, Quercus coccolobifolia, Clethra hartwegii, Arbutus xalapensis y Comarostaphylis glaucescens suelen ser elementos florísticos más restringidos en su distribución, tanto en este estudio como en las áreas antes citadas.
Guerrero y López (1997) señalan la distribución de las especies en Sierra de Quila: Q. coccolobifolia es preferente en la cota de altitud de 1 850 a 2 000 m, y la refieren como poco abundante; C. hartwegii es un elemento escaso dentro de cañadas en zonas húmedas; A. xalapensis es más característico en sitios por arriba de los 2 100 m; y C. glaucescens se considera como parte del conjunto de arbustos secundarios típicos de suelos erosionados y las márgenes de los caminos.
Por su parte, Villavicencio et al. (2012) registraron que el promedio de plántulas de especies arbóreas en Sierra de Quila varía entre fragmentos de bosque de diferente tipo (densidad de cobertura abierta y cerrada) y tamaño (10-50 y > 50 ha), en los cuales se confirma un patrón similar al comportamiento de las registradas con mayor reclutamiento en este trabajo. Al comparar los taxa comunes en ambos estudios se observó que Q. resinosa es abundante en cualquier condición (tamaño de fragmento y densidad de cobertura), C. hartwegii es escasa y ocurre principalmente sobre las cañadas, y A. xalapensis se encuentra en mayor proporción en bosques abiertos y mayores a 50 ha. Al respecto, Figueroa y Olvera (2000) hacen referencia al comportamiento en patrones de regeneración en bosques mixtos de encino de la Sierra de Manantlán, los cuales mostraron asociación con las condiciones de hábitat locales como exposición, altitud, topografía, tipo de dosel e intensidad de pastoreo, como las principales variables que explican las diferencias en composición de especies entre las asociaciones de plántulas.
Olvera et al. (2010) distinguieron una diferenciación ambiental en la distribución de las especies de encino con relación a requerimientos ecológicos similares en gradientes de zona de humedad (xérico a húmeda), altitud y heterogeneidad del micro hábitat como posición topográfica y madurez del dosel.
CONCLUSIONES
En el inventario de 2002 se identificaron 17 especies distribuidas en seis familias y ocho géneros; en 2010 disminuyó a 16 especies; asimismo la densidad del arbolado se redujo en 4.2 %, la mayor abundancia de árboles muertos se presentan en Pinus lumholtzii, Quercus resinosa y Q. coccolobifolia, mientras que las especies reclutadas de mayor frecuencia corresponden a Quercus resinosa, Pinus devoniana y P. douglasiana., la primera de estas, es una especie más tolerante a la sombra y se beneficia de la apertura del dosel, lo que explica su elevada densidad y regeneración.
A pesar del descenso poblacional del grupo de pinos tuvieron diferencias estadísticamente significativas en el diámetro, altura y área basal; en contraste con los encinos, la población aumentó, sin embargo no presentaron cambios significativos sobre los mismos parámetros.
El balance de la dinámica poblacional del bosque muestra un desequilibrio, que se refleja en las tasas demográficas de mortalidad (1.75 %) y reclutamiento (1.08 %).
La especie con mayor número de árboles reclutados fue Quercus resinosa, la cual registró un aumento poblacional de 2.8 % entre 2002 y 2010, además 43.9 % de los árboles incorporados pertenecen a este taxon. En contraste, entre la población más abundante en el bosque templado, la segunda instancia la ocupó el pino triste (Pinus lumholtzii), el cual evidenció un decremento poblacional mayor que sus árboles reclutados entre 2002 y 2010.
Los códigos de mortandad permiten diferenciar causas generadas de forma natural, de las antrópicas; en este sentido, los 121 árboles registrados como muertos se dividieron en dos grupos: los originados por factores antropogénicos, que coinciden con los códigos de muerto cortado y sin rastro aparente y los de origen natural, clasificados como: muertos en pie y derribados. Las causas que originaron su muerte correspondieron a factores exógenos y endógenos, del ambiente o de los mismos individuos.
AGRADECIMIENTOS
El proyecto fue financiado por la Universidad de Guadalajara (P3E2010 -107971 y P3E2011 -123405). Al Comité Regional de Protección, Promoción y Fomento de los Recursos Naturales de la Sierra de Quila, A. C. por las facilidades brindadas en la realización de este estudio. A todos los brigadistas del Comité Regional por los apoyos otorgados durante los muestreos de campo. Así como a los estudiantes del Centro Universitario de Ciencias Biológicas y Agropecuarias de la Universidad de Guadalajara, quienes otorgaron su tiempo y apoyaron con entusiasmo la remedición de los sitios.
REFERENCIAS
Aguilar, M. y C. Reynel. 2011. Dinámica forestal y regeneración de un bosque montano nublado de la selva central del Perú. Asociación Peruana para la Promoción del Desarrollo Sostenible. Universidad Nacional Agraria. La Molina. Lima, Perú. 165 p. [ Links ]
Aguilar, S., M. Mazá, A. Hernández y G. Cifuentes. 2006. Desarrollo de un sistema de parcelas forestales permanentes de muestreo en Guatemala. Ministerio de Agricultura Ganadería y Alimentación (MAGA). Guatemala, Guatemala. 146 p. [ Links ]
Arias, J., A. Gallegos, R. Cabrera, D. Sánchez, G. González, J. Castañeda, y A. Mora. 2009. Respuesta de poda de brotes postincendio en Quercus resinosa Liebm. y Q. viminea Trel. (Fagaceae) en el Bosque La Primavera, Jalisco, México: Análisis preliminar. II Foro de Investigación y Conservación del "Bosque La Primavera". 5 - 6 de marzo de 2009. Zapopan, Jal. México. 10 p. [ Links ]
Arriaga, L. 2000. Types and causes of tree mortality in a tropical montane cloud forest of Tamaulipas, Mexico. Journal of Tropical Ecology. 16(5):623-636. [ Links ]
Bellingham, P., S. Wiser, D. Coomes and A. Dunningham. 2000. Review of permanent plots for long-term monitoring of New Zeland's indigenous forests. Science for Conservation 151. Department of Conservation. Wellington, New Zealand. 75 p. [ Links ]
Bormann, F. H. and G. E. Likens. 1979. Pattern and process in a forested ecosystem. Springer-Verlag. New York, NY. USA. 253 p. [ Links ]
Brassard, B. W. and H. Y. Chen. 2010. Stand structure and composition dynamics of boreal mixed wood forest: Implications for forest management. Sustainable Forest Management Network. Edmonton, AB. Canada.19 p. [ Links ]
Carey, E. V., S. Brown, A. J. Gillespie and A. E. Lugo.1994. Tree mortality in lowland tropical moist and tropical lower montane moist forests of Venezuela. Biotropica. 26(3):255-265. [ Links ]
Castillo L., P., J. A. Flores C., J. R. Aguirre R. and R. I. Yeaton H. 2008. Dinámica sucesional del encinar de la sierra de Álvarez, San Luis Potosí, México. Madera y Bosques. 14(1):21-35. [ Links ]
Comité regional de la Sierra de Quila. 2007. Informe General de Saneamientos Forestales realizados dentro del área natural protegida "Sierra de Quila" Ciclo(s) 2002-2007. Comité Regional de Protección, Promoción y Fomento de los Recursos Naturales de la Sierra de Quila, A.C. Tecolotlán, Jal. México. 7 p. [ Links ]
Figueroa R., B. L. y M. Olvera V. 2000. Regeneration patterns in relation to canopy species distribution and site variables in mixed oak forests in the Sierra de Manantlán Biosphere Reserve, Mexico. Ecological Research 15:249-261. [ Links ]
Flores C. J. A. 2007. Análisis de gradiente y dinámica sucesional de bosques de encino (Quercus) en las sierras Madre Oriental y Occidental de México. Tesis de Doctorado. Facultades de Ciencias Químicas, Ingeniería y Medicina, Universidad Autónoma de San Luis Potosí. San Luis Potosí, SLP. México. 95 p. [ Links ]
Frelich, L. E. 2002. Forest dynamics and disturbance regimes: Studies from temperate-evergreen deciduous forests. Cambrige University Press. West Nyack, NY. USA. 278 p. [ Links ]
García, E. 1964. Modificaciones al sistema de clasificación climática de Köppen para adaptarlo a las condiciones de la República Mexicana. Ed. Offset Larios. Guadalajara, Jal. México. 69 p. [ Links ]
Gomes, E. P., W. Mantovani and P. Y. Kageyama. 2003. Mortality and recruitment of trees in a secondary montane rain forest in southeastern Brazil. Brazilian Journal of Biology. 63(1):47-60. [ Links ]
González V., L. M. 1986. Contribución al conocimiento del género Quercus (Fagaceae) en el estado de Jalisco. Colección Flora de Jalisco. Instituto de Botánica, Universidad de Guadalajara. Guadalajara, Jal. México. 241 p. [ Links ]
González V., L. M. 1990. Las Ericáceas de Jalisco, México. Colección Flora de Jalisco. Instituto de Botánica, Universidad de Guadalajara. Guadalajara, Jal. México. 141 p. [ Links ]
Guerrero N., J. J. y G. A. López C. 1997. La vegetación y la flora de Sierra de Quila. Universidad de Guadalajara. Guadalajara, Jal. México. 134 p. [ Links ]
Hernández D., F., F. Huerta O., B. Barrios D. y M. A. Posadas G. 2012. Análisis dasométrico y propuesta de ordenamiento agroforestal del bosque en Tetela de Ocampo, Puebla. Avances en Investigación Agropecuaria. 16(3): 75-82. [ Links ]
Hernández L., I. 2007. Cambios en la estructura y composición del bosque bajo dos tratamientos silviculturales en la Comunidad de Capulálpam de Méndez, Ixtlán, Oaxaca, México. Tesis de maestría. Programa de Educación para el Desarrollo y la Conservación. Centro Agronómico Tropical de Investigación y Enseñanza (CATIE). Turrialba, Costa Rica. pp. 5-20. [ Links ]
Jardel P. E. J., A. L. Santiago P., C. Cortés M., y F. Castillo N. 2004. Sucesión y dinámica de rodales. In: Cuevas G., R. y E. J. Jardel P. 2004. Flora y Vegetación de la Estación Científica Las Joyas. Universidad de Guadalajara. Guadalajara, Jal. México. pp. 179-203. [ Links ]
Kolb, T. E., M. R. Wagner and W. W. Covington. 1994. Concepts of forest health: Utilitarian and ecosystem perspectives. Journal of Forestry. 92:10-15. [ Links ]
Londoño, A. C. y E. M. Jiménez. 1999. Efecto del tiempo entre los censos sobre la estimación de las tasas anuales de mortalidad y de reclutamiento de árboles (períodos de 1, 4 y 5 años). Crónica Forestal y del Medio Ambiente. 14(1):41-58. [ Links ]
Lugo, A. E. and F. N. Scatena.1996. Background and catastrophic tree mortality in tropical moist, wet, and rain forests. Biotropica. 28(4a):585-599. [ Links ]
Milena, S. y G. Venegas. 1999. Tablas de volumen comercial con corteza para encino, roble y otras especies del bosque pluvial montano de la Cordillera de Talamanca, Costa Rica. (CATIE). Serie Técnica. Informe Técnico Núm. 306. pp. 10-13. [ Links ]
Oliver, C.D. and Larson, B.C. 1996. Forest stand dynamics. John Wiley and Sons, New York, NY. USA. 520 p. [ Links ]
Olvera V., M., B. L. Figueroa R. y J. M. Vázquez L. 2010. Is there environmental differentiation in the Quercus dominated forests of west-central Mexico? Plant Ecology 211:321-335. [ Links ]
Organización de las Naciones Unidas para la Agricultura y la Alimentación. (FAO). 2011. Situación de los bosques del mundo. Roma, Italia. 176 p. http://www.fao.org/docrep/013/i2000s/i2000s.pdf (22 de noviembre de 2013). [ Links ]
Päivinen, R., H. Gyde L., S. Poso y T. Zawila-Nieszwiecki. 1994. Directrices internacionales de IUFRO para la monitorización de los recursos forestales. Unión Internacional de Institutos de Investigación Forestal. IUFRO World Series Vol. 5. Viena, Austria. pp. 55-97. [ Links ]
Quinto M., H., R. Rengifo I. y Y. A. Ramos P. 2009. Mortalidad y reclutamiento de árboles en un bosque pluvial tropical de Chocó (Colombia). Revista Facultad Nacional de Agronomía - Medellín. 62(1), 4855-4868. [ Links ]
Ramírez, H., A. Torres y J. Serrano. 2002. Mortalidad y reclutamiento de árboles en un bosque nublado de la cordillera de los Andes, Venezuela. Ecotropicos 15(2): 177-184. [ Links ]
Santiago P., A. L. and R. Villavicencio G. 2007. Multiple analyses of environmental and structural variables to characterization of Quercus Pinus forests in the La Primavera, Jalisco, Mexico. In: The Annual Meeting of the Association for Tropical Biology and Conservation. 15 al 19 de julio del 2007. Morelia, Mich. México. 381 p. [ Links ]
Secretaría de Agricultura y Recursos Hidráulicos (SARH). 1993. Estudio de la flora y fauna de la zona protectora forestal Sierra de Quila. SARH-SFF. Guadalajara, Jal. México. 69 p. [ Links ]
Sheil, D., S. Jennings and P. Savill. 2000. Long-term permanent plot observations of vegetation dynamics in Budongo: a Ugandan rain forest. Journal of Tropical Ecology. 16:765-800. [ Links ]
Smith, T. F., D. M. Rizzo and M. North. 2005. Patterns of mortality in an old-growth mixed-conifer forest of the southern Sierra Nevada, California. For. Sci. 51:266-275. [ Links ]
Spurr, S. H. y B. V. Barnes. 1982. Ecología Forestal. A. G. T. Editor. México D.F. México. 690 p. [ Links ]
Veblen, T. T., J. J. Armesto, B. R. Burns, T. Kitzberger, A. Lara, B. León, and K. R. Young. 2005. The coniferous forests of South America. In: Andersson, F. and S. Gessel (eds.). Ecosystems of the World: Coniferous Forests. Elsevier. Amsterdam, Netherlands. 633 p. [ Links ]
Vega R., D., R. Villavicencio G., A. L. Santiago P., J. M. Chávez A. y S. L. Toledo G. 2011. La capacidad de infiltración de agua de la microcuenca del río Santa Rosa de la Sierra de Quila, y su valoración al pago como un servicio ambiental. In: Villavicencio G., R., A. L. Santiago P., V. C. Rosas E. y L. Hernández L. (cords.). 2011. I Foro de conocimiento, uso y gestión del Área Natural Protegida Sierra de Quila. 11 y 12 de noviembre de 2011. Universidad de Guadalajara. Tecolotlán, Jal. México. pp. 22-26. [ Links ]
Villavicencio G., R. 2004. Kartierung von Vegetationsstrukturen und deren Veränderung in Naturschutzgebieten mit Hilfe von Fernerkundung und terrestrische Inventurverfahren. Dargestellt am Beispiel des Schutzgebietes für Flora und Fauna Sierra de Quila im Bundesstaat Jalisco im Westen Mexikos. Cuvillier Verlag, Göttingen. Deutschland. 161 p. [ Links ]
Villavicencio G., R., A. L. Santiago P., J. J. Godínez H., J. M. Chávez A. y S. L. Toledo G. 2012. Efecto de la fragmentación sobre la regeneración natural de la Sierra de Quila, Jalisco. Revista Mexicana de Ciencias Forestales 3(11): 9-23. [ Links ]
Villavicencio G., R., P. Bauche P., A. Gallegos R., A. L. Santiago P. y F. M. Huerta M. 2006. Caracterización estructural y diversidad de comunidades arbóreas de la Sierra de Quila. Ibugana. 13(1) 67-76. [ Links ]
Villavicencio G., R., S. L. Toledo G., R. Ávila C., A. L. Santiago P. y B. Martínez R. 2013. Áreas potenciales de infiltración en tres microcuencas de la Sierra de Quila, estado de Jalisco In: Méndez G., J., A. Zarate L. y H. D. González L. (comp.). Memorias de resúmenes del XI Congreso Mexicano sobre Recursos Forestales. 1 al 4 de octubre de 2013. Somerefo/Universidad Autónoma Agraria Antonio Narro. Saltillo, Coah. México. 297 p. [ Links ]
Woods, K. D. 2000. Long-term change and spatial pattern in a late-successional hemlock-nothern hardwood forest. J. Ecol. 88:267-282. [ Links ]

