










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de ciencias forestales
versión impresa ISSN 2007-1132
Rev. mex. de cienc. forestales vol.4 no.19 México sep./oct. 2013
Artículos
Variación fenotípica de poblaciones naturales de Pinus oocarpa Schiede ex Schltdl. en Chiapas
Phenotypic variation of natural Pinus oocarpa Schiede ex Schltdl. populations in Chiapas
Benito N. Gutiérrez Vázquez1, Martín Gómez Cárdenas2, Mario H. Gutiérrez Vázquez3 y Carlos Mallén Rivera4
1Universidad Autónoma Agraria Antonio Narro. Centro Regional Académico Chiapas. Correo-e:bn_gutierrez@hotmail.com
2 Campo Experimental Valles Centrales. CIR - Pacifico Sur. INIFAP.
3 Instituto Tecnológico Superior de San Miguel El Grande.
4Centro Nacional de Investigación Disciplinaria en Conservación y Mejoramiento de Ecosistemas Forestales. INIFAP.
Fecha de recepción: 12 de noviembre de 2011;
Fecha de aceptación: 3 de julio de 2012.
RESUMEN
De acuerdo con la Norma Oficial Mexicana NOM-059 SEMARNAT-2010, Pinus oocarpa no es una especie en peligro de extinción, pero sus poblaciones adaptadas localmente son de considerable valor como recursos genéticos, y en la actualidad son objeto de destrucción severa debido al cambio de uso de suelo. Por ello se llevó a cabo un estudio en tres de sus áreas naturales con el objetivo de determinar la variación fenotípica entre ellas. Se evaluaron 12 variables morfológicas: cinco dasométricas, tres de conos y cuatro de acículas; y se practicaron varios tipos de análisis: de varianza multivariado, de componentes principales y de agrupamiento. Se obtuvieron diferencias significativas (P<0.01) entre las poblaciones de Motozintla, General Leyva y Corazón del Valle para las variables dasométricas, conos y acículas. La de Corazón del Valle es fenotípicamente diferente a las de Motozintla y General Leyva. La variación en las características dasométricas está ligada a factores ambientales. Las correspondientes a los conos y las acículas muestran una fuerte tendencia a que las de Motozintla y General Leyva pertenezcan a una misma unidad genética y estén estrechamente ligadas a las poblaciones de América Central. Es posible que la variación inherente observada en las correspondientes a las de Corazón del Valle se restrinja a una adaptación local hacia un clima cálido subhúmedo, debido a la gran plasticidad ecológica de la especie.
Palabras clave: Análisis multivariado, conservación de recursos genéticos, genética, Pinus oocarpa Schiede ex Schltdl., variación ambiental, variación fenotípica.
ABSTRACT
According to the NOM-059 Mexican Official Standard, Pinus oocarpa is not an endangered species; however, its locally adapted populations are considerably valuable as genetic resources and are currently object of severe destruction due to the change of land use. For this reason a study was carried out in three of its natural populations with the purpose of determining the phenotypic variations between them. Twelve morphological variables –i.e. five mensuration, three cone and four needle variables– were assessed, and multivariate variance, principal component and grouping analyses were practiced. Significant differences (P<0.01) were found among the populations of Motozintla, General Leyva and Corazón del Valle, according to the mensuration, needle and cone variables. Variation in the mensuration characteristics is linked to environmental factors. The variations in the needles and cones show that it is very likely that the populations of Motozintla and of General Leyva belong to the same genetic unit and are closely linked to the populations of Central America. The inherent variation observed in the cones and needles of Corazón del Valle may be due strictly to a local adaptation to a subhumid warm climate made possible by the great ecological plasticity of the species.
Key words: Multivariate analysis, preservation of genetic resources, genetics, Pinus oocarpa Schiede ex Schltdl., environmental variation, phenotypic variation.
INTRODUCCIÓN
Se considera que la subdivisión Oocarpae del género Pinus, definida por Little y Critchfield (1969) evolucionó en México. Un estudio molecular realizado por Krupkin et al. (1996) muestra que la subdivisión es monofilética, pero hay puntos de vista alternos (Gernandt et al., 2005). El ancestro de Oocarpae, que incluye a los de Pinus oocarpa Schiede ex Schltdl., probablemente existieron en el Mioceno (Axelrod y Cota, 1993). Muchos de estos pinos, son importantes en las plantaciones forestales en el trópico y en el subtrópico (Barnes y Styles, 1983).
Pinus oocarpa Schiede ex Schltdl. var. oocarpa es de madera dura de la subdivisión Oocarpae, y el más común en Mesoamérica. Se distribuye desde el sur de Sonora en México, al norte de Nicaragua, en una longitud de 3 000 km. Es un árbol pequeño (10-13 m), de apariencia rústica en sitios secos de la Sierra Madre Occidental del noreste de México, pero es más alto (20–35 m) y desarrolla ejemplares mejor conformados en áreas con mayor precipitación, desde el sur de México hasta Nicaragua (Robbins, 1983).
Debido a la distribución geográfica tan extensa, Pinus oocarpa es, para algunos autores, la especie ancestral de la subdivisión Oocarpae; un análisis mediante marcadores RAPD de las relaciones evolutivas entre 10 taxas de esta lo confirmaron (Dvorak et al., 2000b). La población ubicada al noroeste de México en la Sierra referida parece representar históricamente una colonización reciente, hipótesis consistente con la información geológica sobre el vulcanismo y formación de las montañas en el área (Dvorak et al., 2000b).
A nivel nacional, P. oocarpa tiene una amplia distribución en las Sierras Madre Oriental y Occidental y al sur del Eje Neovolcánico. Está presente en todo el estado de Chiapas con mayor dominancia al este de Ocosingo, entre los 300 y 700 m de altitud en la región de Arriaga y Ocosingo, lo que sugiere una enorme plasticidad ecológica (Challenger, 1998). Desde el punto de vista económico es muy importante en México como madera aserrada y material combustible (Zamora, 1981).
En Chiapas es uno de los taxa de pino más utilizado en plantaciones forestales (Gutiérrez et al., 2010), en zonas tropicales y subtropicales de Colombia, Brasil y en varios países de África se han establecido extensas áreas de P. oocarpa con este mismo objetivo, en donde la calidad de su madera en ocasiones supera a la de otros pinos de rápido crecimiento, como P. patula Schltdl. et Cham., P. maximinoi Moore, y P. tecunumanii Eguiluz et Perry (Dvorak et al., 2000a). A pesar de su importancia en otros países, en México recibe una atención marginal, por lo que es necesario realizar estudios que permitan mejorar su uso en plantaciones comerciales, de restauración ecológica y en programas de conservación biológica (Viveros et al., 2005).
Aunque Pinus oocarpa no está catalogada en peligro de extinción en la Norma Oficial Mexicana NOM-059-2010 (Semarnat, 2010) en la actualidad las poblaciones localmente adaptadas y de considerable valor científico corren el riesgo de ser destruidas por el cambio de uso de suelo, al transformar los bosques en pastizales y a cultivos agrícolas. Antes de que sea demasiado tarde es indispensable establecer un programa de conservación de esta especie, el cual requerirá de la protección de un número de rodales representativos de los recursos genéticos. Para diseñarlo y decidir el lugar, el número y tamaño de las unidades de manejo forestal (Millar y Libby, 1991), se deben conocer los patrones de variación genética y fenotípica entre y dentro de las poblaciones.
Las variantes ambientales de una región a otra hacen posible suponer que existe una diferenciación geográfica con respecto a los caracteres morfológicos y anatómicos de los árboles que las conforman (Reyes et al., 2006). Los patrones de variación natural en especies y sus complejos han sido analizados por diversos autores mediante técnicas de análisis multivariado; de esta forma podría establecerse el patrón de asociación o variación entre las poblaciones de Pinus oocarpa.
A continuación se describe un estudio orientado a determinar la variación fenotípica entre tres poblaciones naturales de Pinus oocarpa, en el estado de Chiapas, México; los resultados únicamente son indicativos de tendencias de datos que podrían obtenerse usando tamaños de muestra más grandes. Sin embargo, al considerar la falta de información sobre la variación genética y fenotípica de poblaciones de pino en México, esta ayudará a plantear prioridades de conservación, no solo basadas en la diversidad de marcadores moleculares, ya que se puede inducir a la pérdida de poblaciones adaptadas localmente.
MATERIALES Y MÉTODOS
Se muestrearon tres poblaciones de Pinus oocarpa ubicadas en los ejidos de Motozintla, municipio Motozintla de Mendoza; General Leyva Velázquez, municipio Las Margaritas y Corazón del Valle, municipio Cintalapa de Figueroa en el estado de Chiapas (Figura 1), donde la especie se distribuye de manera natural (Cuadro 1). A partir de recorridos de campo se seleccionaron y delimitaron las áreas de estudio de manera directa, y en consideración de la madurez de los individuos que integraban cada una de las poblaciones. Se aplicó un muestreo simple aleatorio con sitios circulares de 500 m2, en los que se registraron las características dasométricas de los árboles.


En cada localidad se obtuvieron muestras vegetativas y reproductivas de 10 a 12 árboles, la recolecta se hizo de manera sistemática con base en los gradientes altitudinales dentro del sitio. Se consideró que las muestras fueran representativas de las condiciones de altitud, exposición y pendiente del terreno.
También se llevó a cabo la recolecta de conos y acículas mediante un recorrido dentro del área que delimita cada una de las poblaciones para la selección de los individuos con las mejores características de porte; dominantes, bien conformados, libres de plagas y enfermedades, y con presencia de conos maduros al momento de su recolección (Rodríguez y Capó, 2005). La edad se obtuvo con la extracción de un cilindro de madera a 1.30 m de altura del fuste, con un taladro de Pressler. Los árboles se escogieron con un distanciamiento entre ellos de 50 a 100 m (Munive et al., 2008), para disminuir el grado de parentesco
Trabajo de laboratorio
Las mediciones de los conos se hicieron sobre 10 ejemplares por árbol, mismos que se tomaron en función de su madurez y la presencia de pedúnculo; no se incluyeron los que presentaban señales de abertura. Se trabajó con un vernier digital con aproximación a milímetros. La longitud se registró desde la inserción del pedúnculo hasta el ápice. El diámetro se midió a partir de la parte más gruesa del mismo (Munive et al., 2008). La longitud del pedúnculo se consideró desde la inserción de la rama hasta la base del cono. Para evaluar a las acículas se separaron ocho fascículos de cada árbol y se utilizó una regla graduada en milímetros.
Variables evaluadas
Se analizaron 12 características dasométricas y anatómicas de conos y acículas, como variables importantes desde el punto de vista taxonómico. Las del primer tipo fueron la altura total, la altura de fuste limpio, el diámetro normal, el diámetro de la copa y el grosor de la corteza. Las del cono fueron la longitud de cono, su diámetro y la longitud de pedúnculo. En las acículas se contabilizó su número por fascículo y se determinó el largo y ancho de cada una y la longitud de la vaina.
Análisis estadístico
Antes del análisis multivariado se estimó el coeficiente de correlación entre pares de variables, mediante los valores promedio por población, con el objetivo de eliminar aquellas con una correlación > 0.85; de acuerdo con los resultados, no se destacó ninguna variable, por lo que el análisis final incluyó los 12 caracteres.
Con el procedimiento MANOVA de SAS versión 9.1 (SAS Institute, 1999), se realizó un análisis de varianza multivariado para determinar la presencia de una variación significativa entre poblaciones, para lo cual se consideraron por separado las características dasométricas, de las de los conos y las acículas evaluadas. El modelo estadístico utilizado fue:
Yij = μ + Pi + εi(j) (1)
Donde:
Yij = Valor multivariado observado en el j-ésimo árbol en la i-esima población
μ = Valor multivariado promedio
Pi = Efecto aleatorio de la i-ésima población [E (Pi)=0; Var (Pi)= σ2P]
εi(j) = Error aleatorio del j-ésimo árbol dentro de la i-ésima población [E (εi(j))=0 Var (εi(j))= σ2ε].
Además, se aplicó un análisis de componentes principales (CP) para identificar las variables más importantes en la expresión de la diferenciación de las poblaciones y reducir la dimensión de la variabilidad en el menor número posible de CP. En este se utilizó la matriz de correlaciones con el procedimiento PRINCOMP de SAS versión 9.1 (SAS Institute, 1999). Con los valores propios obtenidos de los dos componentes principales, se representaron gráficamente las poblaciones para identificar el patrón de agrupamiento entre ellas y su ubicación con respecto a las muestras de Pinus oocarpa incluidas en el estudio. El análisis de varianza multivariado y el de componentes principales se llevó a cabo por separado de acuerdo a las variables dasométricas, conos y acículas, porque se trabajó con diferente número de árboles y de observaciones por individuo.
Con los datos promedio por población de las características originales estandarizadas se determinaron las distancias entre pares de poblaciones y se efectuó un análisis de agrupamiento de las mismas, a partir del procedimiento CLUSTER del SAS versión 9.1 (SAS Institute, 1999) con el método del Ward Pseudo. A partir de este análisis se generó el dendrograma correspondiente, al incorporar la opción TREE del procedimiento anterior. También se practicó un análisis de correlación de Pearson entre las dos doce variables evaluadas y la precipitación media anual, la temperatura media anual y la altitud.
RESULTADOS
Análisis de varianza multivariado
Se obtuvo una variación significativa (p≤0.001) entre poblaciones al incluir las 12 variables en el análisis estadístico (Cuadro 2). En el análisis de las poblaciones, con base en las variables dasométricas, se observó una similitud entre Motozintla y General Leyva, ya que no presentaron diferencias significativas. Sin embargo, la de Corazón del Valle fue estadísticamente diferente de las otras dos, pues sus valores fueron más bajos en casi todas variables dasométricas evaluadas, en contraste con las de Motozintla que registraron los valores más altos (Cuadro 3).


El patrón de variación en las variables del cono fue diferente, ya que las poblaciones de Corazón del Valle y General Leyva fueron estadísticamente iguales pero diferentes a Motozintla, en la que también los registros fueron superiores en las características de los conos. Con respecto a las acículas se determinó el mismo patrón que en las variables dasométricas en las que se observó una similitud entre Motozintla y General Leyva, excepto en el ancho de la acícula (Cuadro 3).
Análisis de componentes principales
Los primeros dos componentes se consideraron suficientes, ya que en ellos se concentra poco más de 80% de la variación total de las características incluidas en el análisis (Cuadro 4); además de, su mayor contribución relativa individual. De acuerdo a las características dasométricas, las variables con mayor contribución relativa en el primer CP fueron la altura total y el diámetro normal; en el segundo, la altura del fuste limpio. En las variables de conos, la longitud y el diámetro hicieron el mayor aporte al primer CP y la longitud del pedúnculo en el segundo. De las correspondientes a las acículas, la longitud es la que más contribuye en el primer CP y la longitud de vaina en el segundo CP (Cuadro 5).

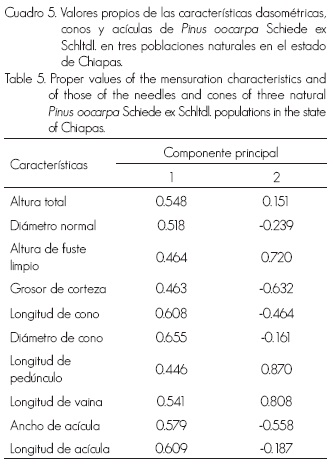
La dispersión de las poblaciones, con base en las variables dasométricas en los dos primeros componentes principales, permite distinguir la formación de dos grupos (Figura 2). El primero de ellos está integrado por las poblaciones Motozintla y General Leyva, mientras que el otro solo por la población de Corazón del Valle. Al observar los dos primeros CP, resulta que Motozintla y General Leyva están muy cercanas entre sí, pero separadas de Corazón del Valle. De acuerdo con los eigenvectores, la población de Corazón del Valle se diferencia de las otras dos poblaciones por tener menor altura total y diámetro normal (Figura 2).

Al igual que las variables dasométricas, al realizar el análisis de las variables de conos, la dispersión de las poblaciones con base en los dos primeros componentes principales mostró dos grupos diferentes (Figura 3). El primero está formado por las poblaciones General Leyva y Corazón del Valle, y uno más por la de Motozintla. Se observa que Corazón del Valle y General Leyva, a partir de los dos primeros CP, están muy cercanas y se traslapan entre sí, pero muy apartadas de la población de Motozintla. Los valores indican que la población de Motozintla se diferencia de las otras dos poblaciones por tener una mayor longitud y diámetro del cono. Asimismo se reconoce que la de General Leyva a pesar que se traslapa con Corazón del Valle, presenta una distribución más restringida, lo que obedece a valores de longitud y diámetro de conos de tamaño más reducido (Figura 3).

La dispersión de datos en las variables de acículas con relación a los componentes principales uno y dos, representa a la población de General Leyva que se comporta como intermedia; se traslapa con las de Corazón del Valle y Motozintla, las cuales están muy separadas entre sí. Aunque General Leyva se sobrepone con ambas poblaciones, se advierte ligeramente más cercana a la de Motozintla. Los valores propios muestran que en el CP1 Corazón del Valle son bajos en longitud de vaina y Motozintla, más altos, lo que permite distinguirlas mejor (Figura 4).

Análisis de agrupamiento
A través del dendrograma se corroboran los resultados del análisis de componentes principales (Figura 5). La agrupación sugiere la separación perfecta de dos grupos. Uno de ellos compuesto por las poblaciones de Motozintla y General Leyva y el otro únicamente por Corazón del Valle. Lo anterior evidencia que las 12 variables evaluadas son satisfactorias para clasificar fenotípicamente a las poblaciones estudiadas.

Análisis de correlación de Pearson
Los valores de correlación indican que solo el diámetro normal presenta correlación significativa (P<0.01), con la temperatura una correlación negativa y con la precipitación, positiva. Sin embargo, se detectó un patrón bien definido, en el cual las variables dasométricas y la temperatura, la precipitación y la altitud produjeron coeficientes de correlación altos, aunque no fueron significativos (P>0.05). Las variables de conos y acículas arrojaron coeficientes de correlación bajos, excepto para la longitud de acícula que reveló valores altos de correlación negativa con la temperatura y positiva con la precipitación, aunque no fueron significativos (Cuadro 6).

DISCUSIÓN
Las diferencias significativas y el patrón de variación en las tres poblaciones estudiadas coinciden con los resultados de Gutiérrez et al. (2010) al evaluar la densidad básica de la madera en las mismas localidades, donde Corazón del Valle fue estadísticamente mejor que Motozintla y General Leyva.
La disociación aparente de Corazón del Valle con respecto a las otros grupos analizados en las variables dasométricas está directamente influenciada por la altura total, el diámetro normal y la altura de fuste limpio. El análisis de correlación indica que estas características guardan un estrecho vínculo con el medio ambiente en el que se desarrollan. Viveros et al. (2005) obtuvieron valores de control genético elevados en diámetro y altura. A pesar de ello, se considera que la variación fenotípica en las variables dasométricas depende, en gran medida, de la variación en los factores ambientales como la altitud, la precipitación y la temperatura.
La altura también tiene una relación con la densidad de la vegetación que crece en el sitio, ya que es superior cuando la densidad es abundante, por la competencia entre los individuos (Sanz et al., 2003). Sin embargo, se observó un patrón diferente en la población de Corazón del Valle, ya que es la población con mayor densidad de individuos y registró los valores más bajos en altura.
La variación fenotípica que separa a Corazón del Valle de las otras dos poblaciones, al reunir los valores más bajos en crecimiento y desarrollo en altura y diámetro podría atribuirse a la altitud, la precipitación y la temperatura, pues esta localidad está ubicada a 800 msnm, y la diferencia con respecto a Motozintla es de alrededor de 700 m y de 1 200 m con General Leyva; además, sus valores de correlación positiva fueron destacados en relación a la altura total, el diámetro normal y la altura del fuste limpio, variables que hicieron una mejor contribución a la separación de las poblaciones. Aunado a lo anterior es importante destacar que Pinus oocarpa experimenta mejor crecimiento y desarrollo de los 1 500 a los 2 500 msnm (Perry, 1991). No obstante, Alba et al. (2003) consignaron que la probabilidad de que P. oocarpa esté presente en la altiplanicie central de Chiapas se entiende, en buena medida, por el efecto inverso de la altitud. Al igual que en el presente estudio, al ajustar modelos de regresión logística para la especie de interés, dichos autores sugieren que la temperatura y la precipitación explican su distribución de manera directa.
Al evaluar la variación isoenzimática en un gradiente altitudinal en poblaciones de Pinus oocarpa en Michoacán. Sáenz et al. (2003) determinaron que las poblaciones no se agruparon en función de la altitud; tampoco detectaron diferencias genéticas a lo largo del gradiente, y la mayor variación genética está dentro de las poblaciones. Sin embargo, las características observadas en Corazón del Valle podrían responder a una adaptación local a la altitud, que no es detectada por los marcadores moleculares debido a que estos parecen ser indicadores pobres de la variación heredable en caracteres cuantitativos (McKay y Latta, 2002). Si fuese un evento local sería muy significativo, ya que se ha reconocido que la variación genética adaptativa de especies no solo es el resultado de fuerzas bióticas selectivas o estrés climático, sino también del estrés biótico o abiótico local, como los herbívoros o los parásitos, entre otros factores (Mitton, 1997). Lo anterior resulta relevante en materia de conservación, el tener genes localmente adaptados y de importancia económica, si se trata de una población que reúne características valiosas como alta densidad de la madera (Gutiérrez et al., 2010) y que pueden ser utilizadas en programas de mejoramiento genético. Ecológicamente, la heterogeneidad del hábitat mantiene la variación genética entre las poblaciones. Plantear prioridades de conservación basadas solo en la diversidad de marcadores moleculares puede inducir a la pérdida de poblaciones adaptadas a condiciones locales (McKay et al., 2001).
En cuanto a las variables de conos se infiere que la similitud entre Corazón del Valle y General Leyva y la separación con respecto a Motozintla podría ser de tipo genético, ya que las características de dichas estructuras reciben mayor influencia de los factores ambientales (Iglesias y Tivo, 2006). La distancia que guarda la población de la segunda localidad, con respecto a las otras dos quizás se deba a que se ubica hacia el sur del estado de Chiapas; lo que significa, que tal vez Pinus oocarpa desarrolla conos grandes mientras se aleja hacia el sur, pues las poblaciones de de esta especie en la entidad son más parecidas a las de América Central que al resto de las que se desarrollan en México (Dvorak et al., 2009). Así, las de Guatemala tienen conos más grandes (Lamprecht, 1990), lo que pudiera ser una causa de separación de las otras poblaciones, ya que la de Motozintla es cercana a Guatemala.
Con respecto a la variación fenotípica en las variables de las acículas, se fortalece la posibilidad de que las poblaciones del sur, Motozintla y General Leyva, correspondan al centro de diversidad de Pinus oocarpa de América Central propuesto por Dvorak et al. (2009), y la población de Corazón del Valle estaría ligada a las poblaciones ubicadas al norte (centro de diversidad del Eje Neovolcánico de México). Es posible que las de Motozintla y General Leyva, con mayores similitudes, pertenezcan a una misma unidad genética, ya que al encontrase a una distancia más corta (130 km) que Corazón del Valle (221 km) podrían estar intercambiando información mediante flujo genético.
El análisis referente a la variación fenotípica entre las poblaciones estudiadas no permite determinar si es genética o ambiental, o en qué proporción participan cada una de ellas. Por lo tanto, al igual que Gutiérrez et al. (2010), se considera necesario realizar ensayos de procedencias-progenie en varios sitios, con un mayor número de poblaciones, para precisar el potencial de las evaluadas aquí y otras en el establecimiento de futuras plantaciones y definir si la variación es de tipo genética o ambiental.
CONCLUSIONES
Existen diferencias significativas entre las poblaciones de Pinus oocarpa de Motozintla, General Leyva y Corazón del Valle de acuerdo con sus características dasométricas, de conos y de acículas; la de la última localidad es fenotípicamente diferente a las primeras dos. La variación en las variables dasométricas puede estar más relacionada a factores ambientales. La variación identificada en las variables de los conos y las acículas muestra una fuerte tendencia a que las poblaciones de Motozintla y General Leyva pertenezcan a una misma unidad genética y estén muy ligadas a las poblaciones de América Central. Es posible que la variación que presenta la población de Corazón del Valle sea un caso de adaptación local a un clima cálido subhúmedo, debido a la gran plasticidad ecológica de la especie.
REFERENCIAS
Alba L., M. P., M. González E., N. Ramírez M. y M. A. Castillo S. 2003. Determinantes de la distribución de Pinus spp. en la altiplanicie central de Chiapas, México. Boletín de la Sociedad Botánica de México 73: 7-15. [ Links ]
Axelrod, D. I. and J. Cota. 1993. A further contribution to closed-cone pine history. Am. J. Bot. 80: 743-751. [ Links ]
Barnes, R. D. and B. T. Styles. 1983. The closed-cone pines of Mexico and Central America. Commonw For. Rev. 62:81-84. [ Links ]
Challenger, A. 1998. Utilización y conservación de los ecosistemas terrestres de México, pasado presente y futuro. Comisión Nacional para el Conocimiento y Uso de la Biodiversidad. Instituto de Biología. Universidad Nacional Autónoma de México. Agrupación Sierra Madre, S.C. México, D.F. México. 847 p. [ Links ]
Dvorak, W. S., K. M. Potter, V. D. Hipkins and G. R. Hodge. 2009. Genetic diversity and gene exchange in Pinus oocarpa, a Mesoamerican pine with resistance to the pitch canker fungus (Fusarium circinatum). Int. J. Plant Sci. 170 (5):609-626. [ Links ]
Dvorak, W. S., E. A. Gutiérrez, L. F. Osorio, G. R. Hodge and J. T. Brawner. 2000a. Pinus oocarpa. In: CAMCORE Cooperative (ed.). Conservation and testing of tropical and subtropical forest tree species by the CAMCORE Cooperative College of Natural Resources, North Carolina State University. Raleigh, NC. USA. pp. 129-147. [ Links ]
Dvorak, W. S., A. P. Jordan, G. R. Hodge and J. L. Romero. 2000b. Assessing evolutionary relationships of pines in the Oocarpae and Australes subsections using RAPD markers. New For. 20:163-192. [ Links ]
Gernandt, D., G. Geada L., S. Ortiz G. and A. Liston. 2005. Phylogeny and classification of Pinus. Taxon 54:29-42. [ Links ]
Gutiérrez V., B., M. Gómez C., S. Valencia M., E. H. Cornejo O., J. A. Ruiz P. y M. H. Gutiérrez V. 2010. Variación de la densidad de la madera en poblaciones naturales Pinus oocarpa Schiede ex Schltdl. en el estado de Chiapas, México. Rev. Fitotec. Mex. 33 (4):75-78. [ Links ]
Iglesias A., L. G. y F. Tivo 2006. Caracterización morfométrica de la población de Pinus hartwegii Lindl. del Cofre de Perote Veracruz. Ra Ximhai 2:449-468. [ Links ]
Krupkin, A. B., A. Liston and S. H. Strauss. 1996. Phylogenetic analysis of the hard pines (Pinus subgenus Pinus, Pinaceae) from chloroplast DNA restriction site analysis. Am. J. Bot. 83:489-498. [ Links ]
Lamprecht, H. 1990. Silvicultura en los trópicos. Deustche Gesellschaft fúr Technische Zusammenarbeit. Eschborn Hesse, Deustcheland.. pp. 36-50. [ Links ]
Little, E. L., Jr. and W. B. Critchfield. 1969. Subdivisions of the genus Pinus (Pines). USDA Forest Service Miscellaneous Publication 1144. Washington, DC USA.18 p. [ Links ]
McKay, J. K., J. G. Bishop, J. Z. Lin, J. H. Richards, A. Sala and O. T. Mitchell. 2001. Local adaptation across a climatic gradient despite small effective population size in the rare sapphire rockcress. Proc. Royal Society- Biological Sciences. 268 (1477):1715-1721. [ Links ]
McKay, J. K. and R. G. Latta. 2002. Adaptive population divergence: markers, Qtl and traits. Trends in Ecology and Evolution 17:285-291. [ Links ]
Millar, C. I. and W. J. Libby. 1991. Strategies for conserving clinal, ecotypic and disjunct population diversity in widespread species. In: Falk, D. A. and K. E. Holsinger. (eds.). Genetics and conservation of rare plants. Oxford University Press, New York, NY. USA. pp. 149-170. [ Links ]
Mitton, J. B. 1997. Selection in natural populations. Oxford University Press. New York, NY. USA. 240 p. [ Links ]
Munive M., E., O. Vázquez C., E. M. Zamora C., E. Fernández P. y E. García G. 2008. Variación de conos y semillas de Pinus ayacahuite var. veitchii Shaw de dos procedencias del estado de Tlaxcala. Foresta Veracruzana 10(1):39-46. [ Links ]
Perry, J. P. Jr. 1991. The pines of Mexico and Central America. Timber, Portland, OR. USA. 231 p. [ Links ]
Reyes H., V. J., J. J. Vargas H., J. López U. y H. Vaquera H. 2006. Similitud fenotípica de poblaciones mexicanas de Pseudotsuga Carr. Agrociencia 40:545-556. [ Links ]
Robbins, A. M. J. 1983. Pinus oocarpa Schiede. Seed leaflet No. 3. DANIDA Forest Seed Centre, Humlebaek, Denmark. 17 p. [ Links ]
Rodríguez L., R. y M. Capó A. 2005. Morfología de acículas y conos en poblaciones naturales de Pinus arizonica Engelm. Ra Ximhai 1:131-152. [ Links ]
Sanz de la F., M. del Río G. y R. Sierra de G. 2003. Efectos de la densidad de regeneración tras incendios sobre la características dasométricas del regenerado de Pinus pinaster Aiton, en la región de procedencia de Oña. Madrid, España. Ciencia Forestal 15:177-182. [ Links ]
Statistic Analysis System Institute (SAS). 1999. SAS/STAT Guide for Personal Computers. 9.1 Version. Cary, NC. USA. 518 p. [ Links ]
Secretaría del Medio Ambiente y Recursos Naturales (Semarnat). 2010. Norma Oficial Mexicana NOM–059–SEMARNAT–2010, Protección ambiental – Especies nativas de México de Flora y Fauna Silvestres – Categorías de Riesgo y especificaciones para su inclusión, exclusión o cambio – Lista de Especies en Riesgo. Diario Oficial de la Federación 2a. Sección, 30 de diciembre del 2010. http://dof.gob.mx/nota_detalle_popup.php?codigc. (10 de septiembre de 2011). [ Links ]
Viveros V., H., C. Sáenz R. y R. R. Guzmán R. 2005. Control genético de las características de crecimiento en vivero de plántulas de Pinus oocarpa. Rev. Fitotec. Mex. 28: 333-338. [ Links ]
Zamora S., C.1981. Algunos aspectos sobre Pinus oocarpa Schiede en el estado de Chiapas. Ciencia Forestal Vol. 6 (32): 25-53. [ Links ]

