










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista mexicana de ciencias forestales
Print version ISSN 2007-1132
Rev. mex. de cienc. forestales vol.4 n.17 México May./Jun. 2013
Artículo
Efecto de la ganadería en la composición y diversidad arbórea y arbustiva del matorral espinoso tamaulipeco
Efect of livestock in the composition and diversitiy of trees and shrubs in the tamaulipan thornscrub
Carlos Alberto Mora Donjuán1, Javier Jiménez Pérez1, Eduardo Alanís Rodríguez1, Ernesto Alonso Rubio Camacho2, José Israel Yerena Yamallel1 y Marco Aurelio González Tagle1
1Facultad de Ciencias Forestales, Universidad Autónoma de Nuevo León. Correo–e: biologomora@gmail.com
2Campo Experimental Centro Altos de Jalisco, CIR-Pacífico Centro, INIFAP
Fecha de recepción: 1 de febrero de 2013.
Fecha de aceptación: 1 de abril de 2013.
RESUMEN
El pastoreo se realiza en 26% de la superficie terrestre; mientras que la producción de forrajes en 33% de las tierras de cultivo agrícola. En México, la ganadería es la actividad productiva más común en el medio rural: abarca 110 millones de hectáreas; es decir, 56% del territorio nacional. El matorral es el ecosistema más abundante e históricamente del que se obtiene mayor número de productos en las zonas áridas y semiáridas del país. La presente investigación evaluó el impacto de la ganadería en la composición y diversidad en diferentes áreas del Matorral Espinoso Tamaulipeco. Se seleccionaron tres: Referencia, Regeneración y Ganadería; y se establecieron cuatro sitios de muestreo de 1 600 m2 en cada una. Se determinaron variables estructurales de abundancia, dominancia, frecuencia e Índice de Valor de Importancia; además se estimaron los índices de riqueza de especies, diversidad alfa y beta. Se registraron 22 especies, distribuidas en 20 géneros y 14 familias. El peso ecológico estuvo representado en el área de Referencia por Diospyros texana, en la de Regeneración por Acacia farnesiana y en la de Ganadería por Prosopis laevigata. El estudio revela que la ganadería disminuye significativamente la abundancia, dominancia y diversidad alfa de las comunidades arbóreas y arbustivas, y que las resultantes muestran una baja similitud de especies.
Palabras clave: Comunidad de referencia, diversidad alfa y beta, indicadores ecológicos, Matorral Espinoso Tamaulipeco, regeneración, restauración ecológica.
ABSTRACT
Grazing is carried out in 26% of the land cover, while forage production in 33% of crop lands. In Mexico, livestock is the most common productive activity in the rural scope: it gathers 110 million hectares, that is, 56% of the national territory. The scrub is the most abundant ecosystem and, historically, from which are obtained the greatest number of products in the arid and semiarid zones of the country. This research assessed the impact of livestock in the composition and diversity in different areas of the Tamaulipan Scorncrub. Three were selected: Referencia, Regeneración and Ganadería and four sampling plots of1 600 m2 each were established. Structural variables of abundance, dominance, frequency and the Importance Value Index were determined, in addition to the indexes of species richness, alfa and beta diversity. Twenty-two species were recorded, distributed into 20 genus and 14 families. The ecological weight was represented in the Referencia area by Diospyros texana, in Regeneración by Acacia farnesiana and in Ganadería, by Prosopis laevigata. The study reveals that livestock lowers significantly the abundance, dominance and alfa diversity of the tree and scrub communities and that those resulting, show low similitude among species.
Key words: Reference community, alfa and beta diversity, ecological indicators, Tamaulipan Thornscrub, regeneration, ecological restoration.
INTRODUCCIÓN
El ganado representa 40% del valor mundial de la producción agrícola y es la base de los medios de subsistencia y de la seguridad alimentaria de casi mil millones de personas (FAO, 2009). El pastoreo se realiza en 26% de la superficie terrestre, que no está cubierta por hielo, mientras que la producción de forrajes emplea 33% de las tierras de cultivo agrícola (Steinfeld et al., 2006); ambas constituyen prácticamente 80% de todas las actividades que se llevan a cabo en tierras agrícolas, y de ellas 3 400 millones de hectáreas están sujetas al pastoreo y 500 millones a cultivos para la alimentación animal (Steinfeld et al., 2006).
En México, la ganadería es la actividad productiva más común en el medio rural, ya que se practica, sin excepción, en todas las regiones ecológicas y aun en condiciones climáticas adversas. Ocupa 110 millones de hectáreas, lo que equivale a casi 56% del territorio nacional (Sagarpa, 2006). En particular, el estado de Nuevo León cuenta con una superficie ganadera de 5.5 millones de hectáreas (86% del total estatal), de las cuales 90% son de agostadero y 10% praderas (Sagarpa, 2009).
Hay diferentes visiones sobre los problemas que generan las actividades ganaderas y estas varían dependiendo de cada región; sin embargo, existen consensos significativos sobre los impactos más preocupantes como la deforestación de los bosques, la erosión y compactación de los suelos, las emisiones de gases nocivos para la atmósfera, la contaminación de aguas, la eutroficación de zonas costeras, los cambios en la cobertura vegetal, la disminución de la biodiversidad y el uso de recursos no renovables: la energía fósil y los fertilizantes (Sadeghian et al., 1999; Abril, 2011; Villanueva et al., 2011).
El matorral es el ecosistema más abundante e históricamente más utilizado en las zonas áridas y semiáridas de México (García y Jurado, 2008). A través del tiempo ha sido afectado por actividades antrópicas como la extracción de especies vegetales para diferentes usos (Estrada et al., 2004; Rzedowski, 2006; García y Jurado, 2008); y a una continua deforestación para el establecimiento de zonas agrícolas, industriales y urbanas (Alanís et al., 2008a; Arriaga, 2009). Por otra parte, debido a que el uso ganadero es la práctica más común, el efecto sobresaliente del pastoreo es la sustitución paulatina de las plantas nativas (Rzedowski, 2006; García y Jurado, 2008). En el periodo 1993-2002, el matorral perdió 953 mil ha por cambio de uso de suelo, por lo que es el segundo ecosistema más afectado en México, después de las selvas (Semarnat, 2006).
Una de las principales causas del cambio de uso de suelo es el establecimiento de pastos exóticos para la ganadería intensiva. Las áreas destinadas a ese fin son empleadas durante un tiempo y abandonadas, cuando bajan su productividad. Posteriormente, pueden regenerarse, aunque se desconoce cómo serán las comunidades vegetales resultantes. Existen algunas investigaciones sobre la estructura de la vegetación leñosa, como las de Alanís et al. (2008a), Jiménez et al. (2009), González et al. (2010), Jiménez et al. (2012), Pequeño et al. (2013) quienes evaluaron o caracterizaron diversas áreas del MET con diferente historial de uso, pero sin analizar áreas donde actualmente se desarrolla la actividad pecuaria. Molina et al. (2013) documentaron una modificación de la vegetación natural como resultado del pastoreo de ganado bovino, mas no la compararon con otra área disturbada o con una de referencia.
A consecuencia del intenso cambio de uso de suelo y de los escasos estudios realizados en ecosistemas como el MET es necesario desarrollar trabajos en las comunidades resultantes ya que proveen bases documentales para determinar, de manera objetiva, cuál es la relación gestión–impacto y cuál es la dirección del desarrollo sucesional de las distintas asociaciones vegetales (García y Jurado, 2008).
La presente investigación tiene como objetivo evaluar en tres áreas el efecto de la ganadería en la composición y diversidad arbórea y arbustiva en el Matorral Espinoso Tamaulipeco (MET) del noreste de México.
MATERIALES Y MÉTODOS
Área de estudio
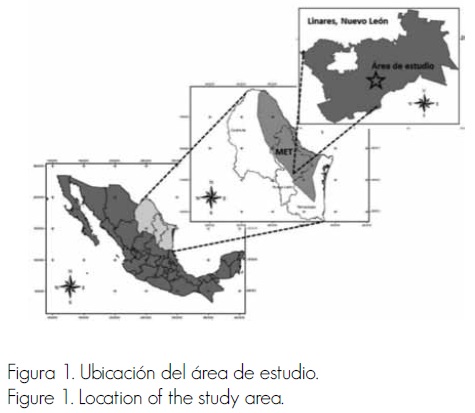
La investigación se desarrolló en Linares, Nuevo León (Figura 1), entre las coordenadas 25°09’ y 24°33’ de latitud norte y 99°54’ y 99°07’ de longitud oeste, a 4 km del ejido Las Barretas, en el rancho El Consuelo, ubicado a 24° 81’ latitud norte y 99° 41’ longitud oeste. Las especies que destacan por su abundancia y cobertura son Acacia amentacea DC., Acacia farnesiana (L.) Willd., Havardia pallens (Benth.) Britton & Rose, Cordia boissieri A. DC., Karwinskia humboldtiana (Schult.) Zucc. y Prosopis glandulosa Torr. (Espinoza y Návar, 2005; Alanís et al., 2008a).
Análisis de la vegetación
En la primavera de 2012 se seleccionaron tres áreas para evaluar la vegetación en un Matorral Espinoso Tamaulipeco (MET): 1) Referencia, donde se tiene la certeza de que no se ha realizado cambio de uso de suelo en los últimos 70 años y no hay algún tipo de intervención antropogénica en el periodo 1983–2012, ya que existe un cerco perimetral desde 1983; 2) Regeneración, esta área se desmontó con maquinaria agrícola (1989) para dedicarla a cultivos durante cuatro años (1990-1994), después se abandonó y desde entonces (1994-2012) se usa como área ganadera o de agostadero; y 3) Ganadería, zona de agostadero activo, espacio que en ningún momento fue desmontado, solo se introdujo ganado.
En cada área se establecieron sitios de muestreo cuadrados de 1 600 m2; forma que se utilizó debido a la facilidad para su delimitación y medición de la vegetación densa (Pequeño et al., 2013), respecto a la forma circular tradicional (Alanís et al., 2008b); su distribución fue al azar, y para la obtención de su número mínimo se elaboró una curva especie–área para cada comunidad, a partir del criterio de Müeller y Ellenberg (1974). Se fijaron cuatro sitios de muestreo en cada una, en los cuales se efectuó un censo de todos los individuos leñosos ≥5cm de diámetro (d0.10), y se midieron los diámetros de copa (dcopa). Se realizaron colectas botánicas de todas las especies para su posterior identificación por personal de la Facultad de Ciencias Forestal, UANL.
Análisis de la información
La riqueza específica (S) es el número total de especies presentes en determinada área o sitio. Para estimarla se utilizó el índice de Margalef (DMg) y para la diversidad el índice de Shannon–Weiner (H´) (1948), mediante las ecuaciones:

Donde:
S = Número de especies presentes
N= Número total de individuos
ni= = Número de individuos de la especie i
Para cada especie se determinó su abundancia, de acuerdo al número de individuos; su dominancia, en función de la cobertura de copa; y su frecuencia, con base en su existencia en los sitios de muestreo. Los resultados se emplearon para calcular un valor ponderado a nivel de taxón, denominado Índice de Valor de Importancia (IVI), el cual se expresa en términos porcentuales en una escala de 0 a 100 (Müeller y Ellenberg, 1974; Magurran, 2004). Para la estimación de la abundancia relativa se usó la ecuación:
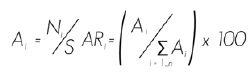
Donde:
Ai= Abundancia absoluta
ARi= Abundancia relativa de la especie i respecto a la abundancia total
Ni=Número de individuos de la especie i
S= Superficie de muestreo (ha)
La dominancia relativa se evaluó con la ecuación:

Donde:
Di= Dominancia absoluta
DRi = Dominancia relativa de la especie i respecto a la dominancia total
Abi= Área de copa de la especie i
S= Superficie (ha)
La frecuencia relativa se obtuvo con la expresión:

Donde:
Fi= Frecuencia absoluta
FRi= Frecuencia relativa de la especie i respecto a la frecuencia total
Pi= Número de sitios en los que está presente la especie i
NS = Número total de sitios de muestreo
El Índice de Valor de Importancia (IVI) adquiere valores porcentuales de 0 a 100% y se define como (Whittaker, 1972; Moreno, 2001):
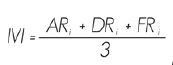
Donde:
ARi = Abundancia relativa de la especie i respecto a la abundancia total
DRi = Dominancia relativa de la especie i respecto a la dominancia total
FRi = Frecuencia relativa de la especie i respecto a la frecuencia total
En las tres áreas de estudio se obtuvo el área basal y área de copa de las especies con la fórmula:

Donde:
A= Área
d= Diámetro
A fin de estimar la similitud (o disimilitud) entre los sitios de estudio se utilizó el índice de Bray–Curtis (Beals, 1984), uno de los más usados en la ecología cuantitativa actual y sus expresiones de similitud y disimilitud son:

Donde:
Min = Mínimo obtenido de las dos variables en las mismas muestras
Xij = Abundancia de la especie i en la muestra j
Xik = Abundancia de la muestra i en la muestra k
En el análisis de los datos se empleó el software de licencia libre PAST©. Este índice concede, todavía, un importante peso a los valores altos, ya que en su expresión, el numerador incluye la diferencia entre los atributos. No obstante, dado que la sumatoria de las diferencias no se eleva al cuadrado y posteriormente se divide entre la sumatoria de las sumas individuales, el índice de Bray-Curtis es una opción menos sesgada que la distancia euclidiana (Herrera, 2000).
Con objeto de evaluar si existía una diferencia significativa de las variables de riqueza específica, índice de Margalef, índice de Shannon, densidad, área de copa y área basal entre las áreas evaluadas, se procedió a calcular los valores promedio de los sitios de muestreo.
Para definir si los datos presentaban una distribución normal se utilizó la prueba de normalidad de Shapiro Wilks en grupos pequeños; los resultados indicaron que si tenían una distribución normal. A fin de determinar si las diferencias entre sitios fueron estadísticamente significativas se usó el análisis de varianza. Posteriormente, en aquellos que resultaron con varianzas distintas se aplicó el estadístico de T con ajuste de Bonferroni para múltiples comparaciones y así contrastar dichas diferencias entre sitios. La elección de Bonferroni, en lugar de Tukey, responde al tamaño de las muestras.
RESULTADOS Y DISCUSIÓN
Se registraron 22 especies arbóreas y arbustivas, distribuidas en 20 géneros y 14 familias (Cuadro 1). La familia que tuvo mayor riqueza específica fue Fabaceae con ocho taxa. Estos resultados concuerdan con González et al. (2010) y Molina et al. (2013), quienes señalan en sus estudios a Fabaceae como la mejor representada. Es importante mencionar que solo Prosopis laevigata se observó en las tres áreas de estudio. García y Jurado (2008), al analizar un matorral en condiciones prístinas consignaron una cantidad menor de especies; un comportamiento similar documentaron Jiménez et al. (2012) en un MET con historial agrícola, mientras Canizales et al. (2009) citan una riqueza de especies mayor a la presentada en un matorral submontano. Sin embargo, estas investigaciones tenían otros objetivos y analizaron a todos los individuos mayores a un centímetro de diámetro; en contraste, para este estudio se consideraron todos los individuos mayores a cinco centímetros de diámetro.
Indicadores ecológicos
Abundancia. Esta variable mostró diferencias estadísticamente significativas entre áreas (f=36.2, g.l.=2, p<0.001). El área de Referencia las tuvo con las de Ganadería y Regeneración con p<0.000 en ambos casos; por el contrario, estas dos no difieren estadísticamente (Figura 2).

La abundancia (densidad) está representada en individuos por unidad de área (ind ha-1). La que exhibió los valores más altos fue la de Referencia, con 1763±426 ind ha-1; en segundo término, el área de Regeneración, con 491±89 ind ha-1, por último, con el menor registro del estudio, la de Ganadería, con 398 ± 54ind ha-1. La investigación de Jiménez et al. (2009, 2012), en la que analizaron áreas con diferente historial, difiere de los resultados obtenidos en este estudio. Pequeño et al. (2013) lograron una densidad superior a la determinada en las áreas evaluadas del presente análisis.
La Figura 3 evidencia la densidad de individuos por hectárea, de acuerdo con las clases diamétricas. En el área de Referencia se nota una línea de tendencia exponencial negativa conforme aumentan los diámetros: la clase 4-8 cm fue la que presentó más individuos (1259 ind ha-1). Esto indica que en las clases diamétricas menores se concentraron los individuos, lo cual demuestra que el área está en un estado de regeneración activo. Esta información coincide con la asentada en un trabajo realizado en el MET por Jiménez et al. (2012). Asimismo, el área de Ganadería concentra el mayor número de ejemplares en la clase diamétrica de 4-8 cm (180 ind ha-1), con una tendencia exponencial negativa. No obstante, a diferencia del área anterior, también cuenta con un porcentaje considerable en las clases diamétricas 14-17 y 17-20 (97 y 75 ind ha-1, respectivamente). Una de las posibles explicaciones es que en muchas áreas ganaderas del noreste de México desmontan los agostaderos casi por completo y solo dejan algunos ejemplares grandes para sombra y descanso para el ganado. Igualmente, Pequeño et al. (2013) lograron el agrupamiento más grande de individuos en la primera clase diamétrica, en un área pospecuaria. El área de Regeneración, igual que las anteriores, congrega el mayor número de individuos en la primera clase diamétrica (310 ind ha-1) y tiene una tendencia exponencial negativa similar a la de Referencia; sin embargo, esta no presenta ejemplares en las dos últimas clases diamétricas. Lo anterior responde a su historial agrícola, pues fue una zona que se desmontó, en su totalidad, para practicar agricultura y después se convirtió en un agostadero.

Dominancia (área de copa). La comparación entre áreas (Figura 4) mostró diferencias estadísticamente significativas (f=14.6, g.l.=2, p=0.001).

El contraste de medias registró que las áreas de Referencia y Regeneración, así como las de Ganadería y Regeneración difieren estadísticamente entre sí (p=0.001). El área de Referencia y la de Ganadería no exhibieron diferencias estadísticamente significativas. El área con más cobertura de copa fue la de Referencia, con 7 046.96 m2. Después le siguió en importancia la de Ganadería, con 5 436.03 m2, y por último, con menor cobertura la de Regeneración, con 3 193.32 m2 (Figura 4). Jiménez et al. (2012) estimaron una cobertura menor a la consignada en esta investigación, respecto a las tres áreas. Pequeño et al. (2013) estudiaron una comunidad regenerada pospecuaria del MET que tuvo una cobertura menor a la del área de Referencia, pero superior a las de Ganadería y Regeneración. El área de Referencia mostró diferencias significativas en relación con la de Regeneración; mientras que con la de Ganadería no tuvo diferencias; la de Regeneración sí reveló diferencias respecto a las otras dos.
La dominancia por especie en cada una de las áreas fue la siguiente: en la de Referencia, Dyospiros texana consignó 2 088.56 m2; es decir, 29.63% de la cobertura; en la de Ganadería, Prosopis laevigata fue la de más cobertura, con 4 562.65 m2, que representa 61.18% del total; y en la de Regeneración, la especie mejor representada resultó Acacia farnesiana, con 1 252.65 m2, que constituye 39.23% del total de la cobertura. García y Jurado (2008) señalaron a Helietta parvifolia (A. Gray) Benth. como la especie con mayor cobertura en un área de matorral submontano; Jiménez et al. (2009) realizaron un estudio en áreas con distinto historial (agrícola, ganadero y matarrasa) en MET, y registraron a Acacia amentacea como la especie con mayor cobertura para las áreas agrícola y matarrasa, y a Diospyros texana para la de ganadería. González et al. (2010) en un análisis de tres sitios del MET señalan a Prosopis laevigata como el taxón con más cobertura; Jiménez et al. (2012) citan a Acacia amentacea los valores más altos de cobertura en áreas con diferente historial. Pequeño et al. (2013) reconocen a Acacia farnesiana con la cobertura superior en una zona pospecuaria.
Índice de Valor de Importancia (IVI). Los taxa que sobresalieron por su peso ecológico fueron en el área de Referencia, Diospyros texana con 20.46%; en la de Ganadería, Prosopis laevigata con 52.54%, y en la de Regeneración, Acacia farnesiana con 33.93%. (Cuadro 2). Jiménez et al. (2009) evaluaron tres comunidades con historial diferente, y designaron a Diospyros texana, Bernardia myricaefolia (Scheele) S. Watson y Acacia amentacea como los taxa con mayor registro de IVI. Jiménez et al. (2012) coinciden con el área Referencia, pero no con las otras dos; por otra parte, González et al. (2010) disienten de los resultados de esta investigación, pues ellos citan a Acacia amentacea como la de más peso ecológico. Pequeño et al. (2013) concuerdan con el área de Regeneración, ya que en un área pospecuaria mencionan a Acacia farnesiana con el valor de IVI más alto.
Diversidad alfa
Riqueza de especies (S). El área que presentó una riqueza de especies superior, respecto a las otros dos, fue la de Referencia, con 21 especies; seguida por la de Ganadería, con ocho, y la de Regeneración, con cuatro (Figura 5). García y Jurado (2008) registraron una riqueza menor que la del área de Referencia, pero superior a las otras dos, mientras que Jiménez et al. (2009) la determinaron todavía mayor, ya que, entre otras razones, los autores evaluaron a todos los individuos con más de un centímetro; mientras que en el presente estudio se analizó a todos los individuos con más de cinco centímetros, lo cual puede significar la diferencia entre ambas investigaciones. Jiménez et al. (2012) señalaron una riqueza de especies menor a la del área Referencia, pero mayor a las otras dos, lo que fue confirmado por Pequeño et al. (2013) en el área pospecuaria que trabajaron.
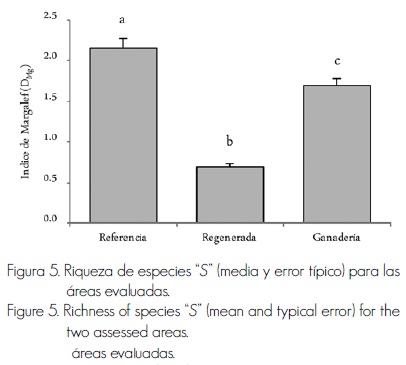
Índice de Margalef (DMg). La comparación entre áreas (Figura 6) evidencia diferencias estadísticamente significativas (f=189.5, g.l.=2, p<0.001). El área de Referencia obtuvo un índice de riqueza de especies de Margalef (DMg) de 2.16 ±0.17, el más alto en relación con las otras dos; el de Ganadería, de 1.69±0.05, y el de Regeneración, de 0.69±0.30. Margalef (1951) afirma que valores menores de dos y cercanos a cero denotan una baja riqueza del sitio; en cambio, valores superiores y más lejanos a dos sugieren una riqueza de especies alta. En sitios con disturbio agropecuario, Alanís et al. (2008a) documentan valores similares a los del área de Referencia, pero muy diferentes a las otras dos áreas. Pequeño et al. (2013) coinciden con el área de Ganadería, pues registraron índices inferiores a dos en un área pospecuaria. La riqueza de especies y la diversidad de especies son dos de los atributos ecológicos más afectadas por la actividad ganadera, probablemente a causa de la dieta del ganado, el cual es selectivo y no consume toda la vegetación existente, sino solo algunas especies palatables.

Índice de Shannon-Weiner (H´). El análisis de varianza entre áreas demostró la existencia de diferencias estadísticamente significativas (F=55.5, G.L.=2, P<0.001). En la presente investigación, la diversidad alfa (H´) se obtuvo con el índice de Shannon–Weiner, con los siguientes resultados: para el área de Referencia se registró un valor de 1.95±0.17, para la de Regeneración de 1.31±0.02, y la de Ganadería, de 1.21 ± 0.06. Las diferencias entre las áreas de Referencia/Ganadería y Referencia/[Regeneración fueron altamente significativas, con P<0.000 en ambos casos; la comparación de [Regeneración/Ganadería no difieren estadísticamente (Figura 7). Shannon–Weiner (1948) mencionan que valores cercanos a cero representan una baja diversidad, en tanto los cercanos a cinco o mayores indican una alta diversidad. Alanís et al. (2008a), González et al. (2010), Jiménez et al. (2012), Molina et al. (2013) consignaron en áreas con diferente historial del MET un índice superior al estimado en las del presente estudio; Pequeño et al. (2013) señalaron valores inferiores en un área pospecuaria del MET.

Diversidad beta
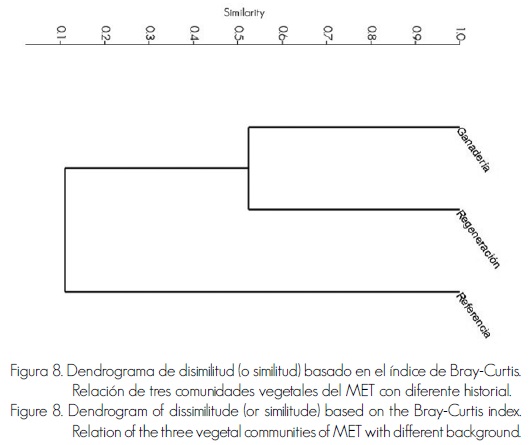
Índice de similitud (o disimilitud) Bray–Curtis. Las áreas con mayor similitud entre sí fueron las de Ganadería y Regeneración, las cuales tuvieron un porcentaje de similitud de poco más de 50%; por otra parte, estas mismas áreas, apenas registraron poco más de 10% de similitud en relación con el área de Referencia (Figura 8). De manera cuantitativa, se evidencia que áreas con similitud de condiciones ambientales (edáficas, topográficas, altitudinales, clima) tienen diferencias en la composición de especies debido a su historial de uso. Esta información concuerda con la de Alanís et al. (2013), quienes evaluaron áreas del MET con diferente historial de uso silvoagropecuario y obtuvieron valores de similitud menores a 28%.
CONCLUSIONES
La actividad ganadera disminuye significativamente la abundancia (ind/ha), dominancia (área de copa) y diversidad alfa (índice de Margalef e índice de Shannon) de las comunidades de especies arbóreas y arbustivas, y las comunidades resultantes muestran una baja similitud de especies, lo cual favorece el establecimiento de especies en las primeras fases sucesionales. Este análisis aporta elementos cuantitativos de la vegetación arbórea y arbustiva en tres comunidades con diferente historial antrópico del Matorral Espinoso Tamaulipeco, que sentarán las bases para futuros programas de manejo y restauración de dicho ecosistema.
AGRADECIMIENTOS
A la Facultad de Ciencias Forestales de la Universidad Autónoma de Nuevo León, por todas las facilidades otorgadas para el establecimiento y desarrollo de la investigación. Al Ing. Francisco Guadalupe Pérez Pérez por su apoyo en las actividades de campo. El estudio aquí descrito fue financiado por el proyecto PROMEP/103.5/12/3585.
REFERENCIAS
Abril, Y. R. 2011. Sistemas agroforestales como alternativa de manejo sostenible en la actividad ganadera de la Orinoquia Colombiana. Revista Sistemas de Producción Agroecológica 2 (1):103-127. [ Links ]
Alanís, E., J. Jiménez, D. Espinoza, E. Jurado, O. A. Aguirre y M. A. González, 2008a. Evaluación del estrato arbóreo en un área restaurada post-incendio en el Parque Ecológico Chipinque. Revista Chapingo. Serie Ciencias Forestales y del Ambiente 14 (2):113-118. [ Links ]
Alanís, E., J. Jiménez, O. A. Aguirre, E. Treviño, E. Jurado y M. A. González. 2008b. Efecto del uso del suelo en la fitodiversidad del matorral espinoso tamaulipeco. Revista Ciencia UANL 11 (1):56-62. [ Links ]
Alanís, E., J. Jiménez, M. A. González, J. I. Yerana, L. G. Cuellar, A. Mora-Olivo. 2013. Análisis de la vegetación secundaria del matorral espinoso tamaulipeco, México. Phyton International Journal of Experimental Botany. 82:221-228. [ Links ]
Arriaga, L. 2009. Implicaciones del cambio de uso de suelo en la biodiversidad de los matorrales xerófilos: un enfoque multiescalar. Investigación ambiental 1 (1):6-16. [ Links ]
Beals, E. W. 1984. Bray-Curtis ordination: an effective strategy for analysis of multivariate ecological data. Advances in Ecological Research 14 (1):55. [ Links ]
Canizales, P. A., E. Alanís, R. Aranda, J. M. Mata, J. Jiménez, G. Alanís, J. I. Uvalle y M. G. Ruíz. 2009. Caracterización estructural del matorral submontano de la Sierra Madre Oriental, Nuevo León. Revista Chapingo. Serie Ciencias Forestales y del Ambiente. 15 (2):115-120. [ Links ]
Espinoza, R., B. y J. Návar. 2005. Producción de biomasa, diversidad y ecología de especies en un gradiente de productividad en el matorral espinoso tamaulipeco del nordeste de México. Revista Chapingo. Serie de Ciencias Forestales y del Ambiente 11 (1):25-31. [ Links ]
Estrada, E., A. D. Yen y J. Villarreal. 2004. Leguminosas del centro del estado de Nuevo León, México. Anales del Instituto de Biología. Serie Botánica 75:73-85. [ Links ]
García, J. y E. Jurado. 2008. Caracterización del matorral con condiciones prístinas en Linares, N. L., México. Ra Ximhai 4 (1):1-21. [ Links ]
González, H., R. Ramírez, I. Cantú, M. Gómez y J. I. Uvalle. 2010. Composición y Estructura de la vegetación en tres sitios del estado de Nuevo León, México. Polibotánica. 29:91-106. [ Links ]
Herrera, A. 2000. La clasificación numérica y su aplicación en ecología. Instituto Tecnológico de Santo Domingo, Santo Domingo, República Dominicana. pp. 221-227. [ Links ]
Jiménez, J., E. Alanís, O. A. Aguirre, M. Pando y M. A. González. 2009. Análisis sobre el efecto del uso del suelo en la diversidad estructural del matorral espinoso tamaulipeco. Madera y Bosques. 15 (3):5-20. [ Links ]
Jiménez, P., J., E. Alanís R., J. L. Ruiz G, M. A. González T., J. I. Yerena Y. y G. J. Alanís F. 2012. Diversidad de la regeneración leñosa del matorral espinoso tamaulipeco con historial agrícola en el noreste de México. Revista Ciencia UANL.15 (58): 66-71. [ Links ]
Magurran, A. 2004. Measuring Biological Diversity. Blackwell Science Ldt. Blackwell Publishing Company. pp. 106-121. [ Links ]
Margalef, R. 1951. Diversidad de especies en las comunidades naturales. Publicaciones del Instituto de Biología Aplicada. 9:5-27. [ Links ]
Molina, V. M.; M. Pando, E. Alanís, P. A. Canizales, H. González y J. Jiménez. 2013. Composición y diversidad vegetal de dos sistemas de pastoreo en el matorral espinoso tamaulipeco del Noreste de México. Revista Mexicana de Ciencias Pecuarias. 4(2):361-371 [ Links ]
Moreno, C. E. 2001. Métodos para medir la biodiversidad. Manual y Tesis SEA. Cooperación Iberoamericana (CYTED), Unesco (Orcyt) y SEA. Vol. 1. Pachuca, Hgo. México. 83 p. [ Links ]
Müeller-Dombois D. and H. Ellenberg. 1974. Aims and methods of vegetation ecology. John Wiley and Sons. New York, NY. USA. 54p. [ Links ]
Organización de las Naciones Unidas para la Agricultura y la Alimentación (FAO). 2009. El estado mundial de la agricultura y la alimentación. La ganadería, a examen. Organización de las Naciones Unidas para la Agricultura y la Alimentación. Roma, Italia. pp. 3-4. [ Links ]
Pequeño, M. A., E. Alanís, J. Jiménez, M. A. González, J. I. Yerena, L. G. Cuellar y G. A. Mora. 2013. Análisis de la restauración pasiva post-pecuaria en el matorral esponoso tamaulipeco del noreste de México. Ciencia UAT. Ciencia UAT 24(2):48-53. [ Links ]
Rzedowski, J. 2006. Vegetación de México. Comisión Nacional para el Conocimiento y Uso de la Biodiversidad. México, D.F. México. 504 p. [ Links ]
Sadeghian, K., S. Rivera y J. M. Gómez. 1999. Impacto de la ganadería sobre las características físicas, químicas y biológicas de suelos en los andes de Colombia. Agroforestería para la producción animal en América Latina. Estudio FAO Producción y Sanidad Animal No. 143. FAO. Roma, Italia. pp. 123-142. [ Links ]
Secretaría de Agricultura, Ganadería, Desarrollo Rural, Pesca y Alimentación (SAGARPA). 2006. Programa sectorial de agricultura, ganadería, desarrollo rural, pesca y alimentación 2001-2006. México, D.F. México. 26 p. [ Links ]
Secretaría de Agricultura, Ganadería, Desarrollo Rural, Pesca y Alimentación. (SAGARPA). 2009. Nuevo León: Agenda de innovación agroindustrial. México, D.F. México. 30 p. [ Links ]
Secretaría de Medio Ambiente y Recursos Naturales (SEMARNAT). 2006. El Medio Ambiente en México 2005: en resumen. México, D.F. México. 91p. [ Links ]
Shannon, C. E. and W. Weaver. 1948. The mathematical theory of communication. University of Illinois Press. Urbana-Champaign, IL USA. pp. 134-154. [ Links ]
Steinfeld, H., P. Gerber, T. Wassenaar, V. Castel, M. Rosales and C. de Haan. 2006. Livestock’s long shadow. Environmental issues and options. FAO, Roma, Italia. 5-6 pp. [ Links ]
Villanueva, C., C. J. Sepúlveda y M. Ibrahim. 2011. Manejo agroecológico como ruta para lograr la sostenibilidad de fincas con café y ganadería. Serie Técnica. Informe Técnico 387. CATIE.Turrialba, Costa Rica. pp. 96-97. [ Links ]
Whittaker, R. H. 1972. Evolution and measurement of species diversity. Taxon 21:213–251. [ Links ]

