










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista mexicana de ciencias forestales
versão impressa ISSN 2007-1132
Rev. mex. de cienc. forestales vol.3 no.13 México Set./Out. 2012
Artículos
Ecuaciones alométricas para estimar biomasa aérea de Prosopis laevigata (Humb. & Bonpl. ex Willd.) M.C. Johnst.
Allometric equations to estimate aerial biomass of Prosopis laevigata (Humb. & Bonpl. ex Willd.) M.C. Johnst.
Jorge Méndez González1, Osvaldo Augusto Turlan Medina1, Julio Cesar Ríos Saucedo2 y Juan Abel Nájera Luna3
1 Universidad Autónoma Agraria Antonio Narro. Correo-e: jorgemendezgonzalez@gmail.com
2 Campo Experimental Valle del Guadiana. CIR-Norte Centro. INIFAP
3 División de Estudios de Posgrado e Investigación. Instituto Tecnológico de El Salto
Fecha de recepción: 28 de marzo de 2012;
Fecha de aceptación: 10 de septiembre de 2012
RESUMEN
La medición y evaluación de la biomasa aérea de árboles juega un papel importante en el manejo de los recursos forestales. Existen muchos modelos alométricos para especies tropicales y subtropicales, pero muy pocos para las zonas semiáridas. Un total de 144 árboles de Prosopis laevigata, de siete rodales naturales localizados en el noreste de México, fueron muestreados por un método destructivo con la finalidad de desarrollar ecuaciones para la predicción de la biomasa total. La selección de los individuos cubrió varios intervalos de diámetro basal (5.2 a 41.8 cm) y de altura (1.4 a 9.7 m). Se contrastaron procedimientos de ajuste no-lineal y lineal de la ecuación alométrica y = a·xb + ε, para estimar la biomasa aérea de P. laevigata. Aunque, el procedimiento no-lineal tuvo el mejor ajuste (R2 = 0.95 y P < 0.001) vs el lineal (R2 = 0.84 y P < 0.001), los resultados demuestran que la transformación logarítmica y el uso de un factor de corrección en alometría mejora de forma significativa la predicción de la biomasa. Se concluye que los modelos de regresión obtenidos al usar el diámetro basal pueden ser aplicados para la estimación de biomasa total, en árboles de Prosopis laevigata. Así mismo, el número mínimo de observaciones necesarias para calcular los parámetros a y b, con varianza mínima es de 40 ejemplares.
Palabras clave: Alometría, estimación de biomasa, mezquite, manejo forestal, modelo lineal, modelo no-lineal.
ABSTRACT
The measurement and assessment of aboveground tree biomass plays a key role in the management of forest resources. Many allometric models exist for tropical and subtropical species, but only a few studies for species of semiarid zones. A total of 144 Prosopis laevigata trees from seven native stands (sites) located in northeast Mexico, were destructively sampled to develop total biomass prediction equations. Sampling covered various ranges of basal diameter (5.2 to 41.8 cm) and height (1.4 to 9.7 m). Here, we contrast nonlinear and linear fitting approaches of the allometric equation y = a·xb + ε, for estimating aboveground biomass of Prosopis laevigata. Although nonlinear procedure had the best fits (R2 = 0.95 and P < 0.001) vs linear (R2 = 0.84 and P < 0.001), the results highlight that the log-transform and the use of a weighted correction factor in allometry, improves significantly the biomass prediction of this specie. It is concluded that the obtained regression models using basal diameter, can be applied for the estimation of total biomass in Prosopis laevigata trees, but also that the minimum number of observations needed is 40 harvested trees to calculate parameters a y b with the least variance.
Key words: Allometry, biomass estimates, mesquite, forest management, nonlinear and linear model.
INTRODUCCIÓN
El dióxido de carbono atmosférico (CO2) es un componente esencial del proceso de fotosíntesis y, en consecuencia, de la vida; al mismo tiempo, actúa como una capa protectora del planeta, al cual mantiene con su temperatura actual: ~15 °C que, de no existir, alcanzaría hasta -18 °C (IPCC, 2001). No obstante, sus altos niveles atmosféricos han alterado el sistema climático. Así, el carbono (C) está presente en la atmósfera (750 Gt C); en los ecosistemas terrestres (plantas, 601 Gt C; suelo, 1,580 Gt C); y en los océanos (38,100 Gt C) (Beedlow et al., 2004). Se ha documentado que el CO2 atmosférico se ha incrementado en más de 100 ppm (36%) en los últimos 250 años, es decir, de 275 (era preindustrial) hasta 390 ppm (2010), lo que representa casi 2.0 ppm año-1 (Peters et al., 2011) y 50 ppm en los últimos 30 años (Soon et al., 1999). De acuerdo a las tendencias industriales, se pronostican concentraciones de 540 a 970 ppm de CO2 para finales del presente siglo (Prentice et al., 2000), que producirá un aumento en la temperatura media global de entre 3 y 5 °C (IPCC, 2007); cabe señalar que esta ha variado cerca de 1 °C en mil años (Soon et al., 1999).
Los ecosistemas terrestres son importantes en el ciclo global del carbono, ya que almacenan en su biomasa 638 Gt C, poco menos que el C atmosférico (Brown, 1997a), cuyos flujos son de 120 Gt C año-1 durante el proceso de la fotosíntesis (Beedlow et al., 2004); sin embargo, de 2005 a 2010, en la biomasa forestal, el C ha disminuido 0.5 Gt C año-1 (FAO, 2011) debido a la pérdida de bosques. Gran parte de los estudios de biomasa se han realizado en las zonas tropicales (Brown et al., 1989; Chave et al., 2004) y templadas (Návar, 2009; Mendoza y Galicia, 2010).
Las zonas áridas de México representan más de 62 millones de hectáreas (Masera et al., 1997); no obstante, de manera independiente a la ubicación geográfica, son pocos los estudios sobre almacenes de C que se han realizado en ellas (Gaillard et al., 2002; Méndez et al., 2006; Iglesias y Barchuk, 2010). Aun así, se ha documentado que el reservorio más importante de carbono en el país se ubica en la vegetación semiárida: 5 Gt CO2; es decir, 31% del carbono (Masera et al., 1997). La cuantificación de biomasa es de gran importancia, puesto que aproximadamente 50% de ella corresponde a este elemento (Brown, 1997b).
El género Prosopis es nativo y representativo de las zonas áridas y semiáridas (Pasiecznik et al., 2004; López et al., 2006), constituye un grupo primitivo de la subfamilia Mimosoideae, tal vez originado en África tropical (Burkart, 1976). En México, se distribuye, principalmente, en los estados del norte y del centro (Rzedowski, 1988), y está representado por 11 especies (Palacios, 2006). Es un árbol espinoso, termo-xerófilo, perenne y alcanza hasta 12 m de altura; pertenece a la familia Leguminosae (Fabaceae) (Rzedowski, 1988; Pasiecznik et al., 2004).
Dada su importancia, en cuanto al uso, distribución, capacidad para almacenar CO2 por periodos prolongados, y a la escasez de estudios antes mencionada es relevante e ineludible generar modelos para cuantificar su biomasa a escala geográfica local y regional.
La hibridación interespecífica en Prosopis, de manera especial en la sección Algarobia, genera confusión y dificulta su clasificación taxonómica (Rzedowski, 1988; Palacios, 2006). Por ello, para identificar la especie de los árboles analizados se llevaron a cabo estudios con marcadores morfológicos, cuyos resultados indicaron se trataba de Prosopis laevigata (Rosales et al., 2011).
Los objetivos de la presente investigación fueron:1) generar ecuaciones alométricas para cuantificar biomasa aérea de árboles individuales de Prosopis laevigata (Humb. & Bonpl. ex Willd.) M.C. Johnst. en siete poblaciones distribuidas en el norte-centro de México; 2) evaluar la estimación de biomasa de Prosopis laevigata al ajustar el modelo alométrico en forma no-lineal y lineal; y 3) analizar los estadísticos de regresión en función del tamaño de muestra, para determinar el posible tamaño óptimo de observaciones para construir modelos alométricos en Prosopis spp.
MATERIALES Y MÉTODOS
Área de estudio
La investigación se realizó en cuatro estados del norte-centro de México: Durango, Zacatecas, Coahuila y Chihuahua. Y se incluyeron siete localidades (Figura 1) conformadas por ejidos, predios particulares y campos experimentales del Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP). Los suelos predominantes son del tipo Regosol (calcárico y eútrico), de textura media (INIFAP-CONABIO, 1995). De acuerdo a García-CONABIO (1998), el clima es BS 1 hw (semiárido - semicálido), con temperatura media de 18 °C, precipitación anual de 125 a 600 mm. El intervalo altitudinal de los sitios fue de 1,097 a 2,001 m (INEGI, 1990) (Cuadro 1).
Método destructivo para obtener biomasa de Prosopis spp.
Se utilizó el método propuesto por Schlegel et al. (2000) que consiste en derribar el árbol, pesar en fresco sus componentes (hojas, ramas y fuste) y secarlos en estufa. Se seleccionaron individuos, sanos, distribuidos en el área y representativos de las categorías de diámetro y altura (Brown et al., 1989), a los cuales se les midió, en pie el diámetro basal (DB), la altura total (H), el diámetro de copa (DC) y el número de ramas (NR). El uso de DB obedeció a que la mayoría de los individuos evaluados presentaron ramificaciones desde la base, lo que dificultó el uso del diámetro normal. Posterior al derribo, los componentes fueron pesados en verde con una báscula romana marca Rotter de 50, 25 kg de capacidad y 100 g de precisión. De cada uno de ellos, se obtuvieron cuatro muestras para calcular el contenido de humedad.
En el laboratorio, el material se secó en una estufa marca Riossa a 105°C, hasta obtener el peso constante (Brown, 1997b; Schlegel et al., 2000); después, de la razón entre el peso seco y el húmedo se calculó la biomasa seca. La biomasa total por árbol (BT), se obtuvo al sumar la biomasa de todos los componentes.
Modelo alométrico para cuantificar biomasa de Prosopis laevigata
La biomasa total de los árboles de cada sitio se estimó con la aplicación del modelo alométrico (Smith y Brand, 1983; Sprugel, 1983; Brown et al., 1989; Tausch, 1989; Zianis y Mencuccini, 2004), de la forma:
y = α·xb + ε (1)
Ajustada en forma logarítmica:
lny = ln(α) + b·ln(x) + ε (2)
Donde:
α = Exp(a) en ecuación 2
y = Biomasa aérea total (kg)
x = Diámetro basal (cm)
αy b = Parámetros del modelo
ln = Logaritmo natural
ε = Error.
La transformación logarítmica corrige la heterogeneidad de la varianza de y con respecto αx (Brown et al., 1989), al incrementar la validez estadística del modelo debido a la homogenización de la varianza (Sprugel, 1983; Zianis y Mencuccini, 2004). Se obtuvo la R2 ajustada, conocida como índice de ajuste, para evaluar la predicción de los modelos; así como otros estadísticos: error, coeficiente de variación y significancia del modelo (Návar, 2010). El modelo se ajustó con el programa SAS (versión 9.0) al utilizar ‘proc nlin' (con derivadas y el algoritmo de Newton - Raphson) en la ecuación (1) y ‘proc reg' para la ecuación (2), en ambas se resolvió con cuadrados mínimos (Brown et al., 1989; Gaillard et al., 2002).
Cuando las unidades originales son re-transformadas en el modelo (2) los valores más grandes se subestiman (Brown et al., 1989); para corregirlo, Sprugel (1983) propuso la aplicación de un factor de corrección ponderado, el cual se expresa por:

Donde:
FC = Factor de corrección
exp = Exponencial
SEE = Error estándar de la estimación.

Donde:
yobs = Biomasa observada
yest = Biomasa estimada
N = Número de observaciones.
Modelo global para cuantificar biomasa de Prosopis laevigata en todos los sitios
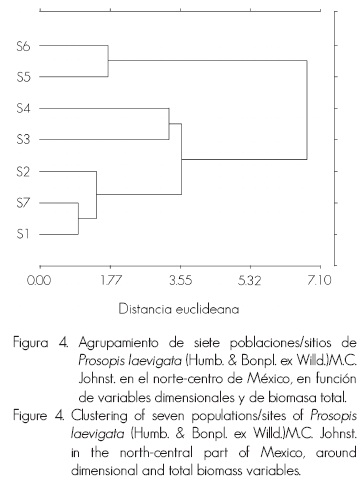
Las ecuaciones 1 y 2 se utilizaron en todo el conjunto de datos de biomasa de Prosopis laevigata (144 árboles) para verificar la aplicabilidad de una ecuación global. El ajuste de los modelos consideró los criterios estadísticos citados con anterioridad. Además, se realizó un análisis multivariado de conglomerados, en el cual se usaron las variables estandarizadas: DB, H, DC, NR y BT, a través de la distancia euclidiana y agrupamiento jerárquico de Ward (Ward, 1963).
Así mismo, la ecuación 1 se ajustó a diferentes tamaños de muestra n (10, 20, 30... 130 y 140) obtenidos al azar con reemplazo. Los estadísticos R2, coeficiente de variación y parámetros de regresión (a y b) se consideraron para determinar las mejores estimaciones de biomasa, en función del tamaño de muestra y, con estas últimas, probar la teoría fractal de que b = 2.67 (Enquist et al., 1998).
RESULTADOS Y DISCUSIÓN
El diámetro basal de Prosopis laevigata varió de 5.2 cm (S2 y S7) a 41.8 cm (S5); es decir, comprendió distintas categorías diamétricas. Se observa que sus poblaciones, localizadas en los cuatro estados, poseen estructuras dimensionales diferentes (Cuadro 1) pero, en general, similares a los estudios realizados con Prosopis glandulosa, var. torreyana (L.D. Benson) M. C. Johnst. (diámetro de 4 a 45 cm) en el noroeste de México. En el matorral espinoso Tamaulipeco, Návar et al. (2004) registraron un diámetro basal de hasta 24 cm en árboles de Prosopis glandulosa Torr. y de 34 cm en Prosopis laevigata. Al norte de Argentina, Prosopis flexuosa DC. y P. torquata DC. sus diámetros basales alcanzan valores de 35.5 y 29.3 cm, respectivamente (Iglesias y Barchuk, 2010). Por otra parte, se apreciaron diferencias importantes en la biomasa de los árboles entre sitios de muestreo; por ejemplo, diámetros basales similares máximos (S2 y S7; S3 y S4) poseen más de 100 kg de diferencia (Cuadro 1), lo cual pudo deberse al aprovechamiento parcial o a las actividades silvícolas diferenciales practicadas en las poblaciones de Prosopis laevigata.
Estimaciones de biomasa de Prosopis laevigata en sitios individuales
El modelo alométrico ajustado en las formas 1 y 2 presentó alta significancia estadística (P < 0.0001) en los sitios evaluados. El modelo 1 mostró los mejores ajustes con valores de R2 > 0.90 en todos los casos (Cuadro 2), con un máximo de 0.98 (S2) y 0.97 (S1 y S6). Por el contrario, en el modelo 2 (logarítmico) se registraron R2 más bajas (hasta 0.73 en S7); es decir, un ajuste menor, 10% en promedio, en comparación con el modelo 1.
El análisis de resultados evidencia que la mayor variación de biomasa en los árboles de Prosopis laevigata se observa en S5, según se indica por el CV de 39.45 y 56.89% en los modelos 1 y 2; mientras que, la variación más pequeña se determinó en individuos con menor cantidad de biomasa (S1 y S2), con alrededor de 20% de CV (Cuadro 2). El menor CV promedio (28.87%) obtenido con el modelo 1, se traduce, también, en un mejor ajuste para estimar la biomasa aérea total en esta especie (Cuadro 2). Los resultados demuestran variaciones importantes en la biomasa estimada; por ejemplo, a un diámetro de 40 cm se tienen diferencias desde 200 hasta 300 kg (BT, S1 - BT con respecto a los otros sitios), excepto en S7, y de 100 hasta 200 kg calculados con los modelos 1 y 2, respectivamente (figuras 2C y 2D). Lo anterior podría deberse a la intensidad de aprovechamiento (podas o cortas de saneamiento), a nivel árbol.

En Prosopis glandulosa var. torreyana, Méndez et al. (2006) consignan una R2 de 0.87 y CV de más de 50%, al usar el modelo 1. A partir del ajuste de la ecuación logarítmica, Iglesias y Barchuk (2010) calcularon una R2 de 0.87 y 0.88 para Prosopis torquata y Prosopis flexuosa DC., pero con CV de hasta 80%. La aplicación del factor de corrección FC, que promedia 1.043 a la ecuación logarítmica, aumentó las estimaciones de biomasa de 2.8% (S2) a 5.9% (S3) (Cuadro 2); sin embargo, el modelo 1 predijo mejor la biomasa de Prosopis laevigata que el modelo 2. Chave et al. (2004) han señalado que el FC en bosques tropicales es de 1.068 ± 0.022 y, en cuanto a bosques templados, Návar (2010) indica 1.065 ± 0.072 (media ± desviación estándar), con ello se demuestra que el FC es similar casi en todas las especies. Návar et al. (2004) sostienen que en Prosopis se obtienen estimaciones sesgadas de biomasa con el modelo 1. Se ha documentado, en distintas especies, que la varianza con el modelo 1 se incrementa con el tamaño del árbol (Tausch, 1989; Brown et al., 1989), lo cual le resta validez al modelo. Algunos autores (Tausch, 1989; Litton y Kauffman, 2008) prefieren usar la forma no-lineal para estimar la biomasa aérea en especies vegetales, siempre y cuando se satisfaga la varianza común.
Un modelo global para la estimación de la biomasa de Prosopis laevigata
En el análisis global de los datos de biomasa de Prosopis laevigata del área de estudio, se observó que el mejor ajuste se alcanzó con la ecuación 1 (R2 = 0.91), que fue muy superior a la obtenida con el modelo 2 (R2 = 0.69), aunque ambas tuvieron alta significancia (P < 0.0001) (Cuadro 3), incluso 0.04 unidades de R2 más que el promedio de los siete sitios (Cuadro 2). Tausch (1989) ha citado mejores ajustes con el modelo 1, y ha señalado que esto depende de la especie. El considerar todos los datos de biomasa de Prosopis laevigata en un solo modelo (lineal y no-lineal) amplió la variación en y (Figura 3A), al incrementar de forma substancial el error estándar (hasta 38.32 kg: modelo lineal) y el coeficiente de variación (54.09%: modelo lineal), con 10.71 kg y 18.3% respectivamente, más que el promedio de los sitios (cuadros 2 y 3).
El factor de corrección (1.198) derivado del análisis global (Cuadro 3) mejoró las estimaciones de biomasa, las cuales fueron semejantes a las obtenidas con el modelo 1 (Figura 3A).
Las diferencias más significativas de biomasa (modelo no-lineal - modelo lineal), de 100 kg a un diámetro de 40 cm obtenidos con el modelo global, se registraron en árboles de mayores dimensiones, mismas que se incrementan de modo exponencial, a partir del punto de inflexión entre ellos: a 23 cm de diámetro (Figura 3B). La biomasa promedio de N = 144 árboles fue de 60.85 kg y el error estándar promedio por árbol estimado con  (Chave et al., 2004) resultó de 4.83 kg, que puede ser calculado de forma individual con:
(Chave et al., 2004) resultó de 4.83 kg, que puede ser calculado de forma individual con:
y' = 0.2638x1.0246 (R2 = 0.9077 y P < 0.001)
Donde:
y' = Error estándar árbol-1 (kg)
x = DB (cm).
Lo anterior expresa el buen ajuste del modelo logarítmico, incluso la aplicabilidad de ambos para estimar la biomasa aérea de Prosopis laevigata en ciertas categorías diamétricas. Resultados que concuerdan con lo registrado por Tausch (1989) y Návar (2009); al respecto, este último recomienda usar dos ecuaciones: una para diámetros de árboles pequeños y otra para diámetros de árboles grandes.
El ajuste global fue bueno, si se considera que los datos representan poblaciones de cuatro estados del norte-centro de México (Figura 1). No obstante, los individuos de los sitios evaluados muestran diferencias substanciales de biomasa aérea en las mismas categorías diamétricas, por lo que se decidió clasificar los sitios para proponer un mejor uso de los resultados. El análisis multivariado de conglomerados arrojó una correlación cofenética de 0.769, que resultó en la conformación de algunos grupos y subgrupos (Figura 4), semejantes a sitios geográficamente cercanos (Figura 1). Así, se evidencia una clara similitud entre S1 y S7, S5 y S6 (pares de curvas: parte inferior y superior Figura 2A), cuya semejanza entre curvas modelo no-lineal/dendrograma es concordante por tratarse de datos no logarítmicos en Figura 2A.
Estadísticos de regresión en función de n
La aplicación del modelo 1 a los datos de biomasa con diferentes tamaños de muestra presentó, en todos los casos, alta significancia estadística (P < 0.0001). El mejor ajuste del modelo se obtuvo con n = 20 (R2 = 0.96 y CV = 32.47%); por el contrario, el ajuste más bajo (R2 = 0.88 y CV = 54.78%) se calculó con n = 100 (Cuadro 4). De acuerdo a los resultados, se observó un aumento de b al mismo tiempo que CV (r = 0.5914; P = 0.0259); un incremento de Sxy (r = 0.5272; P = 0.0527) y de CV (r = 0.6066; P = 0.0214) conforme aumenta n. Por último, se registró una mayor variación en la estimación de biomasa (CV) de Prosopis laevigata al aumentar el diámetro basal promedio  (r = 0.6491; P = 0.0120). Los resultados demuestran una correlación estrecha entre los parámetros a y b al seguir una función de la forma:
(r = 0.6491; P = 0.0120). Los resultados demuestran una correlación estrecha entre los parámetros a y b al seguir una función de la forma:
b = 1.5855a-0.1381 (R2 = 0.9933 y P < 0.0001)
a = 26.995b-7.1950 (R2 = 0.9933 y P < 0.0001)
Relaciones similares han sido documentadas por Zianis y Mencuccini (2004) y Návar (2009).
Los estadísticos R2 y CV muestran una tendencia logarítmica con respecto a n, por ello, se aplicó el modelo del tipo:
y = a + b · ln(x)
Donde:
"y" representa R2 (Figura 5A) y CV (Figura 5B)
"x" = Tamaño de muestra.
No obstante, para magnificar la tendencia se obtuvo la primera derivada del modelo:
dy/dx = b/x.
Los resultados revelan una estabilización de ambos estadísticos (R2 y CV), cuando n se aproxima a 40 (Figura 5), es decir, con n > 40 se obtienen los mejores ajustes del modelo para cuantificar biomasa aérea total de Prosopis laevigata. La varianza mínima, representada por el error de a y b (barras, figuras 5C y 5D), disminuye después de n = 40 y se mantiene constante después de este valor. Al respecto, Brown et al. (1989) documentaron estudios de biomasa de cuatro y cinco árboles; mientras que, Zianis et al. (2005) registraron que en Europa, de más de 600 modelos generados para estimar la biomasa, 30% utilizan entre 11 y 20 árboles. Al analizar esta misma relación (Figura 5) con más de 594 datos de biomasa, Návar (2010) indica que el tamaño de muestra ideal para construir modelos es de 60 árboles.
Teoría fractal (b = 2.67)
Se generó la hipótesis de que la media de b de la relación alométrica y = a·xb es 2.67 (~ 8/3) (Enquist et al., 1998; Zianis y Mencuccini, 2004; Návar, 2010). Se realizaron pruebas de "t" para una media de dos colas (95%) con la finalidad de probarla para un posible uso de b en futuras estimaciones de biomasa y, por ende, en Prosopis spp. El promedio del parámetro b de la ecuación 1, el cual usó diferente n de esta investigación fue de 2.4436 ± 0.240 (Cuadro 4). La prueba de "t" aportó información suficiente para aceptar la hipótesis nula:
µ1 - µ2
Donde:
µ1 = b de este estudio y
µ2 = b de otros estudios.
Lo anterior demostró que es similar (t = 1.032; P = 0.3035) al parámetro b de Smith y Brand (1983), 2.4925 ± 0.6955, estimado a partir de 215 modelos para 98 especies de herbáceas y arbustivas de Estados Unidos de América y Canadá; igual (t = 1.199; P = 0.2520) al citado por Zianis y Mencuccini (2004), 2.3679 ± 0.016, calculado en 279 estudios de biomasa y semejante al registrado por Návar (2009) 2.3785 ± 0.051 en 78 ecuaciones alométricas (t = 0.025; P = 3239) de grupos de especies localizadas en América Latina. No obstante, este y otros trabajos, demuestran que b es diferente (t = -3.53; P = 0.0037) al valor 2.67 expuesto en la teoría fractal (Enquist et al., 1998).
CONCLUSIONES
El diámetro basal está correlacionado de manera importante con la biomasa aérea de Prosopis laevigata; por lo tanto, puede usarse para estimar almacenes de carbono en la región de estudio y en Prosopis spp.
El modelo no-lineal sobreestima la biomasa de Prosopis laevigata en árboles de mayores dimensiones, al contrario de lo que ocurre con el modelo lineal; de tal forma que, se recomienda usar este último; además de, que su transformación y la aplicación del FC incrementa la validez estadística del modelo. El FC aplicado al modelo lineal incrementó las estimaciones de biomasa en más de 5%.
Existen diferencias en las características dasométricas entre sitios y, en consecuencia, en las curvas de biomasa, por lo que se sugiere que, a pesar del buen ajuste del modelo global, se aplique la ecuación específica del sitio.
El análisis de parámetros (a y b) y estadísticos (R2 y CV) del modelo en función de n indica que el mejor ajuste se obtiene con n > 40, el cual pudiera ser el tamaño óptimo para construir modelos alométricos en el género Prosopis.
El parámetro b (2.4436 ± 0.240) de Prosopis spp. calculado en la presente investigación es diferente a lo expuesto en la teoría fractal, pero similar al de la mayoría de las especies documentadas en la literatura.
AGRADECIMIENTOS
Al Fondo Sectorial CONAFOR - CONACYT, Proyecto clave "Generación validación y/o desarrollo de tecnología para el manejo sustentable de la cadena productiva del mezquite en la región norte centro de México." Clave: 115942. A la Universidad Autónoma Agraria Antonio Narro, por el apoyo complementario para el análisis de datos.
REFERENCIAS
Beedlow, P. A., D. T. Tingey, D. H. Phillips, W. E. Hogsett and D. M. Olszyk. 2004. Rising atmospheric CO2 and carbon sequestration in forests. Frontiers in Ecology and the Environment. 2 (6):315–322. [ Links ]
Brown, S. 1997a. Los bosques y el cambio climático: el papel de los terrenos forestales como sumideros de carbono. Congreso Forestal Mundial. Antalya Turquía. (1):107-116. [ Links ]
Brown, S. 1997b. Estimating biomass and biomass change of tropical forests. A primer. FAO Forestry Paper 134. Rome Italy 55 p. [ Links ]
Brown, S., A. J. R. Gillespie and A. E. Lugo. 1989. Biomass estimation methods for tropical forest with applications to forest inventory data. Forest Science. 35(4):881-902. [ Links ]
Burkart, A. 1976. A monograph of the genus Prosopis (Leguminosae subfam. Mimosoideae). Journal of the Arnold Arboretum. 57(3):217-249. [ Links ]
Chave, J., R. Condit, S. Aguilar, A. Hernández, S. Lao and R. Pérez. 2004. Error propagation and scaling for tropical forest biomass estimates. Philosophical Transactions of the Royal Society London B. Biological Science. 359:409–420. [ Links ]
Enquist, B. J., J. H. Brown and J. B. West. 1998. Allometric scaling of plant energetics and population density. Nature. 395:163–165. [ Links ]
Organización de las Naciones Unidas para la Agricultura y la Alimentación (FAO). 2011. Situación de los bosques del mundo. Roma, Italia. 64-66 pp. [ Links ]
Gaillard de Benitez, C., M. Pece, M. de Juárez, S. Vélez, A. Gómez y M. Zárate. 2002. Determinación de funciones para la estimación de biomasa aérea individual en Jarilla (Larrea divaricata) de la provincia de Santiago del Estero, Argentina. Foresta Veracruzana. 4 (2):23-28. [ Links ]
García, E. - Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (CONABIO). 1998. Climas (Clasificación de Köppen, modificado por García). Escala 1:1 000 000. México. [ Links ]
Iglesias, M. R. y A. H. Barchuk. 2010. Estimación de la biomasa aérea de seis leguminosas leñosas del Chaco Árido (Argentina). Ecología Austral. 20(1):71-79. [ Links ]
Instituto Nacional de Estadística, Geografía e Informática (INEGI). 1990. Hipsometría. Extraído de Hipsometría y Batimetría. Atlas Nacional de México. Vol. I. Escala 1:4 000 000. Instituto de Geografía, UNAM. México, D.F. México. [ Links ]
Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP) - Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (CONABIO). 1995. Edafología. Escalas 1: 250 000 y 1:1 000 000. México. [ Links ]
Intergovernmental Panel on Climate Change (IPCC). 2001. Climate Change 2001: The Scientific Basis. Contribution of Working Group I to the Third Assessment Report of the Intergovernmental Panel on Climate Change. In: Houghton, J. T., Y. Ding, D. J. Griggs, M. Noguer, P. J. Van Der Linden, X. Dai, K. Maskell and . A. Johnson (Eds.). Cambridge University Press. Cambridge, UK. and New York, NY, USA. 881 p. [ Links ]
Litton, C. M. and J. B. Kauffman. 2008. Allometric models for predicting aboveground biomass in two widespread woody plants in Hawaii. Biotropica. 40 (3):313-320. [ Links ]
López F., F. M. Goycoolea, M. A. Valdéz y A. M. Calderón. 2006. Goma de mezquite: una alternativa de uso industrial. Interciencia. 31 (3):83-189. [ Links ]
Masera O., R., M. J. Ordoñez and R. Dirzo. 1997. Carbon emissions from Mexican Forests: Current Situation and Longterm Scenarios. Climatic Change. 35 (3):265-295. [ Links ]
Méndez G., J., A. Santos, J. A. Nájera y V. González. 2006. Modelos para estimar volumen y biomasa de árboles individuales de Prosopis glandulosa, var. torreyana en el Ejido Jesús González Ortega No 1, Mpio. de Mexicali, B.C. Agrofaz. 6 (2):226-235. [ Links ]
Mendoza P., A. and L. Galicia. 2010. Aboveground and belowground biomass and carbon pools in highland temperate forest landscape in Central Mexico. Forestry. 83 (5):497-506. [ Links ]
Návar J., E. Méndez, A. Nájera, J. Graciano, V. Dale and B. Parresol. 2004. Biomass equations for shrub species of Tamaulipan thornscrub of North-eastern Mexico. Journal of Arid Environments. 59:657–674. [ Links ]
Návar, J. 2009. Biomass component equations for Latin American species and groups of species. Annals Forest Science. 66(2): 208-216. [ Links ]
Návar, J. 2010. Alometría para biomasa en especies arbóreas del noroeste de México. Tropical and Subtropical Agroecosystems. 12:507-519. [ Links ]
Palacios, R. A. 2006. Los mezquites Mexicanos: biodiversidad y distribución geográfica. Boletín de la Sociedad Argentina Botánica. 41 (1-2):99 –121. [ Links ]
Pasiecznik, N. M., P. J. C. Harris and S. J. Smith. 2004. Identifying tropical Prosopis species: A field guide. HDRA Publishing. Coventry, UK. 29 p. [ Links ]
Peters, G. P., G. Marland, C. Quéré, T. Boden, J. G. Canadell and M. R. Raupach. 2011. Rapid growth in CO2 emissions after the 2008-2009 global financial crisis. Nature Climate Change. 2:2-4. [ Links ]
Prentice, I. C., M. Heimann and S. Stich. 2000. The carbon balance of terrestrial biosphere: Ecosystem models and atmospheric observations. Ecology applied. 10:1553-1573. [ Links ]
Rosales S., R., L. M. Valenzuela, J. C. Ríos, R. Jiménez y J. M. Ibarra. 2011. Diversidad genética en poblaciones naturales de mezquite del norte-centro de México. In: Ríos S., J. C., R. Trucíos, L. M. Valenzuela, G. Sosa y R. Rosales (Eds.). Importancia de las poblaciones de mezquite en el norte-centro de México. Libro Técnico. No. 8. INIFAP, CENID – RASPA, Gómez Palacio Durango. Dgo, México. 220 p. [ Links ]
Rzedowski, J. 1988. Análisis de la distribución geográfica del complejo Prosopis (Leguminosae, Mimosoideae) en Norteamérica. Acta Botánica Mexicana. 3:7-19. [ Links ]
System Statistical Analysis (SAS). 2008. SAS/STAT® 9.2 User's Guide. Cary, NC: SAS Institute Inc. Cary. NC, USA. [ Links ]
Schlegel, B., J. Gayoso y J. Guerra. 2000. Manual de procedimientos muestreos de biomasa forestal. Proyecto Medición de la capacidad de captura de carbono en bosques de Chile y promoción en el mercado mundial. FONDEF- UACH- INFOR. Valdivia, Chile. 20 p. [ Links ]
Smith, W. B. and G. J. Brand. 1983. Allometric biomass equations for 98 species of herbs, shrubs and small trees. U. S. Forest Service. Research Note: NC-299. Saint Paul. MN. USA. 8p. [ Links ]
Soon, W., S. L. Baliunas, A. B. Robinson and Z. W. Robinson. 1999. Environmental effects of increased atmospheric carbon dioxide. Climate Research. 13:149-164. [ Links ]
Sprugel, D. G. 1983. Correcting for bias in log-transformed allometric equations. Ecology. 64 (1):209-210. [ Links ]
Tausch, R. J. 1989. Comparison of regression methods for biomass estimation of sagerrush and runchgrass. Great Basin Naturalist. 49 (3):373-380. [ Links ]
Ward, J. H. Jr. 1963. Hierarchical grouping to optimize an objective function. Journal of American Statistical Association. 58:236-244. [ Links ]
Zianis, D. and M. Mencuccini. 2004. On simplifying allometric analyses of forest biomass. Forest Ecology and Management. 187:311-332. [ Links ]
Zianis, D., P. Muukkonen, R. Mäkipää and M. Mencuccini. 2005. Biomass and stem volume equation for tree species in Europe. Silvia Fennica, Monographs No. 4. Tampere, Finland. 63 p. [ Links ]

