










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista mexicana de ciencias forestales
versão impressa ISSN 2007-1132
Rev. mex. de cienc. forestales vol.3 no.13 México Set./Out. 2012
Artículos
Variación morfológica altitudinal entre poblaciones de Pinus devoniana Lindl. y la variedad putativa cornuta en Michoacán
Altitudinal morphological variation among Pinus devoniana Lindl. Populations and putative variety cornuta in state of Michoacan
Cuauhtémoc Sáenz-Romero1, Selene Aguilar-Aguilar2, Miguel Ángel Silva-Farías3, Xavier Madrigal-Sánchez2, Sabina Lara-Cabrera2 y Javier López-Upton4
1 Instituto de Investigaciones Agropecuarias y Forestales, Universidad Michoacana de San Nicolás de Hidalgo (UMSNH). Correo-e: csaenzromero@gmail.com
2 Facultad de Biología, Universidad Michoacana de San Nicolás de Hidalgo
3 Facultad de Ingeniería en Tecnología de la Madera, Universidad Michoacana de San Nicolás de Hidalgo
4 Programa Forestal, Colegio de Postgraduados.
Fecha de recepción: 26 de abril de 2011;
Fecha de aceptación: 18 de agosto de 2012
RESUMEN
La comprensión de los patrones de variación morfológica a lo largo de gradientes altitudinales es útil para distinguir entidades taxonómicas y tomar decisiones de manejo. Conos, semillas, fascículos y yemas fueron analizados en individuos de 16 poblaciones naturales de Pinus devoniana (ex P. michoacana). El material se recolectó a lo largo de un transecto de 1,600 a 2,450 msnm, a razón de una población cada 50 m, cerca de la ciudad de Morelia, en el estado de Michoacán. Se realizaron análisis univariados (ANOVA) y multivariados (componentes principales y de conglomerados) para determinar las variedades putativas presentes. Los resultados evidenciaron la existencia de un patrón altitudinal significativo de variación morfológica, en el cual las poblaciones de localidades bajas tienen hojas, semillas y conos más largos que las desarrolladas en lugares con altitudes medias y altas. El análisis multivariado discriminó dos grupos evidentemente distintivos: uno compuesto por individuos de poblaciones originadas entre 1,600 y 1,850 msnm: Pinus devoniana var. devoniana (equivalente a P. michoacana típica), y otro en el que se integraron ejemplares de poblaciones procedentes de sitios ubicados en un intervalo de 1,950 a 2,450 msnm: Pinus devoniana var. cornuta (ex P. michoacana var. cornuta). Así, se sugiere la separación de las poblaciones estudiadas de P. devoniana en al menos dos variedades: P. devoniana var. devoniana y P. devoniana var. cornuta.
Palabras clave: Análisis de conglomerados, componentes principales, Pinus devoniana Lindl., Pinus michoacana Martínez var. cornuta Martínez, variación altitudinal, variación morfológica.
ABSTRACT
Understanding the patterns of morphological variation along environmental gradients is useful to distinguish taxonomical entities and take management decisions. Morphological variation of cones, seeds, fascicles and buds were analyzed on individuals from 16 natural Pinus devoniana (ex P. michoacana) populations collected along an altitudinal transect (from 1,600 to 2,450 m, one population every 50 m of altitudinal difference) near Morelia city, in the state of Michoacán (Mexico). Univariate (ANOVA) and multivariate (principal components and cluster) statistical analyses were conducted, in order to clarify if putative varieties exist. There is a significant pattern of morphological altitudinal variation, where populations from lower altitudes have larger cones, needles and seeds than populations from middle and higher altitudes. The multivariate analysis discriminated individuals and populations in two clearly distinctive groups: one group made of individuals from populations originated between 1,600 m to 1,850 m of altitude, which would correspond to Pinus devoniana var. devoniana (equivalent to P. michoacana typical); a second group is composed of individuals from populations originated between 1,950 m to 2,450 m, which would correspond to Pinus devoniana var. cornuta (ex P. michoacana var. cornuta). Thus, we suggest the separation of P. devoniana in at least two varieties: P. devoniana var. devoniana and P. devoniana var. cornuta.
Key words: Altitudinal variation, cluster analysis, morphological variation, Pinus devoniana Lindl., Pinus michoacana Martínez var. cornuta Martínez, principal components.
INTRODUCCIÓN
Pinus devoniana Lindl. es un pino que se distribuye, preferentemente, en el territorio mexicano: Colima, Chiapas, Guanajuato, Guerrero, Hidalgo, Jalisco, Michoacán, Estado de México, Morelos, Nayarit, Oaxaca, Puebla, Tlaxcala, Veracruz y Zacatecas; aunque su área de distribución se extiende a los departamentos de Chimaltenango, Guatemala, Quiche, Quetzaltenango, Sacatepequez y Totonicapán, en Guatemala. Habita en zonas de clima templado cálido, con una precipitación anual de 1,000 a 1,500 mm; en altitudes de 1,500 a 2,500 m (con crecimiento óptimo a 2,000 msnm); en bosques de pino-encino y de pino. Pinus devoniana tiene una etapa cespitosa, cuando es una plántula (Perry, 1991; Farjon y Styles, 1997).
La taxonomía de P. devoniana, descrita por primera vez por Lindley (1839), es un asunto no consensuado entre la comunidad científica. Shaw (1909) integró a Pinus devoniana con P. montezumae Lamb., P. hartwegii Lindl. y P. rudis Endl. en una sola especie incluyente: P. montezumae. Este agrupamiento fue dividido, nuevamente, en las cuatro especies originales por Martínez (1944), pero reconoce a P. devoniana bajo el nuevo nombre: Pinus michoacana Martínez, con una subdivisión en dos variedades: var. cornuta y var. quevedoi y tres formas: var. cornuta f. nayaritana, f. tumida y f. procera (Martínez, 1948). Excepto, por la forma tumida, en general, esta clasificación es aceptada por Perry (1991).
Pinus michoacana típico (sensu Martínez, 1948) tiene conos menos curveados, más grandes, oblongo-ovoides o casi cilíndricos y cónicos en el extremo, con vainas fasciculares de color más claro, apófisis más elevadas y ampliamente piramidales en comparación con la variedad cornuta, la cual presenta conos más curvos (semejantes a cuernos), más cortos, menos cilíndricos, gradualmente atenuados, más obscuros y resinosos (casi negros con la edad), con vainas fasciculares y la apófisis aplanada con dos lados (Martínez, 1948; Perry, 1991). Perry (1991) también apoya la diferenciación de la variedad cornuta basándose en las diferencias del contenido de trementina α-pineno y β -pineno.
Sin embargo, las variedades de P. michoacana no han sido reconocidas en clasificaciones posteriores. Rushforth (1987) notó que el nombre anterior y el corregido de P. michoacana correspondían, de hecho, a P. devoniana. McVaugh (1992) y Farjon y Styles (1997) favorecieron esta denominación, como el nombre correcto y concluyeron que no había argumentos para mantener la subdivisión en variedades y formas a partir de los rasgos usados por Martínez (1948), que no siempre fueron consistentes al examinar varios ejemplares de herbario (muchos con caracteres intermedios). Con respecto a la variedad cornuta, McVaugh (1992) indicó que el contenido de trementina no parece ser suficiente para sustentar su diferenciación como una variedad.
El propósito del presente estudio fue explorar la existencia de elementos para reconocer la variedad cornuta, al compararla con la típica P. devoniana con base en un análisis cuantitativo de la variación morfológica entre las poblaciones a lo largo de un gradiente altitudinal en el estado de Michoacán, entidad ubicada en el centro-occidente de México. Las características utilizadas fueron cono, semilla, fascículos y caracteres morfológicos de las yemas. Mediante este documento, se asume que el nombre correcto para P. michoacana es P. devoniana; entonces, P. devoniana var. devoniana se considera la especie típica y P. devoniana var. cornuta como la variedad putativa.
MATERIALES Y MÉTODOS
Muestreo y evaluación morfológica
En un transecto altitudinal, se recolectaron conos y ramillas de siete árboles procedentes de cada una de las 18 poblaciones naturales de P. devoniana. Estas se localizaron con una diferencia de altitud de aproximadamente 50 m, en las cercanías del pueblo de Túmbisca (1,600 msnm, 19°34'40.0'' N y 101°03'55.2'' O) y de San Miguel del Monte (2,450 msnm, 19°35'52.1'' N y 101°07'55.5''O). Los individuos se eligieron al azar entre los que portaban conos maduros, sin diferenciar entre variedades putativas. Los lugares de muestreo a los 1,900 y 2,000 m de altitud se descartaron del análisis, porque P. devoniana tenía muy baja densidad (únicamente dos árboles presentaban conos en cada una de las poblaciones). Los ejemplares representados por las muestras se designaron como poblaciones; mientras que, a la localidad se le denominó procedencia.
Se eligió una rama de los árboles con al menos un cono desarrollado, sin daños por insectos o enfermedades, y con el tamaño promedio de los conos maduros existentes en cada individuo muestreado. Si había más de un cono con dichos criterios, se eligió uno al azar. Antes de abrir, se midió la longitud del mismo, de la base a la punta mediante un calibrador Mitutoyo modelo 700-127 modificado para medir la longitud de un vector base-punta; ancho, a la mitad del cono; curvatura, que se obtuvo con el mismo calibrador y consistió en la distancia de la parte central del cono a su vector base-punta (expresado en porcentaje de la longitud); y forma: 1 = oblongo-ovoide, 2 = intermedio, y 3 = gradualmente atenuado. También se marcó una escama en la parte lateral del cono, justo a la mitad de su longitud. Después de la apertura de los conos, se midió la anchura de la quilla en la escama marcada, así como la altura de la apófisis y se le contó el número de caras. Se extrajeron las semillas del cono y se midieron las que estaban totalmente desarrolladas, con los mismos criterios usados para los conos. A las semillas y las alas se les determinó el largo y ancho y se registró su color: 1 = café suave, 2 = café medio, 3 = café rojizo y 4 = café obscuro. Se seleccionó un fascículo totalmente desarrollado por rama de árbol, sin señales de declinación y se contabilizaron sus acículas, longitud de la acícula más larga, y se definió el color de la vaina: 1 = café, 2 = café negruzco y 3 = negro. Se midió la longitud, el ancho y la forma del catáfilo: 1 = acuminado y 2 = rostrado (Stearn, 1996).
Los individuos se clasificaron como P. devoniana var. devoniana y la variedad putativa cornuta con una clave botánica (Madrigal, 1982, basado en Martínez, 1948). Algunos ejemplares presentaron caracteres ubicados entre ambos taxa, por lo que se clasificaron como intermedios. Por último, se estimó la proporción de individuos por categoría y población.
Análisis Estadísticos
Se hizo el Análisis de Varianza (ANOVA) para cada carácter, a fin de probar la significancia entre poblaciones mediante el programa PROC GLM (SAS, 2004), y los componentes de la varianza fueron estimados con el programa PROC VARCOMP METHOD = REML (SAS, 2004), con el siguiente modelo:
Donde:

Yij = Observación en el Jésimo árbol de la iésima población
µ = Media
τi = Efecto de la iésima población
εij = término del error
l = 1 = 1,...p, j= 1,...n; donde p y n corresponden al número de poblaciones y árboles dentro de las poblaciones.
Se hizo un análisis múltiple de medias por cada variable con la prueba de Tukey (α=0.05). Los caracteres morfológicos que eran conteos o categorías: forma del cono, número de caras de la apófisis y número de acículas, color de la vaina del fascículo, forma del catáfilo y color del ala de la semilla se analizaron por medio de la prueba no paramétrica de Kruskal-Wallis. La relación de la variación entre poblaciones y la altitud de la procedencia solo se evaluó para aquellos caracteres con variación significativa entre poblaciones, para ello se corrió un análisis de regresión del promedio de la población, con respecto a la altitud de la procedencia mediante el programa PROC REG (SAS, 2004) y el modelo:
Donde:

Yij = Media de la población
β0 = Intercepto
βi = Parámetro de regresión (pendiente)
Xi= Altitud (m) de la iésima procedencia
εij= Error.
Así mismo, se llevó a cabo un análisis multivariado por Componentes Principales (PROC PRINCOMP) (SAS, 2004) y un análisis de conglomerados (PROC CLUSTER) en el que se incluyeron únicamente las variables que tuvieron diferencias significativas entre poblaciones en el análisis univariado y se excluyeron las altamente correlacionadas con otras (R2>0.70). Del análisis de Componentes Principales, la diferenciación entre poblaciones fue probada con un ANOVA y una prueba múltiple de medias de los componentes principales relevantes, con el modelo (1).
El patrón de diferenciación entre las poblaciones a lo largo del gradiente altitudinal se determinó a partir de la comparación de los valores individuales de componentes principales con la procedencia. El análisis de conglomerados se corrió con datos estandarizados, para lo cual se usó un método de vinculación promedio (método de grupos pareados no ponderados con media aritmética (UPGMA)) y se generó un diagrama de árbol por medio del programa PROC TREE (SAS, 2004).
Finalmente, se compararon los resultados del análisis cuantitativo con la clasificación tentativa inicial en dos taxa putativos, P. devoniana var. devoniana y la putativa P. devoniana var. cornuta, con base en los criterios explicados por Martínez (1948).
RESULTADOS Y DISCUSIÓN
Distribución altitudinal de variedades basada en la descripción de Martínez (1948).
La proporción de individuos por población perteneciente a P. devoniana var. devoniana y la categoría denominada intermedia (Martínez, 1948) reveló una distribución altitudinal de las variedades putativas (Figura 1). Las poblaciones presentes entre 1,600 y 1,700 msnm pertenecen a P. devoniana var. devoniana, y las del intervalo de 1,950 a 2,450 msnm corresponden a la putativa P. devoniana var. cornuta. Las restantes se disponen en altitudes que varían de 1,750 a 1,850 m, que parece ser una zona de transición donde las poblaciones están compuestas por una mezcla gradual de las variedades putativas, además de individuos clasificados como intermedios (Figura 1).

Los taxa putativos corresponden a una clasificación tentativa inicial basada en los criterios de Martínez (1948)
Análisis univariado: análisis de varianza entre poblaciones y regresión respecto a la altitud
Se consideraron 20 caracteres, y de ellos 16 mostraron diferencias significativas entre poblaciones. El ancho y la forma del catáfilo, número de acículas por fascículo y el color de las alas no fueron significativamente diferentes entre poblaciones (Cuadro 1). Las gráficas de los valores poblacionales promedio respecto a la altitud indicaron, en general, una tendencia altitudinal, en la cual las poblaciones de localidades bajas tuvieron valores promedio más altos, que en las de zonas elevadas. Esto significa, que las primeras tienen árboles, conos, acículas y pedúnculos más grandes; apófisis más prominentes, quillas más amplias, mayor número de caras en las apófisis (Figura 2); así como catáfilos más largos, las semillas más grandes y anchas y aladas. Tendencia similar a la que se observa en otras poblaciones de coníferas; por ejemplo, Pinus oaxacana Mirov, de altitudes bajas (en Chiapas) presenta acículas de mayor tamaño que las especies de altitudes superiores (Bermejo y Patiño, 1981).

Otros caracteres, igualmente, mostraron una tendencia clinal, pero en las poblaciones de elevaciones bajas tuvieron valores promedio inferiores: conos menos curvos y atenuados, vainas fasciculares más cortas y de tonos menos obscuros (Figura 3).
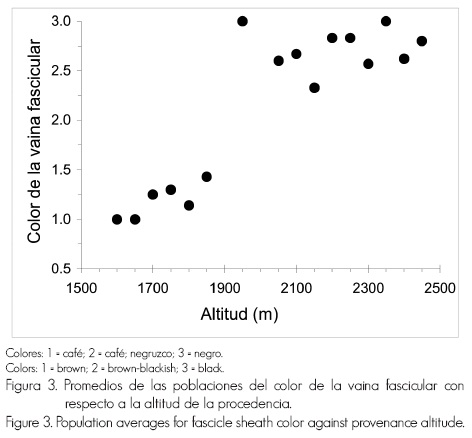
La asociación entre los promedios de la población y la altitud de las procedencias fue significativa en todas las variables examinadas (aquéllas que tenían diferencias significativas entre las poblaciones), después del análisis de regresión con un modelo lineal (Cuadro 1).
Análisis Multivariado: componentes principales y análisis de conglomerados
Cuatro componentes principales (PC) registraron Eigenvalores por encima de 1 (7.19, 1.39, 1.20 y 1.09). El componente principal 1 (PC1) explicó 45.0 % de la varianza. Los componentes PC2, PC3 y PC4 contribuyeron mucho menos a explicar la varianza total: 8.7 %, 7.5 % y 6.8 %, respectivamente. Por esta razón, la atención se centró en PC1.
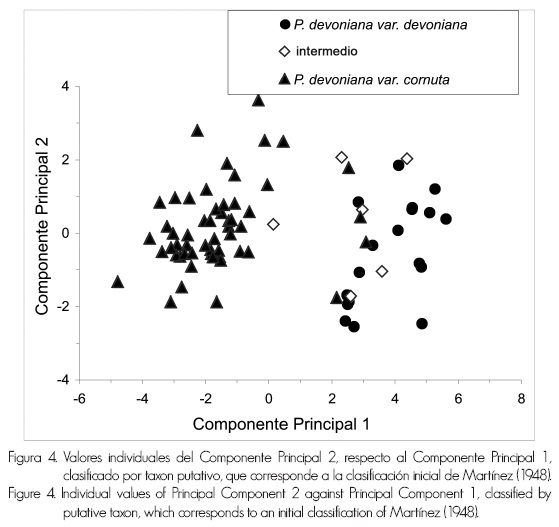
El conjunto de los valores individuales de PC2, en contraste con los de PC1 (Figura 4) indicó una separación de individuos en dos grupos: uno (lado derecho de la Figura 4) que incluye a la mayoría de los individuos inicialmente identificados como P. devoniana var. devoniana, a los intermedios (excepto uno) y a cuatro individuos (procedentes de los 1,850 msnm) pertenecientes a la variedad cornuta; un segundo grupo (lado izquierdo de la Figura 4) en el que se integran los ejemplares de P. devoniana var. cornuta y un individuo de los intermedios.
Los valores individuales del componente principal 1 (PC1) separan dos grupos altitudinales, con un patrón más ecotípico o discontinuo que clinal, el cual corresponde a dos taxa putativos: uno (lado superior izquierdo de la Figura 5) integrado por individuos cuyos valores de PC1, son superiores a 0, y que corresponden a poblaciones distribuidas entre 1,600 y 1,850 msnm; el segundo (lado inferior derecho de la Figura 5) está compuesto por árboles con valores de PC1, en su mayoría, por debajo de 0 con poblaciones presentes en un intervalo altitudinal de 1,950 a 2,450 m. Estos conjuntos coinciden con la separación en dos variedades: el primer grupo corresponde a la variedad devoniana e incluye a los individuos de la zona de transición, también denominados intermedios y a cuatro árboles (a partir de 1,850 m de altitud), tentativamente clasificados como variedad cornuta; y otro grupo identificado como P. devoniana var. cornuta.

El ANOVA de los valores individuales del PC1 señalaron diferencias significativas entre las poblaciones (P≤0.001), y la prueba de Tukey confirmó la separación en dos grupos poblacionales ecotípicos distintos: uno distribuido entre 1,600 y 1,850 msnm, que corresponde a P. devoniana var. devoniana (grupo "a" en la Figura 6), y otro integrado por poblaciones que se ubican entre 1,950 y 2,450 msnm, variedad cornuta (grupo "b" en la Figura 6).

Árboles de la zona de transición (1,750 y 1,850 msnm, identificados de manera tentativa como intermedios y cuatro como variedad cornuta) tuvieron caracteres de los valores promedio que no pueden ser estadísticamente diferenciados de las poblaciones pertenecientes a P. devoniana var. devoniana. Entonces, para propósitos prácticos, las poblaciones de la zona de transición se consideran parte de la variedad devoniana.
Una separación de poblaciones en dos grupos, claramente distintivos, fue confirmada por el análisis de conglomerados, en el cual las distribuidas en altitudes de 1,600 y 1,850 m (lado izquierdo de la Figura 7) se agregan; mientras que, las presentes en el intervalo altitudinal de 1,950 a 2,450 m constituyen un conglomerado aparte (lado derecho de la Figura 7). La separación entre ambos ocurre en el valor de 1.30.
Cuando los caracteres se analizan en conjunto con un análisis multivariado, se observa una marcada distinción entre los dos grupos de poblaciones, en la que parece existir un umbral en la diferenciación morfológica a 1,900 msnm; que contrasta con el patrón continuo de variación genética clinal entre poblaciones, para caracteres morfológicos individuales y para daños por heladas en las mismas poblaciones (Sáenz-Romero y Tapia-Olivares, 2008). Esta diferencia evidencia la actuación diferencial de diversas fuerzas de presión de selección sobre varios caracteres. Las temperaturas ambientales, igualmente, siguen una tendencia clinal que equipara el patrón genético para daño por heladas. Sin embargo, las fuerzas de evolución involucradas en los patrones de variación morfológica parecer tener una dinámica diferente, que dieron origen a una discontinuidad alrededor de 1,900 m de altitud.
CONCLUSIONES
La variación entre poblaciones sugiere una tendencia altitudinal, en la que las poblaciones de altitudes bajas (de 1,600 a 1,850 msnm) tienen conos, acículas y semillas más grandes, y, aparentemente, pertenecen a P. devoniana var. devoniana (variedad típica); mientras que, las poblaciones de altitudes intermedias y altas (de 1,950 a 2,450 m) presentan conos, acículas y semillas más pequeños, y por lo tanto, corresponden a P. devoniana var. cornuta.
Estos resultados tienen implicaciones más amplias, en particular en la taxonomía de las especies, ya que apoya la propuesta original de Martínez que separa la especie típica P. michoacana y P. michoacana var. cornuta. Dado que el nombre correcto es P. devoniana, se sugiere que se usen las denominaciones P. devoniana var. devoniana y P. devoniana var. cornuta. Sin embargo, estos resultados son preliminares, pues se requiere de un mayor análisis de la variación morfológica en otras localidades dentro de la distribución natural de la especie.
AGRADECIMIENTOS
El financiamiento fue proporcionado por CSR con recursos del Fondo Mixto de Investigación Forestal, CONACyT- CONAFOR (proyecto 2002-C01-4655) y por la Coordinación de la Investigación Científica, Universidad Michoacana de San Nicolás de Hidalgo.
REFERENCIAS
Bermejo V., B. y F. Patiño V. 1981. Estudio de variación de las características morfológicas de Pinus pseudostrobus Lindl. y Pinus pseudostrobus var. oaxacana Martínez en Chiapas. Revista Chapingo (29-30):3-8. [ Links ]
Farjon, A. and B. T. Styles. 1997. Pinus (Pinaceae). Flora Neotropica Monograph 75. New York Botanical Garden, NY, USA. 291 p. [ Links ]
Lindley, J. 1839. Miscellaneous notices: Mexican pines. Edwards's Bot. Reg. 25:62-64. [ Links ]
Madrigal S., X. 1982. Claves para la identificación de las coníferas silvestres del Estado de Michoacán. Boletín Divulgativo No. 58. Instituto Nacional de Investigaciones Forestales. México, D.F., México. 100 p. [ Links ]
Martínez, M. 1944. Una nueva especie del género Pinus. Pinus michoacana. Anales del Instituto de Biología 15(1):1-6. [ Links ]
Martínez, M. 1948. Los pinos mexicanos. Editorial Botas. México, D.F., México. 361 p. [ Links ]
McVaugh, R. 1992. Flora Novo-Galiciana. Gymnosperms and Pteridophytes. University of Michigan Press, Ann Arbor, MI. USA. Vol. 17, 467 p. [ Links ]
Perry, J. P. 1991. The Pines of México and Central America. Timber Press. Portland, OR USA. 231 p. [ Links ]
Rushforth, K. D. 1987. Conifers. Facts on File Publications. Christopher Helm, Oxford, UK. 432 p. [ Links ]
Sáenz-Romero, C. and B. L. Tapia-Olivares. 2008. Genetic variation in frost damage and seed zone delineation within an altitudinal transect of Pinus devoniana (P. michoacana) in Mexico. Silvae Genetica 57(3):165-170. [ Links ]
Statistical Analysis System (SAS). 2004. SAS Institute Inc. SAS/STAT User's Guide. Release 9.1. SAS Institute. Raleigh, NC USA. 5136 p [ Links ]
Shaw, G. R. 1909. The Pines of Mexico. Arnold Arboretum No.1 J. R. Ruiter & Co., Boston, MA USA. 29 p. [ Links ]
Stearn, W. T. 1996. Botanical Latin. David & Charles. Newton Abbot, Devon, UK. 546 p. [ Links ]

