










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista mexicana de ciencias forestales
versão impressa ISSN 2007-1132
Rev. mex. de cienc. forestales vol.3 no.10 México Mar./Abr. 2012
Artículo
Emergencia de plántulas de especies maderables nativas de la Península de Yucatán
Emergence of seedlings of native timber trees of Yucatan Peninsula
Benito Bernardo Dzib-Castillo1, Hans van der Wal 2, Cristina Isabel Chanatásig-Vaca1, Pedro Antonio Macario Mendoza3 y Juan Manuel Pat Fernández1
1 El Colegio de la Frontera Sur, Campeche. Correo-e: benitodzib@hotmail.com
2 El Colegio de la Frontera Sur, Tabasco
3 El Colegio de la Frontera Sur, Quintana Roo
Recibido el 13 de enero de 2011
Aceptado el 26 de marzo de 2012.
Resumen
La biodiversidad regional en la Península de Yucatán registra especies arbóreas nativas con potencial para ser usadas en la reforestación; sin embargo, su aprovechamiento requiere el estudio de sus características ecológicas, de ahí que la presente investigación se enfoque en la emergencia de plántulas de taxa maderables procedentes de dicha zona en vivero y bajo diferentes grados de sombra (0, 35, 60 y 90%); así como a la emergencia y supervivencia a partir de la siembra directa de semillas en lomeríos de la misma región. Para ello, se realizó la evaluación del cedro (Cedrela odorata), chakté (Caesalpinia mollis), ciricote (Cordia dodecandra), ja'abín (Piscidia piscipula), tzalam (Lysiloma latisiliquum) y granadillo (Platymiscium yucatanum). En vivero, C. mollis mostró diferencias significativas (p = 0.006), ya que obtuvo la mayor emergencia (53 y 49%) en los tratamientos de 0 y 35% de sombra; mientras que, en campo la consiguió en la posición alta de los lomeríos (p = 0. 015, 1.9%); en contraste con P . piscipula (p = 0.005) que se originó en la baja (14.3%). Por otra parte, altos niveles de sombra (60 y 90%) redujeron los días necesarios para alcanzar 95% de plántulas emergidas en C. odorata, la cual presentó una supervivencia más alta (73.5%) en la posición elevada (p = 0.001) de los lomeríos, a diferencia de C. dodecandra (84.2%) que lo hizo en la inferior (p = 0.02). Los resultados expresan que el porcentaje de sombra influye poco en el tiempo de emergencia total de las plántulas y que los taxa estudiados difieren en sus preferencias por las posiciones en el paisaje.
Palabras clave: Caesalpinia mollis (Kunth) Spreng., Cedrela odorata L., Cordia dodecandra A. DC., Lysiloma latisiliquum (L.) Benth., Piscidia piscipula (L.) Sarg., Platymiscium yucatanum Standley.
Abstract
The regional biodiversity of Yucatan Peninsula includes native tree species with potential to be used en reforestation; however, to use this potential, it is necessary to know the ecological preferences of the species. Therefore, the emergence of se edlings of native timber tre es were assessed under different levels of shade (0, 35, 60 and 90%) at the nursery, and seedling emergence and survival after direct sowing of seeds in the lower and higher parts of the hills that are common in the Pensinsula, as well as the following species: cedar (Cedrela odorata), chakté (Caesalpinia mollis), ciricote (Cordia dodecandra), ja'abín (Piscidia piscipula), tzalam (Lysiloma latisiliquum) and granadillo (Platymiscium yucatanum). At the nursery, high levels of shade (60% and 90%) reduced the number of days required to attain 95% of the number of emerged seedlings in C.odorata. C. mollis was the only species that showed significant differences (p = 0.006) in the emergence of seedlings among shade treatments, obtaining the highest emergence per cent (53 and 49%) at 0% and 35% of shade. In the field, P. piscipula (p = 0.005) showed higher emergence in the lower part of the hills, and C. mollis (p = 0.015) in the higher part (1.9%). C. odorata showed higher survival (73.5%) in the high position (p = 0.001) and C. dodecandra (84.2%) in the low position (p = 0.02). The aforementioned results show that species differ in preferences for positions in the landscape, and that shade per cent has little effect on seedling emergence.
Key words: Caesalpinia mollis (Kunth) Spreng., Cedrela odorata L., Cordia dodecandra A. DC., Lysiloma latisiliquum (L.) Benth., Piscidia piscipula (L.) Sarg., Platymiscium yucatanum Standley.
Introducción
La deforestación y la degradación del bosque disminuyen la cantidad y la calidad de los servicios ambientales de conservación de la biodiversidad (Fahrig, 2003), del balance hidrológico (Croke et al., 2004) y de la captura de carbono (Bawa y Markham, 1995). Así mismo, afectan la provisión de productos forestales maderables y no maderables que satisfacen las necesidades primarias y constituyen medios de subsistencia para la población local de los países en desarrollo (Buschbacher, 1990; Lamb et al., 2005).
En la Península de Yucatán, las actividades productivas regionales (agricultura, ganadería y extracción selectiva de madera) han afectado la extensión, estructura y composición de las comunidades forestales en diversos grados (Escamilla et al., 2000; Martínez y Galindo-Leal, 2002). En Campeche, así como en el sureste de México, la deforestación ha sido consecuencia de esquemas de colonización auspiciadas por el gobierno (Remmers y Koeijer, 1992; Turner II et et al., 2001), situación que ha propiciado la disminución de las poblaciones de especies maderables preciosas como cedro (Cedrela odorata L.) y caoba (Swietenia macrophylla King.) y de otras como ciricote (Cordia dodecandra A. DC.), granadillo (Platymiscium yucatanum Standley.), chakté (Caesalpinia mollis (Kunth) Spreng.) y pucté (Bucida buceras L.).
Como resultado de ello se ha originado la activación de programas gubernamentales para recuperar o incrementar las poblaciones de las primeras dos especies, por su alto valor comercial, como son las plantaciones comerciales o de reforestación en bosque secundario realizadas en 103,795 ha entre 1993 y 2004 (CONAFOR, 2008). En México, las acciones para regenerar estas especies se han enfocado a incorporar plántulas bajo sombra en bosques recién aprovechados o alterados por corta selectiva; sin embargo, debido a la competencia por luz y nutrimentos con la vegetación asociada, así como a su manejo deficiente, es común que se caractericen por una supervivencia y un crecimiento reducidos. A nivel experimental se ha probado la siembra de semillas de S. macrophylla en claros dentro de bosques naturales, donde se observa que existe potencial en la emergencia de plántulas a partir de su siembra en campo (Gerhardt, 1993; Negreros-Castillo et al., 2003).
Lo antes mencionado apoya el uso de semillas de árboles y arbustos nativos de México, los cuales son potencialmente valiosos para ser utilizados en programas de reforestación y restauración ecológica (Vázquez-Yanes et al., 1999; Niembro, 2001). No obstante, para su aprovechamiento es necesario generar información sobre su fenología, morfología, estructura, latencia, viabilidad, germinación y desarrollo inicial de sus individuos jóvenes (Niembro, 2001), cuya emergencia y establecimiento en campo constituyen sus fases críticas en el ciclo de vida (Angevine y Chabot, 1979).
Un factor importante en el proceso de establecimiento es la luminosidad, pues influye en algunas semillas que necesitan luz para germinar (fotoblastismo positivo), a diferencia de otras que pueden hacerlo en la obscuridad (fotoblastismo negativo) (Grime et al., 1981; Vázquez-Yanes y Orozco-Segovia, 1987; Shinomura, 1997; Fenner y Thompson, 2005). Este efecto sobre la emergencia de plántulas de especies arbóreas tropicales del sureste mexicano se ha estudiado en S. macrophylla (Morris et al., 2000), Cecropia obtusifolia Bertol. y Piper aurantium Kunth (Vazquez-Yanes y Orozco-Segovia, 1987).
Por otro lado, la topografía es una fuente de heterogeneidad en los bosques tropicales (Daws et al., 2005), pues afecta la disponibilidad de agua para la planta (Whitmore and Burnham, 1984), la cantidad de hojarasca en el suelo (Becker et al., 1988); así como el pH y la capacidad de intercambio catiónico (Silver et al., 1994), lo que incide en la distribución de algunos taxa (Gibbons y Newbery, 2002). Para la Península de Yucatán se han definido relaciones entre la topografía y la distribución de tipos (Miranda, 1978) y asociaciones de vegetación (Escamilla et al., 2000); sin embargo, son escasos los datos publicados sobre las condiciones ecológicas más convenientes para las especies de mayor abundancia, situación que constituye una laguna en el conocimiento, dada la importancia de la variación edáfica y de humedad a lo largo de las toposecuencias en el ambiente kárstico de esa zona (Hernández, 1985; Duch, 1991).
En la presente investigación se evaluó la emergencia y supervivencia de las plántulas de seis especies maderables nativas de la Península de Yucatán, interesantes por sus aplicaciones locales y comerciales, mediante dos experimentos, uno en un vivero rústico a fin de conocer el efecto de diferentes niveles de sombra y el otro en campo, por siembra directa, para determinar el efecto de las laderas de lomeríos.
Materiales y Métodos
Área de estudio
Los experimentos se realizaron en el estado de Campeche, México: el relativo a la emergencia de plántulas bajo diferentes condiciones de sombra se llevó a cabo en un vivero acondicionado ex professo en un predio de la ciudad de Campeche; el de siembra directa en campo se desarrolló en lomeríos del Ejido de Pomuch, municipio Hecelchakán (20° 2' 50.9" norte y 90° 7' 23.58" oeste), el cual presenta una orografía poco accidentada y un terreno kárstico que se distingue por la alternancia de colinas abundantes que rodean depresiones planas, cuyos sedimentos aluviales arcillosos se inundan durante la época de lluvias debido al ineficiente drenaje de sus suelos (Marshall, 2007). En la transición entre lomeríos y depresiones, en la ladera inferior de los primeros, los suelos son más profundos que en las zonas elevadas donde es común que su carácter pedregoso se acentúe. Por esta razón y por motivo de la gravedad, la humedad es mayor en las partes bajas que en las altas. El clima de la región corresponde al cálido subhúmedo con lluvias en verano, temperatura media anual de 26 °C (INEGI, 2010a) y precipitación media anual de 1,000 a 1,100 mm (INEGI, 2010b).
Especies estudiadas
En campo se estudiaron el cedro (Cedrela odorata), chakté (Caesalpinia mollis), ciricote (Cordia dodecandra), ja'abín (Piscidia piscipula) y granadillo (Platymiscium yucatanum); mientras que, en el de vivero se probaron las mismas, con excepción de la última que fue reemplazada por tzalam (Lysiloma latisiliquum (L.) Benth.) como consecuencia de la falta de semillas.
Recolección de semillas
Se efectuaron recorridos en ejidos de los municipios Champotón, Calakmul, Hopelchén y Campeche, donde fueron seleccionados individuos fisiológicamente maduros de las especies arriba mencionadas. Con la ayuda de productores locales se dio seguimiento a tres individuos de cada una hasta la cosecha de sus frutos. Los relativos a Cedrela odorata y Cordia dodecandra se recolectaron en marzo, los de Caesalpinia mollis, Lysiloma latisiliquum y Platymiscium yucatanum en abril y mayo y los de Piscidia piscipula en junio. Las semillas se extrajeron manualmente y se secaron al sol sin tener una medida exacta de su humedad, por lo que se tomó como referencia su color y textura, se introdujeron en bolsas Ziploc® a 4 °C y se almacenaron durante tiempos distintos, según la época de fructificación y colecta: Cedrela odorata y Cordia dodecandra seis meses, Caesalpinia mollis, Lysiloma latisiliquum y Platymiscium yucatanum cinco meses y Piscidia piscipula alrededor de tres meses, para concluir en la fecha de inicio del experimento.
Emergencia de plántulas bajo diferentes grados de sombra en vivero
El diseño experimental correspondió a uno de bloques completos al azar con cuatro tratamientos y tres repeticiones. Las semillas se depositaron en bolsas de 8 x 15 cm, con suelo Luvisol como sustrato (conocido localmente como kankab) y se distribuyeron en cuatro camas. Para cada tratamiento se emplearon mallas de plástico de 35, 60 y 90% de sombra (Mallas textiles®), excepto para el de 0%. Las repeticiones consistieron en 100 bolsas con una semilla por repetición colocadas de forma aleatoria, a razón de tres por taxón a lo largo de cada cama. Su siembra, realizada el 14 de septiembre de 2007, tuvo una profundidad igual al diámetro más largo de la semilla de cada especie. Los datos de emergencia de las plántulas se tomaron diariamente por 36 días, después de los cuales ya no se observó incremento en este sentido.
Emergencia de plántulas en campo
La siembra directa de semilla de las cinco especies se realizó en puntos de siembra directa, con un metro de separación, en tres líneas paralelas a lo largo de dos brechas, de 100 m de longitud por 2 m de ancho, una en la parte superior (a) y la otra en la inferior (b) de la ladera de cinco lomeríos, en donde se sembraron de forma alternada de una a tres semillas: 180 de Caesalpinia mollis, 120 de Cedrela odorata y Piscidia piscipula, y 60 de Cordia dodecandra y Platymiscium yucatanum (Figura 1).

Figura 1. Representación esquemática de la localización de brechas abiertas en vegetación secundaria para la siembra directa de semillas de cinco especies arbóreas.
Figure 1. Scheme of the location of open paths of secondary vegetation for direct seed sowing of five tree species.
El número de plántulas emergidas se evaluó a los 45 días, del 25 al 28 de agosto de 2006, y la supervivencia, el 18 de febrero de 2007. En esta fase de siembra y emergencia-entre agosto y octubre la precipitación media fue de 234 mm (CONAGUA, 2012) (Figura 2).

Figura 2. Promedio de precipitación mensual en el año de siembra de las semillas en campo (CONAGUA, 2012).
Figure 2. Monthly average precipitation in the year of seed sowing in the field (CONAGUA, 2012).
Análisis estadísticos
Se examinó el tiempo requerido para alcanzar 95% del número de plántulas emergidas en cada especie y tratamiento de sombra y, con el fin de identificar las diferencias entre tratamientos se efectuaron ANOVAs (Análisis de varianza univariante) y la prueba de Tukey en los que se empleó el paquete estadístico INFOSTAT (INFOSTAT, 2008).
Para determinar el efecto de los tratamientos sobre l o s porcentajes de emergencia, los datos se transformaron a raíz cuadrada de arcoseno y se realizaron igualmente ANOVAs con la prueba de comparación de medias de Tukey. En el experimento en campo, para identificar diferencias tanto en la emergencia de plántulas como en la supervivencia en ambas zonas de los lomeríos, se aplicaron análisis binomiales en el programa SPSS (SPSS, 2003).
Resultados
Tiempo requerido para la emergencia de plántulas en vivero
El número de días para lograr 95% de emergencia varió de manera significativa entre tratamientos de dos especies: Cedrela odorata (p = 0.007) se obtuvo en menos días con 60% de sombra y Cordia dodecandra (p = 0.005) en el de 35% (Figura 3).

Figura 3. Número de días requeridos para que emerja 95% del número total final de plantas emergidas de cinco especies arbóreas bajo 0, 35, 60 y 90% de sombra.
Figure 3. Number of days necessary for the emergence of 95% of the total final number of plants emerged of five tree species under of 0, 35, 60 and 90% shadow.
Caesalpinia mollis fue la única que presentó diferencias importantes (p = 0.006) entre tratamientos, ya que tuvo la mayor emergencia con la luminosidad más elevada (0 y 35% de sombra) (Figura 4).
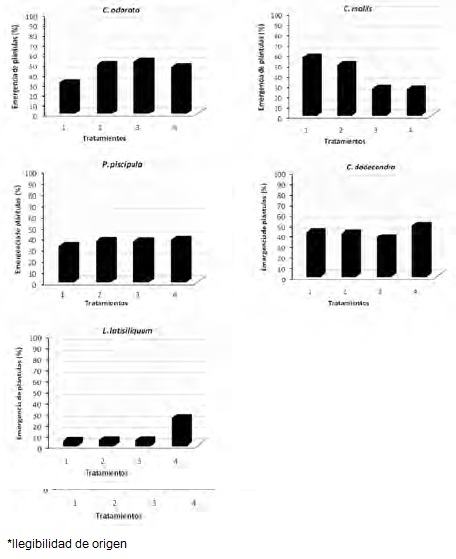
Figura 4. Porcentaje de emergencia de plántulas por especie en los diferentes tratamientos de sombra en vivero (1 = 0%, 2 = 35%, 3 = 60%, 4 = 90% de sombra).]
Figure 4. Seedling emergence by species in the different shadow treatments at the nursery (1 = 0%, 2 = 35%, 3 = 60%, 4 = 90% of shadow).
Emergencia y supervivencia de plántulas en campo
El análisis mostró diferencias estadísticas significativas en la emergencia de plántulas de Piscidia piscipula (p = 0.058) y Caesalpinia mollis (p = 0.015) en las posiciones de los lomeríos: para la primera, la mayor emergencia se presentó en la zona inferior (14%), respecto a la elevada (9.2%) y para la segunda, en la superior (1.9%) en relación a la baja (0.6%). Por otro lado, en Cedrela odorata y Cordia dodecandra se registró en la región alta; mientras que, Platymiscium yucatanum en la baja (Figura 5); aunque las diferencias no fueron estadísticamente significativas.

Figura 5. Emergencia de plántulas de especies maderables de la parte alta y baja de lomeríos de Campeche.
Figure 5. Seedling emergence of timber species in the high and low part of the hillocks of Campeche.
La fracción que sobrevivió en las dos zonas del lomerío fue distinta en algunas especies. En Cedrela odorata la supervivencia fue mayor en la posición superior (74%), ya que la inferior fue de 17% (p = 0.001), al contrario de Cordia dodecandra que se dio en la parte baja (84%), pues la alta fue de 56% (p = 0.020). En cuanto a Caesalpinia mollis y Piscidia piscipula, también se observó en la parte elevada y Platymiscium yucatanum en la inferior (Figura 6); no obstante, las diferencias no fueron estadísticamente significativas.

Figura 6. Supervivencia de plántulas en dos posiciones (alta y baja) en lomeríos de Campeche.
Figure 6. Plant survival at two heights (high and low) in hillocks of Campeche.
Discusión
Emergencia de plántulas en vivero
Las especies estudiadas muestran diferentes respuestas germinativas ante los gradientes de luz. En Caesalpinia mollis el porcentaje más grande de emergencia se logró en menor número de días con 90% de sombra, en contraste con Cordia dodecandra que con 35% de sombra y en un menor tiempo alcanzó 95% de producción. Por su parte, Piscidia piscipula obtuvo 50% de plántulas del total que emergieron en menos días en el tratamiento de 90%.
Lo anterior subraya que el sombreado a diferente intensidad disminuye el tiempo de emergencia de las plántulas de Cedrela odorata, Caesalpinia mollis, Piscidia piscipula y Cordia dodecandra, en particular, pues Lysiloma latisiliquum reaccionó indiferente a los grados de sombra. En consecuencia, se pueden tener plantas listas para el transplante en lapsos más cortos.
Al término del experimento, solo Caesalpinia mollis mostró diferencias estadísticas significativas en las que se tuvo la mayor emergencia en los tratamientos con más exposición solar (0 y 35% de sombra), lo cual indica un fotoblastismo positivo (Grime et al., 1981; Vázquez-Yanes y Orozco-Segovia, 1987; Shinomura, 1997; Fenner y Thompson, 2005), por lo que se desarrolla principalmente en selvas bajas y medianas subcaducifolias, donde la luz solar es abundante en el sotobosque durante los meses de sequía y al inicio de la temporada de lluvias (Vester y Navarro, 2007). Estas observaciones aportan criterios para la selección de sitios para la siembra de sus semillas, en contraste con las de las otras que pueden emerger en intervalos más amplios de luminosidad.
Lysiloma latisiliquum presentó la menor emergencia de plántulas, 9%, valor diferente al de aquellas citadas en la literatura que varían entre 40 y 50% (Cordero y Boshier, 2003). En esta especie, así como en otras de la familia Fabaceae, se han obtenido porcentajes más altos de emergencia después de utilizarse procesos de escarificación (Orozco-Almanza et al., 2003), razón que orienta al empleo de tratamientos pregerminativos para L. latisiliquum.
Emergencia y supervivencia de plántulas en campo
Caesalpinia mollis y Piscidia piscipula fueron las únicas especies que demostraron diferencias estadísticas significativas en la emergencia de plántulas respecto a la pendiente del lomerío, ya que la zona elevada resultó mejor para C. mollis; mientras que, la baja para P. piscipula.
Los porcentajes de emergencia después de la siembra directa de semillas en campo fueron bajos y variaron de 1.2% en Caesalpinia mollis a 17.2% en Cedrela odorata. Sin embargo, este método proporciona varios beneficios como el ahorro en mano de obra para el mantenimiento de plantas, la infraestructura en vivero y el traslado de planta. Así mismo, se elimina el estrés que de manera común causa una supervivencia baja posterior al transplante, como se ha observado en C. odorata en diferentes grados de sombra bajo dosel en enriquecimientos de vegetación secundaria (González-Rivas et al., 2009).
Las diferencias de estos porcentajes en relación a las posiciones de los lomeríos responden a diversos factores. Por una parte, es posible que la lluvia haya removido las semillas, debido a su ubicación en pendiente. No obstante, dado que estas se enterraron y que existe un escurrimiento superficial limitado en las condiciones de karst de la Península (García y Pat, 2000; Zimmerman et al., 2000; Guariguata y Ostertag, 2001; Sedovet al., 2008), se considera poco probable que esto haya ocurrido. De acuerdo con Janzen y Vázquez-Yanes (1991), otra causa pudiera ser la depredación de semillas por animales y el ataque por hongos. De hecho, ellos estiman que la mitad del germoplasma producido por más del 90% de las especies arbóreas tropicales muere antes de germinar.
En el vivero, bajo condiciones controladas de humedad, el porcentaje de emergencia fue mayor que el de campo, lo que sugiere que la falta de agua en este último intervino en dicha respuesta. En consecuencia, explicaciones alternativas se basan en la calidad del sustrato y en una baja exposición a depredadores. La disponibilidad de agua para la emergencia de plántulas es crucial en la Península de Yucatán a causa de que los patrones regionales de precipitación influyen de forma notable en la distribución a gran escala de las asociaciones vegetales (Martínez y Galindo-Leal, 2002). Por consiguiente, en un entorno heterogéneo, la germinación debe ocurrir en un tiempo limitado para que los individuos dispongan de condiciones favorables de luz, agua y nutrimentos (Rincón et al., 1999; Khurana y Singh, 2001).
Los medios propicios para la supervivencia están relacionados con la humedad de las posiciones altas (Cedrela odorata) y bajas (Cordia dodecandra) de los lomeríos; así como con la necesidad de nutrimentos y su disponibilidad en ellas, debido a la variación de los componentes existentes en el suelo (Martínez y Galindo-Leal, 2002). Por ende, el estudio de los elementos que participan en la regeneración de plántulas es esencial para entender la dinámica en selvas tropicales caducifolias y así identificar los principales problemas sobre su restauración y conservación (Ceccon et al., 2006).
Conclusiones
El uso de sombra en vivero disminuye el tiempo de emergencia en Cedrela odorata, Caesalpinia mollis, Piscidia piscipula y Cordia dodecandra; sin embargo, C. mollis fue la única especie que reveló diferencias estadísticas significativas en la emergencia total y registró la mayor emergencia en condiciones de alta luminosidad.
Las especies que presentaron diferencias estadísticas significativas en la emergencia en lomeríos fueron C. mollis, con la mayor emergencia en la parte alta y P. piscipula, en la baja.
Cedrela odorata y Cordia dodecandra mostraron la mayor supervivencia de plántulas en lomeríos, la primera en la posición elevada y la segunda en la inferior.
Aún es necesario realizar más estudios sobre los factores que intervienen en la emergencia y supervivencia de plántulas en campo
Agradecimientos
Al Dr. Juan Carlos Pérez Jiménez por sus asesorías sobre los análisis estadísticos de los datos. Al Instituto Tecnológico de Chiná, Campeche, México y a sus estudiantes por su apoyo en la realización de este trabajo. Al Consejo Nacional de Ciencia y Tecnología (CONACYT) por el financiamiento del proyecto "Uso sustentable de los recursos naturales en la Frontera Sur" (clave SEMARNAT-2002-C01-1109), que permitió realizar el trabajo de campo, así como por la beca proporcionada para los estudios de doctorado del autor principal.
Referencias
Angevine, M. V. and B. F. Chabot. 1979. Seed germination syndromes in higher plants. In: Solbrig, O., S. Jain, G. Johnson and P. Raven (eds.). Topics in plant population biology. Columbia University Press, New York, NY. USA. pp.188-206. [ Links ]
Bawa, K. and A. Markham, 1995. Climate change and tropical forests. Trends in Ecology and Evolution 10:348–349. [ Links ]
Becker, P., P. E. Rabenold, J. R. Idol and A. P. Smith. 1988. Water potential gradients for gaps and slopes in a Panamanian tropical moist forest. Journal of Tropical Ecology. 4:173–184. [ Links ]
Buschbacher, R. 1990. Natural forest management in the humid tropics: ecological, social, and economic considerations. AMBIO 19(5): 253–258. [ Links ]
Ceccon, E., P. Huante and E. Rincón. 2006. Abiotic Factors influencing tropical dry forests regeneration. Brazilian Archives of Biology and Technology. 49 (2):305-312. [ Links ]
Comisión Nacional Forestal (CONAFOR). 2008. Sistema Nacional de Información Forestal. Compendio de estadísticas ambientales. 2006. http://148.223.105.188:2222/snif_portal/index.php?option=com_content&task=view&id=51&Itemid=65 (12 de marzo de 2008). [ Links ]
Comisión Nacional del Agua (CONAGUA). 2012. Climatología: temperatura y precipitación. http://smn.conagua.gob.mx/index.php?option=com_content&view=article&id=12&Itemid=77 (6 de febrero de 2012). [ Links ]
Cordero, J. y D. H. Boshier. 2003. Árboles de Centroamérica: un manual para extensionistas. OFI-CATIE, Turrialba, Costa Rica. 1079 p. [ Links ]
Croke, B. F. W., W. S. Merritt and A. J. Jakeman. 2004. A dynamic model for predicting hydrologic response to land cover changes in gauged and ungauged catchments. Journal of Hydrology. 291(1-2):115–131. [ Links ]
Daws, M. I., T. R. Pearson, D. F. Burslem, C. E. Mullins and J. W. Dalling. 2005. Effects of topographic position, leaf litter and seed size on seedling demography in a semi-deciduous tropical forest in Panama. Plant Ecology 179 (1):93–105. [ Links ]
Duch G., J. 1991. Fisiografía del estado de Yucatán, México. Universidad Autónoma Chapingo. Chapingo, Edo. de Méx. México. 229 p. [ Links ]
Escamilla, A., M. Sanvicente, M. Sosa and C. Galindo-Leal. 2000. Habitat mosaic, wildlife availability, and hunting in the Tropical Forest of Calakmul, México: Habitat disturbance and tropical rainforest mammals. Conservation Biology 14 (6):1592-1601. [ Links ]
Fahrig, L. 2003. Effects of habitat fragmentation on biodiversity. Annual Review of Ecology, Evolution and Systematics 34:487–515. [ Links ]
Fenner, M. and K. Thompson. 2005. The ecology of seeds. Cambridge University Press. Cambridge, UK. 260 p. [ Links ]
García G., G. y F. J. M. Pat. 2000. Apropiación del espacio y colonización en la reserva de la biosfera Calakmul, Campeche, México. Revista Mexicana del Caribe. 10:212-231. [ Links ]
Gerhardt, K. 1993. Tree seedling development in tropical dry forest abandoned pasture and secondary forest in Costa Rica. Journal of Vegetation Science. 4 (1):95-102. [ Links ]
Gibbons, J. M. and D. M. Newbery. 2002. Drought avoidance and the effect of local topography on trees in the understorey of Bornean lowland rain forest. Plant Ecology 164 (1):1-18. [ Links ]
González-Rivas, B., M. Tigabu, M. Castro-Marín, and P. C. Odén. 2009. Seed germination and seedling establishment of Neotropical dry forest species in response to temperature and light conditions. Journal of Forestry Research 20 (2):99-104. [ Links ]
Grime, J. P., G. Mason, A. V. Curtis, J. Rodman, S. R. Band, M. A. G. Mowforth, A. M. Neal and S. Shaw. 1981. A comparative study of germination in a local flora. Journal of Ecology 69:1017-1059. [ Links ]
Guariguata, M. R. and R. Ostertag. 2001. Neotropical secondary forest succession: changes in structural and functional characteristics. Forest Ecology and Management 148 (1-3):185-206. [ Links ]
Hernández X., E. 1985. La agricultura en la Península de Yucatán. Xolocotzia. Tomo 1. Universidad Autónoma Chapingo. Chapingo, Edo. de Méx. México. 799 p. [ Links ]
Instituto Nacional de Estadística y Geografía (INEGI). 2010a. Mapa de Temperatura Media Anual. http://mapserver.inegi.gob.mx/geografia/espanol/estados/camp/temperat.cfm. (15 de abril de 2010). [ Links ]
Instituto Nacional de Estadística y Geografía (INEGI) 2010b. Mapa de Precipitación Promedio Anual http://mapserver.inegi.gob.mx/geografia/espanol/estados/camp/precipit.cfm. (15 de abril de 2010). [ Links ]
INFOSTAT. 2008. Manual de usuario. (Versión 2008). Editorial Brujas. Córdova, Argentina. 336 p. [ Links ]
Janzen, D. H. and C. Vásquez-Yanes. 1991. Aspects of tropical seed ecology of relevance to management of tropical forested wild lands. In: Gomez-Pompa A., T. C. Whitmore and M. Hadley (Eds.). Rainforest regeneration and management. MAB Book Series 6. Carnforth, Lanes, UK. pp. 137-154. [ Links ]
Khurana, E. and J. S. Singh. 2001. Ecology of seed and seedling growth for conservation and restoration of tropical dry forest: a review. Conservation Biology 28 (1):39-52. [ Links ]
Lamb, D., P. D. Erskine, and J. A. Parrotta, 2005. Restoration of degraded tropical forest landscapes. Science 310(5754):1628-1632. [ Links ]
Marshall, J. S. 2007. The geomorphology and physiographic provinces on Central America. In: Bundschuh, J. and G. E. Alvarado (eds.) Central America: Geology, Resources and Hazards. Taylor & Francis. London, UK. pp. 1-51. [ Links ]
Martínez, E. y C. Galindo-Leal. 2002. La vegetación de Calakmul, Campeche, México: clasificación, descripción y distribución. Boletín de la Sociedad. Botánica de México 71:7-32. [ Links ]
Miranda, F. 1978. Vegetación de la Península Yucateca. Colegio de Postgraduados, SARH. Chapingo, Edo. de Méx. México. 271 p. [ Links ]
Morris M., H., P. Negreros-Castillo and C. Mize. 2000. Sowing date, shade, and irrigation affect big-leaf mahogany (Swietenia macrophylla King). Forest Ecology and Management 132 (2-3):173-181. [ Links ]
Negreros-Castillo, P., L. K. Snook, and C. Mize. 2003. Regenerating mahogany (Swietenia macrophylla) from seed in Quintana Roo, Mexico: the effects of sowing method and clearing treatment. Forest Ecology and Management 183 (1-3):351-362. [ Links ]
Niembro R., A. 2001. Las diásporas de los árboles y arbustos nativos de México: posibilidades y limitaciones de uso en programas de reforestación y desarrollo agroforestal. Madera y Bosques 7 (2): 3-11. [ Links ]
Orozco-Almanza, M., L. Ponce de León-García, R. Grether and E. García-Moya. 2003. Germination of four species of the genus Mimosa (Leguminosae) in a semi-arid zone of Central Mexico. Journal of Arid Environments 55 (1):75-92. [ Links ]
Remmers, G. and H. De Koeijer. 1992. The T'OLCHE', a Maya system of communally managed forest belts: the causes and consequences of its disappearance. Agroforestry Systems 18 (2):149-177. [ Links ]
Rincón, E., M. Álvarez A., G. González, D., P. Huante y A. Hernández, R. 1999. Restauración de selvas bajas caducifolias. Gaceta Ecológica. 53:62-71. [ Links ]
Sedov, S., E. Solleiro-Rebolledo, L. F. Scott, T. Pi-Puig, E. Vallejo-Gómez and M. Flores-Delgadillo. 2008. Micromorphology of a Soil Catena in Yucatán: Pedogenesis and Geomorphological Processes in a Tropical Karst Landscape. In: Kapur, S., A. Mermut and G. Stoops. (Eds.). New Trends in Soil Micromorphology. Springer Berlin Heidelberg., Germany. pp. 19-37. [ Links ]
Shinomura, T. 1997. Phytochrome regulation of seed germination. Journal of Plant Research. 110 (1):151-161. [ Links ]
Silver, W. L., F. N. Scatena, A. H. Johnson, T. G. Siccama and M. J. Sánchez. 1994. Nutrient availability in a montane wet tropical forest: spatial patterns and methodological considerations. Plant and Soil 164 (1):129-145. [ Links ]
Statistical Package for the Social Sciences (SPSS 12.0). 2003. User's Guide. USA. 703 p. [ Links ]
Turner II, B., S. Cortina V., D. Foster, J. Geoghegan, E. Keys, P. Klepeis, D. Lawrence, P. Macario M., S. Manson, Y. Ogneva-Himmelberger, A. B. Plotkin, D. Pérez S., R. Roy C., B. Savitsky, L. Schneider, B. Schmook and C. Vance. 2001. Deforestation in the southern Yucatán Peninsular region: an integrative approach. Forest Ecology and Management 154 (3):353-370. [ Links ]
Vázquez-Yanes, C., M. I. Alcocer S., M. Gual D. y C. Sánchez D. 1999. Árboles y arbustos potencialmente valiosos para la restauración ecológica y reforestación. Reporte técnico del proyecto J084. CONABIO-Instituto de Ecología. Universidad Nacional Autónoma de México. México, D. F., México. 15 p. [ Links ]
Vázquez-Yanes, C., A. I. Batis M., I. Alcocer-Silva, M. Gual D. and A. Orozco- Segovia. 1987. Light gap detection by the photoblastic seeds of Cecropia obtusifolia and Piper aurantium, two tropical rain forest trees. Biologia Plantarum (PRAHA) 29 (3): 234-236. [ Links ]
Vester H., F. M. y M. A. Navarro-Martínez. 2007. Fichas ecológicas: Árboles maderables de Quintana Roo. CONACYT-Quintana Roo, Ecosur. Chetumal, Q. Roo, México. 139 p. [ Links ]
Whitmore, T. C. and C. P. Burnham. 1984. Tropical rain forest of the far East. 2nd Ed. Clarendon Press. Oxford, UK. 1984 p. [ Links ]
Zimmerman, J. K., J. B., Pascarella and T. M. Aide. 2000. Barriers to forest regeneration in an abandoned pasture in Puerto Rico. Restoration Ecology 8 (4):350-360. [ Links ]

