










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de ciencias forestales
versión impresa ISSN 2007-1132
Rev. mex. de cienc. forestales vol.3 no.10 México mar./abr. 2012
Artículo
Influencia del fuego sobre la riqueza y diversidad de aves en un bosque templado en Puebla
The influence of fire upon bird and diversity in a temperate forest of Puebla State
Laura Patricia Ponce Calderón1, Beatriz Cecilia Aguilar Valdéz1, Dante Arturo Rodríguez Trejo1 , Elvia López Pérez1 y Javier Santillán Pérez1
1 División de Ciencias Forestales, Universidad Autónoma Chapingo. Correo-e: dantearturo@yahoo.com
Recibido el 5 de enero de 2011
Aceptado el 3 de marzo de 2012
Resumen
Desde el punto de vista ecológico, los incendios tienen efectos positivos sobre la vida silvestre y su hábitat en ecosistemas mantenidos por el fuego y bajo regímenes poco alterados. En México prácticamente no se ha estudiado su impacto en la avifauna. En el presente trabajo se evaluó su influencia en la riqueza y diversidad de aves en un bosque de encino-pino de Chignahuapan, Puebla, bajo tres condiciones: 1ª. No quemado recientemente (más de cinco años, C1); 2ª. Quemado un año antes a baja severidad (C2) y 3ª. Quemado un año antes con alta severidad (C3). Para llevar a cabo el muestreo, se aplicaron los métodos de parcelas circulares y de red de niebla, además, se siguieron las claves correspondientes para identificación de avifauna. Los resultados condujeron al registro de 30 especies de aves, pertenecientes a seis órdenes y 20 familias; 22 de dichas especies se agrupan en C1, 12 en C2 y 10 en C3. Se estimó la diversidad de la avifauna con el índice de Shannon-Wiener para cada condición. Con base en la prueba t Hutchenson, se verificaron diferencias en diversidad entre C2 (2.079) y C3 (1.040) a partir de la red de niebla; y, con las parcelas circulares, C1 (2.198) difirió de C3 (1.850). Con base en los resultados encontrados, se concluye que el fuego es un factor ecológico que provoca cambios importantes en la diversidad y riqueza de aves a corto plazo.
Palabras clave: Avifauna, encinar, incendios forestales, índices de diversidad, quemas prescritas, severidad del fuego.
Abstract
From an ecological point of view, fire has a positive effect on wildlife and their habitat in fire dependant ecosystems and under fire regimes not very altered. There are very few studies regarding the effects of fire on birds in Mexico The aim of this study was to evaluate the influence of fire on the richness and diversity of birds in oak-pine woodland in Chignahuapan, Puebla State, Mexico, under three different conditions: 1st) not recently burned (more than five years, C1); 2nd) burned the year before at low severity (C2) and 3rd) burned the year before at high severity (C3). For bird sampling, circular lots and mist nets were used, as well as taxonomic keys for bird identification. Results lead to the record of 30 species of birds, which belong to six orders and 20 families; 22 of those species are grouped in C1, 12 in C2 and 10 in C3. Bird diversity was determined by the Shannon-Wiener index for each condition. According to the t Hutchenson test, differences were confirmed between C2 (2.079) and C3 (1.040), from the mist net method and from the circular plots, differences were recorded for C1 (2.198) and C3 (1.850). Based on the aforementioned results, it is concluded that fire is an ecological factor that causes significant changes in richness and diversity of birds on the short run.
Key words: Birds, oak forests, diversity indexes, forest fires, prescribed fires, fire severity.
Introducción
El fuego es un elemento esencial que desempeña una función importante en la salud de los ecosistemas que dependen de él. Desde hace miles de años, los seres humanos lo han usado como una herramienta de ordenación de la tierra; sin embargo, debido al cambio climático y al uso sin control del fuego mismo, sus regímenes alterados son una amenaza para los bosques y su biodiversidad (FAO, 2001; Shliski et al., 2007). Aquellos cuya permanencia es determinada por este factor cuentan con especies que han evolucionado en su presencia, por lo cual dicho elemento resulta esencial para el mantenimiento de la diversidad biológica en diferentes paisajes. Si se excluyen o inducen incendios de manera inadecuada, los ecosistemas pueden ser alterados seriamente (Shlisky et al., 2007). Este es el caso del bosque de encino-pino, en donde el fuego aparte de contribuir a mantener la diversidad, reciclar nutrimentos y contribuir a que las coníferas y los encinos que están adaptados a tal factor prevalezcan, también crea ambientes favorables para muchos animales silvestres (Rodríguez et al., 2001; Rodríguez y Myers, 2010); por lo tanto, su monitoreo es de especial interés, pues se les considera indicadores biológicos del estado de un ecosistema (González, 2003), como la avifauna.
En México, 45.5% de las zonas forestales presentan disturbio, por lo que este tipo de organismos podría ayudar a determinar su calidad ambiental, ya que reúnen cualidades como la facilidad para identificarlos, visual y auditivamente a distancia; además, muchos taxa son consumidores secundarios, por lo que concentran efectos contaminantes o muestran cambios en la cadena trófica, lo que resulta adecuado para el monitoreo de impactos en el ambiente (Villaseñor y Santana, 2003). Whelan (1995) señala que las aves son candidatas para llevar a cabo estudios de los efectos directos del fuego en los vertebrados.
En este contexto, Cabrera et al. (2006) demuestran que un buen manejo del fuego origina un mosaico de pastos y arbustos que promueven la diversidad y un hábitat propicio para algunas aves como Xenospiza baileyi Bangs (gorrión serrano), cuya alimentación es fundamentalmente granívora y que está en peligro de extinción. Así mismo, Ugalde-Lezama et al. (2010) señalan una mayor diversidad de aves en bosques mixtos con disturbio en Zoquiapan, Edo. de Méx., con respecto a los que presentan poco disturbio.
Diversos especialistas en ecología del fuego, como Whelan (1995), afirman que se debe poner más énfasis en los beneficios que este le provee a los ecosistemas. No obstante, las consecuencias del uso de esta herramienta de manejo y sus impactos en la biodiversidad requieren de una revisión cuidadosa (Pons et al., 2003).
Los incendios forestales son una de las principales amenazas para los bosques (Céspedes, 2002). Paradójicamente, el fuego es un factor ecológico presente en la mitad de los tipos de vegetación y en más de 40% de la superficie con vegetación en México (Rodríguez, 2008); aunque sus efectos en las aves han recibido muy poca atención.Por lo anterior, el objetivo del presente trabajo fue evaluar la influencia del fuego a diferentes severidades en la riqueza y diversidad de las especies de aves presentes en un bosque de encino-pino de Chignahuapan, Puebla.
Materiales y Métodos
Chignahuapan se localiza en la parte norte del estado de Puebla; las coordenadas geográficas del área de estudio son: 19°48'29" de latitud norte y 97°59'35" de longitud oeste, a 2,525 m de altitud (Figura 1). De acuerdo con García (1973) predomina el clima templado, con una temperatura media anual de 12.6 °C y precipitación media anual de 1,096.6 mm. La vegetación corresponde a bosques de encino-pino, donde dominan Quercus crassifolia Humb. et. Bonpl., Pinus teocote Schltdl. et Cham y P. patula Schlecht. et. Cham (Rzedowski, 1978). La ubicación de las parcelas circulares y redes de niebla se exhibe en el plano coordenado de la Figura 1.
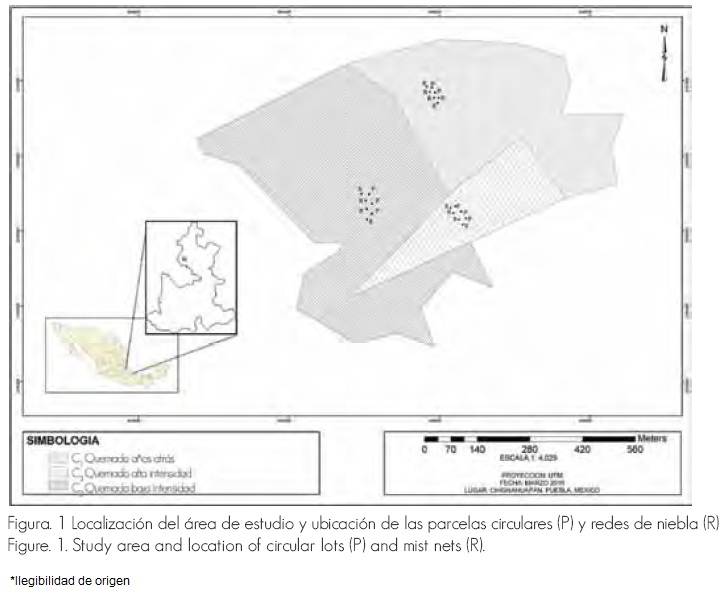
Figura. 1 Localización del área de estudio y ubicación de las parcelas circulares (P) y redes de niebla (R).
Figure. 1. Study area and location of circular lots (P) and mist nets (R).
La riqueza y diversidad de especies de aves se determinó mediante una comparación con diferente grado de afectación o tratamientos entre tres condiciones: encinares no afectados en los últimos cinco años por incendios forestales (C1), con una superficie aproximada de 20 ha; encinares quemados a baja severidad (C2) sobre 28.9 ha, y encinares quemados con alta severidad (C3) en una superficie de 6.89 ha; los dos últimos ocurrieron el año anterior al muestreo. La causa de los incendios fue promover la emisión de pelillo con fines de pastoreo.
Los niveles de severidad se definieron con base en la altura de la cicatriz que el fuego deja sobre el tronco y el nivel de afectación de la copa. La categorización propuesta fue:
1. Área no afectada recientemente (más de cinco años): sin señas evidentes de incendio (C1).
2. Severidad baja: estrato arbóreo con "chamuscados" de copa inferiores a 50% y cicatrices sobre el tronco <1 m (C2).
3. Severidad alta: estrato arbóreo con "chamuscados" de copa de 50% o más y cicatrices sobre el tronco >1 m (C3).
Para las tres condiciones, Quercus crassifolia Humb. resultó ser la especie arbórea dominante. En C1 existen arbustos: Bouvardia ternifolia (Cav.) Schlecht., Ceanothus coeruleus Lag., y el estrato herbáceo está dominado por Muhlenbergia macroura (HBK.). Para el caso de C2, en el dosel intermedio se identifican a Baccharis salicifolia (Ruiz & Pavón) Pers., Eupatorium ligustrinum D.C. y Quercus repanda Bonpl. El estrato herbáceo está representado por Conyza shiediana (Less.) Cronquist, Polypodium guttatum Maxon. y Pteridium feei (W. Schaffn. ex Feé) Faull, entre otros. En la zona C3 se presentan Agrostis sp., Piptochaetium fimbratum (HBK.) Hitchc., Muhlenbergia macroura (Kunth) Hitchc., Stipa ichu (Ruiz & Pav.) Kuth y Archibaccharis serratifolia (Kunth) S.F. Blake.
El trabajo de campo comprendió tres salidas en junio, octubre y noviembre del 2009. Se realizó un muestreo dirigido derivado de visitas prospectivas en el que se definió un tamaño de muestra de 500 m2. El tipo de muestreo respondió a la relativa heterogeneidad de afectación por el fuego en cada una de las condiciones, por lo que se buscaron áreas representativas. Los polígonos fueron georreferenciados con un Sistema de Posición Global (GPS) marca Etrex Garmin.
Los datos de las comunidades de aves se obtuvieron mediante los métodos de recuento en puntos y redes de niebla. Se usaron ambas técnicas por el comportamiento quieto y silencioso de algunas aves, lo que dificulta su identificación (López de Casenave et al., 1998). Con el método de recuento en puntos con radio fijo (Wunderle, 1994) se detectaron todos los individuos en un radio de 25 m por un periodo de 15 min. Se establecieron de manera irregular tres de estos puntos por condición, con una distancia mínima de 100 m entre ellos, para dar una cobertura equitativa. La observación de las aves se hizo con binoculares de 25x50 m marca Bushnell y para su identificación se utilizaron guías de campo (Howell y Webb, 1995; Peterson y Chalif, 2008; National Geographic, 1999; Kaufman, 2005; Udvardy, 2005). Se elaboró una ficha de registro por punto para acumular la información de las aves observadas.
Con base en la disponibilidad del material para la captura, identificación y liberación de las aves se colocaron cuatro redes de niebla: dos de 12 m y dos de 6 m en cada condición del bosque. Estas se ubicaron de manera sistemática con una distancia mínima de 100 m entre ellas. Las revisiones se realizaron por tratamiento de 7 a 12 y de 16 a 19 h, cada 45-60 min, para sumar 72 revisiones.
La riqueza de especies se calculó por el método de muestreo por separado (redes de niebla y puntos de conteo), y representó el número total de especies registradas en cada parcela. A efecto de establecer las diferencias entre riqueza por área y método de muestreo, se aplicó la prueba de Kruskal-Wallis (Zar, 1999). Para cada especie fue asignado un estatus de permanencia en el área (Bojorges y López-Mata, 2006): local o migratoria (Peterson y Chalif, 2008).
A los datos por área se les aplicó el índice de diversidad de Shannon-Wiener (H') (ecuación 1). Posteriormente los valores se sometieron a la prueba de t Hutchenson (Zar, 1999).
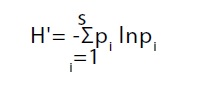 (1)
(1)
Donde:
H'= Índice de diversidad de Shannon-Winer
pi=Abundancia relativa
ln= logaritmo natural
S=Número total de especies en la muestra
También su determinó la similitud de especies entre condiciones, con el coeficiente de comunidad de Jaccard y el de similitud de Sorensen (Magurran, 1988) (ecuaciones 2 y 3).
 (2)
(2)
Donde:
a= Número de especies en la comunidad A
b= Número de especies en la comunidad B
j= Número de especies comunes en ambas comunidades
 (3)
(3)
Donde:
Cs=Coeficiente de Sorensen
a= Número de especies en la comunidad A
b= Número de especies en la comunidad B
j= Número de especies comunes en ambas localidades
Resultados y Discusión
Se registraron 30 especies de aves (17 locales y 13 migratorias), pertenecientes a seis órdenes y 20 familias con predominancia del Passeriformes (22 especies), seguido de los órdenes Strigiformes, Falconiformes, Apodiformes, Columbiformes y Trogoniformes (Cuadro 1).
Cuadro 1. Listado de especies por condición, tipo de muestreo en el que fueron observadas y su distribución.
Table 1. List of species per condition, type of sampling in which they were observed and their distribution.

Algunas aves se atraparon en una sola ocasión con el método de red de niebla, y otras se observaron una vez en la parcela circular, lo que se puede atribuir a la baja densidad predominante durante los muestreos o a disturbios antrópicos, como el cambio de uso del suelo en las cercanías, que han ejercido presión sobre estas áreas (Chi, 2007).
La combinación de ambas técnicas permitió el registro de 22, 12 y 10 especies en las áreas C1, C2 y C3, respectivamente. Se identificaron 20 taxa con la red de niebla, de los cuales 12 fueron capturados (ninguno se observó en las parcelas circulares), si bien siete de los no capturados también fueron consignados con el método de parcelas circulares. En el Cuadro 2 se resumen los resultados de riqueza y diversidad de aves por medio de redes de niebla y parcelas circulares, bajo los tres tratamientos.
Cuadro 2. Resumen de datos obtenidos por medio de los dos métodos de conteo de aves.
Table 2. Data summary obtained from both bird counting methods.
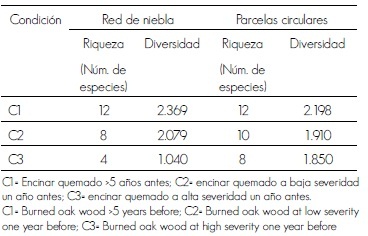
Para el método de red de niebla y con la prueba de t Hutchenson se determinó que hubo diferencia significativa (p<0.05) en la riqueza de especies entre C2 y C3. Respecto al de parcelas circulares, con la misma prueba estadística resultaron con diferencias significativas C1 y C3 (p<0.05).
La mayor diversidad de aves en C1 coincide con lo señalado por Prodon et al. (1987) para un bosque mediterráneo de encino, donde los primeros años después de un incendio hay disminución de la riqueza de aves; posteriormente, se incrementa, una vez recuperada la vegetación. El régimen actual de fuego en la región mediterránea, propensa a los incendios, es probable que haya contribuido a mantener la diversidad de aves en Portugal (Moreira et al., 2001).
El área quemada con alta severidad tuvo mayor riqueza para el muestreo de noviembre; a su vez, Kotliar et al. (2007) citan que antes y después de un incendio, a lo largo de gradientes de severidad de quemas, la riqueza es similar a la existente antes del evento. Según Main y Tanner (2003) algunas especies que se alimentan de forraje hacen uso de áreas quemadas recientemente para la alimentación de sus polluelos. Por lo tanto, el fuego puede influir de manera positiva en poblaciones de vida silvestre. Esto significa que el mantenimiento de las superficies en etapas sucesionales previas al clímax puede favorecer la diversidad, ya que el papel del fuego en la alteración de la sucesión vegetal es reconocido, y se ha utilizado por largo tiempo (Wildlife Society, 1987). En muchos sentidos tiene un efecto semejante al inicio de la primavera, pues ambos sucesos representan un nuevo comienzo para las comunidades vegetales y para la vida silvestre que depende de ellos (Main y Tanner, 2003). Margalef (1968) afirma que todo disturbio suele ser un punto de partida en una sucesión ecológica; además de, un proceso reestructurador de los sistemas naturales.
En C2 y C3 se obtuvo mayor similitud de especies de aves, lo que se debe a que comparten seis de los 16 taxa que se detectaron en ambas. Las áreas C1 y C2 muestran la menor similitud. Las áreas C2 y C3 están directamente relacionadas, ya que las dos fueron sometidas a un disturbio reciente, y la vegetación muestra más complejidad estructural en C1, por el tiempo que ha pasado desde el disturbio (Ugalde-Lezama et al., 2009). El Cuadro 3 resume los resultados de los índices de similitud calculados.
Cuadro 3. Coeficientes de similitud de Sorensen (Cs) y Jaccard (Cj) entre condición, para red de niebla y parcela circular, en el bosque de encino-pino de Chignahuapan, Puebla.
Table 3. Sorensen Cs) and Jaccard (Cj) similarity coefficients per condition for mist net and circular lot in the oak-pine wood of Chignahuapan, Puebla.

Las condiciones C2 y C3 son más similares debido a que comparten las siguientes aves: Certhia americana Bonaparte, Pipilo erythrophthalmus Linnaeus, Cyanocitta stelleri Gmelin, Hylocharis leucotis Vieillot, Myadestes obscurus Gmelin y Trogon mexicanus Swainson, si bien las últimas cuatro estuvieron en las tres condiciones.
En los dos tratamientos quemados recientemente se identificaron ocho especies exclusivas, de las cuales cuatro correspondieron a C2 (Dendroica occidentalis Townsend, Ergaticus ruber Swainson, Regulus calendula Linnaeus y Wilsonia pusilla Wilson y dos de C3 (Dendroica townsendi Townsend y Ergaticus ruber Swaison, Regulus calendula Linnaeus,Wilsonia pusilla Wilson y dos de C3 (Dendroica townsendi Townsend y Turdus migratorius Linnaeus). Certhia americana Bonaparte y Pipilo erythrophthalmus Linnaeus se ubicaron en ambas condiciones. La taxa registrados como únicos para cada caso podrían atribuirse a que son territoriales, pues acorde con Whelan (1995) algunas aves se quedan en su territorio, aunque esté quemado, y las otras se desplazan temporalmente a zonas sin afecctación por el fuego.
Ergaticus ruber, Regulus calendula, Dendroica townsendii, Turdus migratorius y Certhia americana fueron detectadas tanto en bosques con disturbio, como en aquellos afectados recientemente por el fuego en Zoquiapan, Edo de Méx. (Ugalde-Lezama et al., 2010). También se han documentado aumentos en las poblaciones de Certhia americana y Turdus migratorius posteriores a quemas prescritas de baja severidad (Bagne, 2011); un comportamiento igual presentan T. migratorius y Cyanocitta stelleri en bosques de Pinus ponderosa Douglas ex C. Lawson (Dickson et al., 2009). A su vez, el primero es un insectívoro aéreo que responde favorablemente a las quemas prescritas en bosques de Pinus ponderosa del oeste de Estados Unidos de América (Russell et al., 2009). Así mismo se ha observado que emplea las latifoliadas para anidar, de manera importante, de modo tal que Quercus spp. son un recurso valioso para esta y otras aves que se reproducen en ese tipo de vegetación. Por ello, como parte del manejo de su hábitat, se recomienda mantener encinos maduros en las masas de pino, crear claros para favorecer su crecimiento y realizar actividades de manejo, como las quemas prescritas, que promuevan la regeneración de encinos que remplacen a los ejemplares muertos (Purcell y Drynan, 2008).
En Ohio se usan quemas prescritas para mantener y restaurar los bosques mixtos de Quercus spp., en los cuales la densidad de Turdus migratorius se incrementa, siempre que sean repetidas durante varios años (Artman et al., 2001).
Regulus calendula es resiliente al fuego en bosques riparios, que no se queman con frecuencia, si bien su masa se reduce en las localidades incendiadas (Samuels et al., 2005). Dendroica townsendi se registró en un sitio quemado con alta severidad, pero Tiedemann y Woodard (2002) anotan que dicha especie declinó sus poblaciones o estuvo ausente en bosques de Pinus contorta Douglas ex Loudon y Abies lasiocarpa (Hook.) Nutt. con quemas prescritas muy intensas, de reemplazo de vegetación.
A su vez, Dendroica occidentalis disminuyó luego de quemas prescritas, incluso a baja severidad (Bagne, 2011). De manera similar, Pipilo erythrophthalmus resultó más abundante en sitios no quemados de encinares deciduos del medio oeste de los Estados Unidos de América; sin embargo, la riqueza de especies de aves no difirió entre sitos, con o sin quemas prescritas (Aquilani et al., 2003). No obstante, en bosques mixtos del centro de México, se le observó en áreas quemadas moderadamente, mas no en las sin afectación (Ugalde-Lezama et al., 2010).
En el presente trabajo, Hylocharis leucotis se identificó, principalmente, en el área quemada con alta severidad. Se trata de un taxón de hábitos nectívoros, que es atraída por el color rojo o naranja de las flores, a lo que debe agregarse que su pico largo y delgado les permite extraer néctar de flores con corola tubular como Bouvardia ternifolia presente en dicha condición y en la estación del año en la cual se realizó el estudio En otras investigaciones que no incluyen localidades incendiadas Lara (2006) y Ortiz-Pulido y Vargas-Licona (2008) han determinado una relación entre colibríes (Hylocharis leucotis) y la abundancia floral de especies como Bouvardia ternifolia.
Accipiter bixolor Vieillot, Atlapetes pileatus Wagler,Basileuterus belli Giraud, Buteo jamaicencis Gmelin, Campylorhynchus rufinucha Lesson, Caprimulgus ridgwayi Nelson, Caprimulgus vociferus Wilson, Catharus occidentalis Sclater, Diglossa baritula Wagler, Empidonax difficilis Baird, Habia fuscicauda Cabanis, Myioborus miniatus Swainson, Sialia mexicana Swainson y Vireolanius melitophrys Bonaparte se identificaron solo en el área quemada cinco años atrás. En el centro de México, Atlapetes piliatus, Empilonax difficilis y Myioborus miniatus fueron registradas en bosques de coníferas moderadamente disturbados por fuego, pero estuvieron ausentes en los no afectados. Por su parte, Catharus occidentalis, Diglossa baritula y Sialia mexicana se observaron en ambas condiciones (Ugalde-Lezama et al., 2010).
Sialia mexicana se beneficia por tratamientos de aclareos y quemas prescritas frecuentes y de baja intensidad en los bosques de Pinus ponderosa (Gaines et al., 2007; Dickson eet al., 2009; Hurteau et al., 2010); aunque la retención de árboles muertos en pie puede ser problemática en áreas de quemas prescritas (Hurteau et al., 2010). Las actividades de supresión de incendios forestales conducen a una pérdida de hábitat para estas aves, puesto que el fuego mantiene baja la densidad del arbolado, lo cual deriva en mayor abundancia de herbáceas y de invertebrados que le sirven de alimento (Wightman y Germaine, 2006).
La abundancia y riqueza de aves tuvo cambios significativos entre condiciones y muestreos, principalmente, entre C1 y C3, ya que para la segunda fueron muy bajas en los meses de junio y octubre, pero en el mes de noviembre fue más diversa, esto debido a la cantidad de ejemplares migratorios de invierno que arriban en busca de cualidades que el área quemada recientemente les proporciona.
Hutto (1995) (citado por Smith, 2000) señala la presencia de 15 especies de aves más en localidades quemadas poco tiempo antes de la evaluación en el norte de las Montañas Rocosas que en hábitats no quemados recientemente. Algunas comunidades de aves, muestran una gran capacidad de resistencia a los disturbios, por eso el fuego no suele acarrear grandes cambios en ellas (Pons, 2005); además es un factor ecológico que provoca modificaciones en el ecosistema. Bajo un régimen de fuego adecuado, tales cambios son favorables para la avifauna.
La influencia del fuego propicia la heterogeneidad del bosque (cobertura, especies); así, los incendios aportan espacio a la diversidad (por ejemplo, entre áreas quemadas y no quemadas) y tiempo (desde la ocurrencia del fuego), de tal manera que la suma del efecto de las distintas severidades de este fenómeno, permite su análisis integral. Esto se refleja en la mayor riqueza y diversidad de aves en conjunto que se determinó para las tres áreas quemadas, si se le compara con los valores correspondientes a cada una de ellas por separado.
Conclusiones
Las zonas con incendios recientes tuvieron menor riqueza, diversidad y abundancia de aves, con respecto a la incendiada más de cinco años antes, lo que indica que los impactos del fuego al primer año suelen disminuir tales especies, cada una de las cuales hace uso del hábitat para buscar áreas donde encuentran refugio, disponibilidad de alimento y cobertura arbórea para percha.
La riqueza y diversidad de aves en las tres áreas conjuntamente supera las de cada condición individual, lo que denota el aporte a la diversidad de la avifauna de distintos niveles de disturbio por fuego y el tiempo involucrado en cada una.
Muchas de las especies identificadas en el presente trabajo, también se han registrado como típicas de ambientes con disturbio por el fuego tanto en México como en los Estados Unidos de América.
Agradecimientos
Al CONACYT, a la DICIFO, UACH y a la CONAFOR estado de Puebla. Particularmente al Sr. Juan Sánchez y al Biól. Andrés Gelasio Miranda Moreno, así como a Miriam, Paty, Griselda y Marco, por su valioso apoyo y colaboración en campo. Esta investigación forma parte del Proyecto Ajusco de la UACH, sobre ecología del fuego, manejo integral del fuego y restauración de áreas incendiadas.
Referencias
Aquilani, S. M., T. E. Morrell, and D. C. LeBlanc. 2003. Breeding bird communities in burned and unburned sites in a mature Indiana oak forest. Proceedings of the Indiana Academy of Science 112(2): 186-191. [ Links ]
Artman, V. L., E. K. Sutherland and J. F. Downhower. 2001. Prescribed burning to restore mixed-oak communities in Southern Ohio: effects on breeding-bird populations. Conservation Biology 15(5): 1423-1434. [ Links ]
Bagne, K. E. 2011. Short term responses to prescribed fire in fire-supressed forests of California. Journal of Wildlife Management 75(5): 1051-1060. [ Links ]
Bojorges B., J.C. y L. López-Mata. 2006. Asociación de la riqueza y diversidad de especies de aves y estructura de la vegetación en una selva mediana subperennifolia en el centro de Veracruz, México. Revista Mexicana de Biodiversidad 77: 235-249. [ Links ]
Cabrera G., L., J. Velázquez M. and W. Escamilla E. 2006. Identification of priority habitats for conservation of the Sierra Madre sparrow Xenospiza baileyi in Mexico. Oryx 40(2): 211-217. [ Links ]
Céspedes F., C. 2002. Evaluación y análisis geográfico de la diversidad faunística de Chiapas. CONABIO. San Cristóbal de las Casas, Chis. México. pp. 11-21. [ Links ]
Chi P., R. A. 2007. Avifauna en bosque manejado de Pinus patula en la mojonera, Zacualtipán, Hidalgo. Tesis de Maestría en Ciencias. Colegio de Postgraduados, Montecillo, Edo. de Méx., México. 75 p. [ Links ]
Dickson, B. G., B. R. Noon, C. H. Flather, S. Jentsch and W. M. Block. 2009. Quantifying the multiscale response of avifauna to prescribed fire experiments in the southwest United States. Ecological Applications 19(3): 608-621. [ Links ]
Food and Agriculture Organization (FAO). 2001. The Global Forest Resources Assessment 2000 - main report. Estudio FAO Montes Nº 140. Rome, Italy. 479 p. [ Links ]
Gaines, W. L., M. Haggard, J. F. Lehmkuhl, A. L. Lyons and R. J. Harrod. 2007. Short term response of land birds to Ponderosa pine restoration. Restoration Ecology 15(4): 670-678. [ Links ]
García, E. 1973. Modificaciones al sistema de clasificación climática de Köppen. Universidad Nacional Autónoma de México. Instituto de Geografía. México, D. F. México. 246 p. [ Links ]
González O., J. A. 2003. Aplicación de análisis multivariantes al estudio de las relaciones entre las aves y sus hábitats: un ejemplo con paseriformes montanos no forestales. Ardeola 50(1):47-58. [ Links ]
Howell, S. N. G. and S. Webb. 1995. A guide to the birds of Mexico and Northern Central America. Oxford University Press, New York, NY. USA. 851 p. [ Links ]
Hurteau, S. I., B. Dickson and W. Bloc. 2010. Variability in nest density, occupancy and home range size of western bluebirds after forest treatment. Forest Science 56(1): 131-138. [ Links ]
Kaufman, K. 2005. Guía de campo, las aves de Norteamerica. Hillstar Editions L. C. México, D. F. México. 391p. [ Links ]
Kotliar, N. B., P. L. Kennedy and K. Ferree. 2007. Avifaunal responses to fire in Southwestern montane forests along a burn severity gradient. Ecological Aplications 17(2): 491-507. [ Links ]
Lara, C. 2006. Temporal dynamics of flower use by hummingbirds in a highland temperate forest in Mexico. Ecoscience 13(1): 23-29. [ Links ]
López de Casenave, J., J. P. Pellot, S. M. Caziani, M. Mermoz and J. Protomastro. 1998. Responses of avian assemblages to a natural edge in a chaco semiarid forest in Argentina. The Auk 115(2): 425-435. [ Links ]
Magurran, A. E. 1988. Diversidad ecológica y su medición. Princeton University Press. New York, NY. USA. 200 p. [ Links ]
Main, M. B. and G. W. Tanner. 2003. Effects of fire on Florida's wildlife and wildlife habitat. Wildlife Ecology and Conservation Department 137. Department of Agriculture, Cooperative Extension Service, University of Florida, Florida. Gainesville, FL. USA. 4 p. [ Links ]
Margalef, R. 1968. Perspectives in ecological theory. University of Chicago Press. Chicago,IL. USA. 111 p. [ Links ]
Moreira, F., G. Ferreira P., C. Rego F. and S. Bunting. 2001. Landscape changes and breeding bird assemblages in northwestern Portugal: the role of fire. Landscape Ecology 16(2): 175-187. [ Links ]
National Geographic. 1999. Field guide to the birds of North America. 3rd ed. National Geographic Society, Washington, DC. USA. 480 p. [ Links ]
Ortiz-P., R. and G. Vargas-L. 2008. Exploring the relationship between humming bird records and flower abundance with spatio-temporal scaling. Ornitología Neotropical 19: 473-483. [ Links ]
Peterson, R. T. y E. L. Chalif. 2008. Aves de México: guía de campo. Editorial Diana. México, D. F. México. 473 p. [ Links ]
Pons, P. 2005. Consecuencias de los incendios forestales sobre los vertebrados y aspectos de su gestión en regiones mediterráneas. http://www.udg.edu/portals/92/Bio%20Animal/pdf/Pons%20inCamprodon-Plana2007.pdf. pp. 229-245. (noviembre, 2009). [ Links ]
Pons, P., B. Lambert, E. Rigolot and R. Prodon. 2003. The effects of grassland management using fire on habitat occupancy and conservation of birds in a mosaic landscape. Biodiversity and Conservation 12: 1843-1860. [ Links ]
Prodon, R., R. Fons and F. Athias-Binche. 1987. The impact of fire on animal communities in the Mediterranean area. In: Trabaud, L. (Ed.). The role of fire in ecological systems. S. P. B. Academic. The Haghe. The Hague, The Netherlands. pp. 121-157. [ Links ]
Purcell, K. L., and D. A. Drynan. 2008. Use of hardwoods by birds nesting in ponderosa pine forests. Pacific Southwest Research Station. United States Department of Agriculture, Forest Service. Gen. Tech. Rep. PSW-GTR-217. Albany, CA, USA. pp. 417-431. [ Links ]
Rodríguez T., D. A. 2008. Fire regimes, fire ecology, and fire management in Mexico. Ambio 37(7-8): 548-556. [ Links ]
Rodríguez T., D. A., M. Rodríguez- A., F. Fernández S. y S. J. Pyne. 2001. Educación e incendios forestales. 2a ed. Mundi Prensa. México, D. F., México. 201 p. [ Links ]
Rodríguez, T., D. A. and R. L. Myers. 2010. Using oak characteristics to guide fire regime restoration in Mexican pine-oak and oak forests. Ecological Restoration 28: 304-323. [ Links ]
Russell, R. E., J. A. Royle, V. A. Saab, J. F. Lehmkuhl, W. M. Block and J. R. Sauer. 2009. Modelling the effects of environmental disturbance on wildlife communities: avian responses to prescribed fire. Ecological Applications 19(5): 1253-1263. [ Links ]
Rzedowski, J. 1978. Vegetación de México. Limusa. México, D.F. México. 432 p. [ Links ] Samuels, I. A., T. Gardali, D. L. Humple and G. R. Geupel. 2005. Winter site fidelity and body condition of three riparian songbird species following a fire. Western North American Naturalist 65(1): 45-52. [ Links ]
Shlisky, A., J. Waugh, P. González, M. González, M. Manta, H. Santoso, E. Alvarado, A. Ainuddin Nuruddin, D. A. Rodríguez-Trejo, R. Swaty, D. Schmidt, M. Kaufmann, R. Myers, A. Alencar, F. Kearns, D. Johnson, J. Smith, D. Zollner y W. Fulks. 2007. El fuego, los ecosistemas y la gente: amenazas y estrategias para la conservación global de la biodiversidad. Informe Técnico de la Iniciativa Global para el Manejo del Fuego 2007-2. The Nature Conservancy. Arlington, VA. USA. 20 p. [ Links ]
Smith, J. K. 2000. Wildland fire in ecosystems: effects of fire on fauna. United States Department of Agriculture, Forest Service, Rocky Mountain Research Station. Gen. Tech. Rep. RMRS-GTR-42-vol. 1. Ogden, UT. USA. 83 p. [ Links ]
Tiedemann, A. R. and P. M. Woodard. 2002. Multiresources effects of a stand replacement prescribed fire in the Pinus contorta-Abies lasiocarpa vegetation zone of central Washington. United States Department of Agriculture, Forest Service. Pacific North West Research Station. Gen. Tech. Rep. PNW-GTR-535. Portland, OR. USA. 27 p. [ Links ]
Udvardy, M. D. F. (Ed.). 2005. The Audubon Society Field Guide to North America Birds. Western Region. A. A. Knopf. New York, NY, USA. 854 p. [ Links ]
Ugalde-Lezama, S., J. I. Valdez-Hernández, G. Ramírez-Valverde, J. L. Alcántara-Carbajal y J. Velázquez-Mendoza. 2009. Distribución vertical de aves en un bosque templado con diferentes niveles de perturbación. Madera y Bosques 15(1): 5-26. [ Links ]
Ugalde-Lezama, S., J. L. Alcántara C., J. I. Valdéz H., G. Ramírez V., J. Velázquez M. y L. A. Tarango A. 2010. Riqueza, abundancia y diversidad de aves en un bosque templado con diferentes condiciones de perturbación. Agrociencia 44(2): 159-169. [ Links ]
Villaseñor G., J. F. y E. Santana C. 2003. El monitoreo de poblaciones: herramienta necesaria para la conservación de aves en México. In: Gómez D., H., y D. A. Oliveras (Eds.). Conservación de aves. Experiencias en México. Sociedad para el Estudio y Conservación de las Aves en México A.C. Comisión Nacional para la Conservación de la Biodiversidad, Sociedad para el Estudio y Conservación de las Aves de México, A. C.. México, D. F. México. pp. 224-262. [ Links ]
Whelan, R. J. 1995. The ecology of fire. Cambridge University Press. Cambridge, UK. 346 p. [ Links ]
Wightman, C. S. and S. S. Germaine. 2006. Forest stand characteristics altered by restoration affect western blue bird habitat quality. Restoration Ecology 14(4): 653-661. [ Links ]
Wildlife Society. 1987. Manual de técnicas de gestión de vida silvestre. United States of America for the Wildlife Society, Inc. Bethesda, MD. USA. 703 p. [ Links ]
Wunderle, J. M. 1994. Census methods for Caribbean land birds. United States Department of Agriculture, Forest Service. Southern Forest Experimental Station. Gen. Tech. Rep. SO-100. New Orleans, LA. USA. 24 p. [ Links ]
Zar, J. H. 1999. Biostatistical analysis. 4th edition. Prentice Hall Inc. New York, NY. USA. 663 p. [ Links ]

