










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de ciencias forestales
versión impresa ISSN 2007-1132
Rev. mex. de cienc. forestales vol.2 no.7 México sep./oct. 2011
Artículos
Propagación y micorrización de plantas nativas con potencial para restauración de suelos
Propagation and mycorrhization of native plants with soil restoration potential
Laura Hernández-Cuevas 1, Vidal Guerra-De la Cruz2*, Guadalupe Santiago-Martínez 1 y Porfirio Cuatlal-Cuahutencos 1
1 Laboratorio de Micorrizas, CICB, Universidad Autónoma de Tlaxcala.
2 Sitio Experimental Tlaxcala-CIRCE-INIFAP. *Correo-e: guerra.vidal@inifap.gob.mx
Fecha de recepción: 3 de diciembre de 2009.
Fecha de aceptación: 17 de agosto de 2011.
RESUMEN
Se presentan los resultados obtenidos al propagar e inocular con hongos micorrizógenos arbusculares a Amelanchier denticulata (tlaxistle) y Eysenhardtia polystachya (palo dulce), plantas nativas de América, que forman micorriza arbuscular y crecen en México sobre suelos someros y con escasez de nutrimentos. La propagación se hizo a partir de semillas, en vermiculita y turba mezcla (2:1). La mitad de las plantas de cada especie se inoculó con una mezcla de tres cepas de hongos micorrizógenos arbusculares aisladas de suelos de Tlaxcala: Glomus claroideum, Acaulospora laevis y A. morrowiae . El tlaxistle y el palo dulce alcanzaron valores por arriba del 40% de germinación y del 80% de supervivencia al trasplante. Aunque, los porcentajes de colonización micorrízica fueron bajos en los dos casos, menores al 10% para palo dulce y al 20% para tlaxistle, los individuos micorrizados mostraron las mejores respuestas en diámetro, altura, biomasa aérea (p<0.001) y contenido de fósforo. También se analizan algunas variables indicadoras de calidad de planta en relación a la micorrización. Se concluye que la propagación a partir de semillas es una buena estrategia para la obtención de plantas de los dos taxa estudiados y que la micorrización favorece significativamente su desarrollo inicial, ya que mejora sus rasgos de crecimiento, lo que es crítico para las plantas que serán destinadas al campo en programas de restauración.
Palabras clave: Acaulospora laevis Gerdemann et Trape, Acaulospora morrowae Spain et Schenk, Amelanchier denticulata (Kunth) W. D. J. Koch, Eysenhardtia polystachya (Ortega) Sarg., Glomus claroideum Schenk et Smith, hongos micorrizógenos arbusculares.
ABSTRACT
The results of propagation and inoculation with arbuscular mycorrhiza of Amelanchier denticulata ("tlaxistle") and Eysenhardtia polystachya ("palo dulce"), American native plants, which form arbuscular mycorrhiza and commonly grow in Mexico on shallow and nutrient-poor soils are described. Seeds of both species were collected from wild populations in Tlaxcala, and germinated in a vermiculite-peat moss (2:1) mixture. One half of the plants for each species was inoculated with a mix of three arbuscular mycorrhizal strains isolated from Tlaxcala soils: Glomus claroideum, Acaulospora laevis and A. morrowiae . Both plant species showed a good response to propagation with over 40% of germination and 80% of transplantation survival. Mycorrhization was low in both species, less than 10% in sweet stick and less than 20% in "tlaxistle"; however, basal diameter, total height, aerial biomass, and Phosphorus content in mycorrhized plants revealed significant differences (p<0.001) compared to those of non-mycorrhized plants. Other plant quality indicators were also analyzed as related to mycorrhization. It was concluded that propagation from seeds is an appropriated way to obtain seedlings of these two species, and mycorrhization significantly improves development of favorable traits on plants in early stages, keeping good rates of survival and plant growth, which are critical for plants to be used in restoration programs.
Key words: Acaulospora laevis Gerdemann et Trape, Acaulospora morrowae Spain et Schenk, Amelanchier denticulata (Kunth) W. D. J. Koch, Eysenhardtia polystachya (Ortega) Sarg., Glomus claroideum Schenk et Smith , arbuscular mycorrhizal fungi.
INTRODUCCIÓN
Se estima que más del 70% de la superficie de la república mexicana está ocupada por diferentes tipos de vegetación, que incluyen bosques templados, tropicales y de zonas áridas (Challenger y Soberón, 2008). No obstante, gran parte ella ha disminuido, o al menos, está deteriorada por diversos factores de origen humano y natural; por ejemplo: la tala desmedida, los incendios forestales, la sobreexplotación de algunas especies útiles y el desmonte para fines agropecuarios, entre otros, los cuales ponen en riesgo el patrimonio biológico que representa la cubierta forestal (Challenger et al ., 2009).
La pérdida de los bosques ocasiona áreas degradadas, en las que la posibilidad de ocurrencia de procesos erosivos graves aumenta, por la formación geológica de los suelos. Estos fenómenos adquieren particular relevancia en la zona denominada Eje Volcánico Transversal, cuyos suelos de origen volcánico y con alto contenido de sílice, cuando quedan descubiertos se compactan fuertemente y se propicia la aparición de duripanes o tepetates, que se caracterizan por su gran dureza y porque sus nutrimentos son escasos o se están en formas químicas poco o no disponibles (Werner, 1992).
Las características de los tepetates hacen que el establecimiento de la vegetación natural sea muy difícil y, en algunos casos, prácticamente imposible. En la región geográfica mencionada, el riesgo de pérdida de la cubierta vegetal es mayor por la gran presión demográfica y su consecuente demanda de espacios y recursos, lo cual ha propiciado el cambio de uso del suelo y un incremento importante en la frecuencia y magnitud de los procesos erosivos (Haulon et al ., 2005).
Una manera de recuperar los ecosistemas dañados es a través de prácticas de reforestación y restauración que incluyen coníferas y latifoliadas, principalmente de los géneros Pinus y Quercus (PRONARE, 2000). En las últimas décadas se ha propuesto el uso de especies menos convencionales, pero con potencial de adaptación a condiciones edafo-climáticas extremas, entre las que destacan las plantas nativas o autóctonas (Montaño y Monroy, 2000; Rondón y Vidal, 2005). En procesos de restauración son importantes las que brindan ventajas adicionales, como el enriquecimiento del suelo con algunos minerales esenciales; por ejemplo el nitrógeno (Ferrari y Wall, 2004; Bashan et al ., 2005) favorece el establecimiento y desarrollo de comunidades microbianas diversas, cuya participación es relevante en el ciclo de nutrientes (García-Oliva, 2005).
Eysenhardtia polystachya (Ortega) Sarg. (palo dulce) y Amelanchier denticulata (Kunth) W. D. J. Koch (tlaxistle) crecen de manera natural en amplias regiones geográficas del país y soportan características ambientales difíciles (Morfín y Camacho, 1987; Terrones et al ., 2007), por lo que desde el siglo pasado se han recomendado como apropiadas para la recuperación de algunas zonas deterioradas de México (Flores y Lindig-Cisneros, 2005). A. denticulata y E. polystachya forman micorriza arbuscular (Varela, 2000), y bajo ciertas condiciones la segunda puede asociarse simbióticamente con bacterias fijadoras de nitrógeno. Estas características promueven el desarrollo vegetal y el enriquecimiento de los suelos, de tal forma que, se sugiere el uso de ambas especies en programas de manejo y restauración ecológica (Morfín y Camacho, 1987; González y Camacho, 2000; Terrones et al ., 2007).
A pesar de lo antes expuesto, se ha dado poca atención a su estudio, ya que solo para E. polystachya existe información sobre su propagación (González y Camacho, 2000), su adaptabilidad a la sequía (González y Cantú, 2001) y su utilización como elemento funcional en sistemas agroforestales (Magdaleno et al ., 2005; Terrones et al., 2007). Así mismo, se ha documentado que la colonización micorrízica extensiva en sus raíces y el micelio extrarradical favorecen la formación de agregados y a la estructura del suelo en terrenos tepetatosos (Crisóstomo et al ., 1991). Terrones et al . (2007) consignan el potencial de adaptación de A. denticulata, pero se desconocen datos sobre su germinación, propagación o de su micorrización en condiciones controladas.
Con base en lo anterior, los objetivos de este trabajo fueron: evaluar la respuesta germinativa de las semillas de E. polystachya y A. denticulata, determinar la supervivencia de las plántulas y el efecto de inocularlas con hongos micorrizógenos arbusculares.
MATERIALES Y MÉTODOS
Especies vegetales
Se recolectaron frutos de Eysenhardtia polystachya (Fabaceae) y Amelanchier denticulata (Rosaceae) entre septiembre y noviembre de 2005, en el Cerro Tepeticpac, municipio Totolac, Tlaxcala. Esta topoforma se sitúa entre los 19° 20' y 19° 21' de latitud norte y los 98° 13' y 98° 14' de longitud oeste, a una altitud de 2,300 a 2,520 m y cuenta con poblaciones silvestres abundantes de los dos taxa. Los frutos del tlaxistle se descarnaron para extraer las semillas, las cuales se lavaron con agua destilada y secaron a 60°C en un horno de convección digital BINDER BD53L por 24 h. Para palo dulce fue suficiente el secado de las vainas por el mismo tiempo a igual temperatura para facilitar su apertura. En ambos casos las semillas se colocaron en una solución de Tween 20 al 2% durante 4 minutos, para romper la tensión superficial; se enjuagaron con agua destilada estéril por 10 minutos, finalmente se dejaron en hipoclorito de sodio al 5%, durante 30 minutos y se enjuagaron tres veces.
Germinación
Se realizó una primera prueba de germinación en tres charolas de 20 x 12 cm desinfectadas con hipoclorito de sodio al 5%, con una mezcla de vermiculita: turba (2:1, v:v) esterilizada ARA T 6000W50L a 15 lb de presión por una hora, con 24 horas de reposo. En cada charola se colocaron 100 semillas, para un total de 300 por especie. Con base en los resultados obtenidos en esta prueba se procedió a la propagación masiva en germinadores de unicel de 288 y 242 cavidades para tlaxistle y palo dulce, respectivamente. De acuerdo con la disponibilidad, se colocaron 1,440 semillas de la primera especie y 4,840 de la segunda. En la mezcla estéril de vermiculita:turba, se sembraron las semillas y regaron a capacidad de campo con agua destilada. Los germinadores se cubrieron con plástico adherente, para conservar la humedad, hasta la emergencia de las plántulas, en promedio 15 días para palo dulce y 10 para tlaxistle. El ensayo se mantuvo en invernadero 30 días a 25°C, posteriormente se trasplantaron.
Trasplante
El tipo de contenedor se selecciona considerando la morfología radical de las plantas. Así, para tlaxistle, que presenta crecimiento monopodial, se utilizaron tubetes forestales de 12 x 4 cm, mientras que para Palo dulce, cuya raíz es polipodial ramificada, se usaron macetas semicúbicas de 5.8 x 5.2 cm. Los dos tipos de envase se desinfectaron, previamente, con hipoclorito de sodio al 5% y se enjuagaron con abundante agua destilada. En un sustrato estéril de vermiculita: turba (2:1, v:v) se trasplantaron 982 plantas de tlaxistle y 3,068 de palo dulce. La supervivencia, después del trasplante, se evaluó por conteo directo cada dos días, hasta el día 31 en Palo dulce y día 36 para tlaxistle. Al concluir esta etapa las plantas se inocularon con los hongos micorrizógenos arbusculares (HMA).
Inóculos micorrízicos
La mitad de las plantas de palo dulce (1,534) y de tlaxistle (491) se inocularon 31 días después del trasplante con una mezcla de esporas de tres especies de HMA, de acuerdo a la propuesta de Hernández-Cuevas y García-Sánchez (2008), quienes indican emplear una mezcla de 20 esporas de Glomus claroideum Schenk et Smith, 20 de Acaulospora laevis Gerdemann et Trape y 20 de A. morrowiae Spain et Schenk, es decir, un total de 60 esporas por planta. El resto de las plantas no se inocularon. Las cepas de HMA pertenecen a la colección de cultivos puros de hongos micorrizógenos arbusculares del Centro de Investigación en Ciencias Biológicas de la Universidad Autónoma de Tlaxcala, y fueron aisladas originalmente de tepetates recolectados en el municipio de Hueyotlipan, Tlaxcala.
Las plantas se mantuvieron en invernadero a 26°C, con riego a capacidad de campo cada 72 horas, durante 120 días. El diámetro basal y la altura se midieron en 100 plantas de cada especie tomadas al azar, de ellas 10 se utilizaron para obtener los datos de biomasa de las partes aérea y radical. Adicionalmente, en 15 plantas de cada taxón se determinó el contenido total de fósforo (P) y nitrógeno (N) en la parte aérea, con base en los métodos propuestos bajo las normas mexicanas NMX-Y-025-1981 y NMX-F-608-NORMEX02, respectivamente (AOAC 2010a; AOC 2010b). Con las raíces se evaluaron los porcentajes de colonización micorrízica, siguiendo el procedimiento descrito por Hernández-Cuevas et al . (2008). Se calcularon la relación biomasa aérea / biomasa radical y el índice de esbeltez, este último resulta del cociente de la altura entre el diámetro (Rodríguez, 2008). Las variables anteriores se analizaron mediante una prueba de t-student para muestras independientes, en la que se compararon plantas micorrizadas contra no micorrizadas en el programa de análisis estadístico SPSS© (SPSS, 1999).
RESULTADOS Y DISCUSIÓN
La germinación por especie finalizó a los 30 días de la siembra y su evaluación se realizó al término de dicho periodo (Cuadro 1). Los porcentajes registrados en la literatura para Eysenhardtia polystachya son muy variables, desde 28 hasta 85% (González y Camacho, 2000), por lo que la cifra obtenida en el presente trabajo (46%) se considera aceptable, pues es un valor intermedio. Cervantes y Sotelo (2002) indicaron que dicho comportamiento de los números puede responder a la procedencia de la semilla.
El tlaxistle registró un porcentaje ligeramente menor (43%), el cual es difícil calificar dado que, como ya se señaló, no existen datos documentados sobre el particular, pero de una cercana, Amelanchier alnifolia (Nutt.) Nutt. ex M. Roem., se han citado valores de germinación del 7 al 67%, indicándose que dicha variación es atribuible al origen de la semilla (Acharya et al ., 1989), tal como se cita para palo dulce.
La supervivencia al trasplante, previa a la inoculación, fue de 97% para tlaxistle y de 83% para palo dulce (Cuadro 2), cifras que se consideran bastante buenas ya que el trasplante es normalmente una fase crítica para las plantas en vivero.
González y Camacho (2000) obtuvieron una alta mortalidad de plántulas de palo dulce, resultado que contrasta con los porcentajes de supervivencia observados en este estudio, que son muy altos, y se atribuyen a que la mezcla de vermiculita:turba, en la proporción utilizada, permite mantener un pH cercano al neutro (6.2) y a que la pasteurización impidió el desarrollo de microorganismos indeseables que diezman, de manera crítica, a las plántulas durante las etapas iniciales.
Con respecto a la micorrización, fue mayor en tlaxistle que en palo dulce, 18 y 6% en promedio, respectivamente; aunque los valores de colonización en ambos son relativamente bajos, menores a 20% (Cuadro 3). En otras leguminosas leñosas inoculadas con HMA se han determinado 24% de colonización, Ac acia farnesiana (L.) Willd. (Monroy-Ata et al., 2007) y 54% en Mimosa depauperata Benth. (García-Sánchez et al., 2008). No obstante, es importante notar que en una misma especie la colonización es en general muy variable;por ejemplo, en Prosopis laevigata (Willd.) M. C. Johnst. es desde 3.5 hasta 53% (Monroy-Ata et al., 2007; García-Sánchez et al., 2008), mientras que en Cassia tomentosa (Jacq.) H. S. Irwin et Barneby, de 41 a 76% (García-Gallegos et al., 2009). Entre los factores que pueden influir en los niveles de micorrización observados están la infectividad y especificidad de los HMA, así ejemplo, en C. tomentosa las diferencias en la colonización se asociaron con el origen de los inóculos de HMA probados (García-Gallegos et al., 2009).
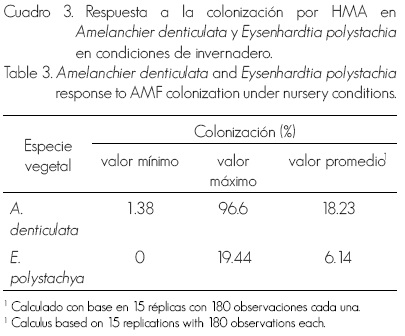
Dado que no existen datos previos para tlaxistle, es difícil calificar los valores de colonización registrados. En algunas rosáceas cultivadas del género Prunus se han consignado micorrizaciones desde menos del 10% hasta cerca del 80%, en función de las especies de hongos micorrizógenos utilizadas (Calvet et al ., 2004).
Como ya se indicó, los valores promedios de colonización fueron relativamente bajos; sin embargo se presentaron diferencias estadísticas altamente significativas (p<0.001) en diámetro, altura y biomasa aérea entre plantas con y sin inoculación micorrizica; en biomasa radical solo hubo diferencias significativas (p<0.05) entre plantas micorrizadas y no micorrizadas de Palo dulce (Cuadro 4). Con base en el tiempo de crecimiento de las plantas, estos efectos son atribuibles a la micorrización, aún cuando los porcentajes de colonización son bajos. Los resultados reafirman lo establecido por Clark et al . (1999) referente a la ausencia de una relación directa entre la infectividad y la efectividad de los HMA.
Un buen desarrollo de las plantas en sus etapas iniciales de crecimiento les confiere ventajas adaptativas, ya que al ser más vigorosas y tener sistemas radicales bien conformados serán capaces de tolerar mejor el trasplante, tal como sucedió con palo dulce. Diversos autores han expresado que un aumento en la biomasa representa mayores probabilidades de establecimiento exitoso en campo (González-Monterrubio et al ., 2005; Monroy-Ata et al ., 2007; Estrada-Luna y Davies, 2008), lo cual es crítico para especies destinadas a programas de restauración (Ferrari y Wall, 2004; Rondon y Vidal, 2005).
Una relación eficiente entre biomasa aérea y biomasa radical (BA / BR) es un indicador de la condición de la planta, y por tanto, de sus posibilidades de supervivencia, sobre todo en ambientes con limitaciones de humedad (Rodríguez, 2008). En ninguna de las dos especies estudiadas hubo diferencia estadística entre individuos con y sin micorriza para dicha relación, pero es importante notar que en el caso del tlaxistle el valor es superior en las micorrizadas, es decir, presentaron menos raíces en proporción al follaje, lo que se reconoce como una característica poco deseable para plantas cuyo final serán sitios con problemas hídricos (Rodríguez, 2008), como los suelos degradados.
En contraste, palo dulce tuvo valores para BA / BR más bajos en el tratamiento con HMA (Cuadro 4) esto indica un mayor desarrollo del sistema radical en proporción al tejido fotosintético, en consecuencia más aptitud para condiciones de poca disponibilidad de agua. Rodríguez (2008) cita que independientemente del sistema de producción e incluso de la especie, una relación menor a uno, como el observado en palo dulce, sugiere una alta probabilidad de supervivencia en sitios secos, contrario a lo que se esperaría si la proporción fuese mayor a uno, como en tlaxistle.
Otro indicador de la calidad de planta que se usa con frecuencia es el Índice de Esbeltez, el cual se refiere a las posibilidades de resistencia de la planta a factores físicos como el viento, daño mecánico, sequía, etc. A medida que su valor disminuye, los individuos tienden a ser más aptos para soportar condiciones como las antes descritas (Rodríguez, 2008). En el presente estudio tampoco hubo diferencia estadística entre plantas micorrizadas y no micorrizadas de ambas especies, pero es evidente que los mejores (más bajos) índices se observaron en palo dulce, a diferencia del tlaxistle, cuyos valores más altos se asocian con ejemplares menos robustos y por tanto más susceptibles a las situaciones adversas del ambiente (Cuadro 4).
Algo similar se observó en el contenido de fósforo (P) en la parte aérea, ya que las dos especies tuvieron registros más altos con la micorrización (Cuadro 4). El nitrógeno (N) fue superior en plantas de palo dulce sin inoculación, mientras que en tlaxistle, prácticamente, no hubo diferencia en esta variable. Lo anterior confirma el papel de las micorrizas arbusculares en el mejoramiento de la absorción de P, pero muy poco en la de N (Tarafdar y Panwar, 2008). La respuesta en Palo dulce refuerza la importancia de analizar con mayor detalle el papel de las bacterias fijadoras de nitrógeno que pueden asociarse con E. polystachya . No obstante, los resultados de los dos taxa son muy prometedores para fines de restauración, ya que en ambientes limitantes la captación de P juega un papel fundamental para su establecimiento y supervivencia en campo (Tarafdar y Panwar, 2008).
Por lo anterior, la inoculación de hongos micorrizógenos arbusculares en tlaxistle y palo dulce es recomendable, así como el estudio de otras especies de HMA, a fin de aumentar sus posibilidades de establecimiento y desarrollo, y en consecuencia, su uso en programas de restauración de suelos erosionados.
CONCLUSIONES
La propagación a partir de semilla en vermiculita y turba pasteurizadas es una buena alternativa para A. denticulata y E. polystachia , ya que muestran altos porcentajes de germinación y de supervivencia al trasplante.
Aún con porcentajes relativamente bajos de micorrización se obtuvieron mejoras significativas en el crecimiento inicial en diámetro y altura de A. denticulata y E. polystachia , lo cual puede conferirles ventajas importantes al momento del trasplante a campo.
AGRADECIMIENTOS
Los resultados de este trabajo forman parte del Proyecto "Uso de árboles y arbustos nativos micorrizados para la restauración de suelos degradados" financiado por Fondos Mixtos CONACYT-Gobierno del estado de Tlaxcala, clave FOMIX-TLAX 2004-C01-20.
REFERENCIAS
Acharya, S. N., C. B. Chu and R. Hermesh. 1989. Effects of population, environment and their interaction on Saskatoon berry ( Amelanchier alnifolia Nutt.) seed germination. Can J. Plant Sci. 69: 277-284. [ Links ]
Association of Official Agricultural Chemists AOAC. 2010a. Quinolinium molybdophosphate phosphorus (water soluble) in fertilizers. In : Horwitz, W. and G. W. Latimer Jr. (Eds.). Chapter 2. Association of Analytical Communities. Official methods of analysis of AOAC International. Gaithersburg, MD. USA. pp. 9. [ Links ]
Association of Official Agricultural Chemists AOAC. 2010b. Foods protein digestibility. In : Horwitz, W. and G. W. Latimer Jr. (Eds.). Chapter 45. Association of Analytical Communities. Official methods of analysis of AOAC International. Gaithersburg, MD. USA. pp. 79-80. [ Links ]
Bashan, Y., M. E. Puente, B. Salazar, L. E. de Bashan, M. Bacilio, J. P. Hernández, L. A. Leyva, B. Romero, R. Villalpando y G. Bethlenfalvay. 2005. Reforestación de tierras erosionadas en el desierto: el papel de las bacterias promotoras de crecimiento en plantas y la materia orgánica. Suelos Ecuat. 35(1): 70-77. [ Links ]
Calvet, C., V. Estaún, A. Camprubi, A. Hernández-Dorrego, J. Pinochet and M. A. Moreno. 2004. Aptitude for mycorrhizal root colonization in Prunus rootstocks. Scientia Hort. 100(1-4): 39-49. [ Links ]
Cervantes S., M. A. y M. E. Sotelo. 2002. Guías técnicas para la propagación sexual de 10 especies latifoliadas de Selva Baja Caducifolia en el estado de Morelos. CIR-Centro, SAGARPA-INIFAP. Cuernavaca, Mor. México. 30 p. [ Links ]
Challenger, A. y J. Soberón. 2008. Los ecosistemas terrestres. In : Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (Ed.). Capital Natural de México, vol. I: Conocimiento actual de la biodiversidad. México, D. F. México. pp. 87-108. [ Links ]
Challenger, A., R. Dirzo, J. C. López, E. Mendoza, A. Lira-Noriega e I. Cruz. 2009. Factores de cambio y estado de la biodiversidad. In : Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (Ed.). Capital Natural de México, Vol. II: Estado de conservación y tendencias de cambio. México, D. F. México. pp. 37-73. [ Links ]
Clark, R. B., S. K. Zeto and R. W. Zobel. 1999. Arbuscular mycorrhizal fungal isolate effectiveness on growth and root colonization of Panicum virgatum in acidic soil. Soil Biol. Biochem. 31(13): 1757-1763. [ Links ]
Crisóstomo, M. S., G. Gómez-Cruz, R. Ferrera-Cerrato, R. Quintero-Lizaola, y J. A. Santizo-Rincón. 1991. La influencia de los hongos endomicorrízicos V-A en la disponibilidad del fósforo en Eysenhardtia polystachya . In : Tovar S., J. L. y R. Quintero-Lizaola (Eds.) La investigación edafológica en México. Memorias XXIV Congreso Nacional de la Sociedad Mexicana de la Ciencia del Suelo, Pachuca, Hgo. México. p. 108. [ Links ]
Estrada-Luna, A. A. y F. T. Davies Jr. 2008. Estado nutrimental y crecimiento de plantas micropropagadas de nopal ( Opuntia albicarpa Scheinvar cv. "Reyna") colonizadas con tres cepas seleccionadas de endomicorrizas. In : Montaño-Arias, N. M., S. L. Camargo-Ricalde, R. García-Sánchez y A. Monroy-Ata (Eds.): Micorrizas arbusculares en ecosistemas áridos y semiáridos. Mundi-Prensa S.A. de C.V., Instituto Nacional de Ecología, SEMARNAT, UAM-Iztapalapa. FES-Zaragoza-UNAM, México, D. F. México. pp. 203-215. [ Links ]
Ferrari, A. E. y L. G. Wall. 2004. Utilización de árboles fijadores de nitrógeno para la revegetación de suelos degradados. Rev. Fac. Agron. La Plata 105(2): 63-87. [ Links ]
Flores O., M. H. y R. Lindig-Cisneros. 2005. La lista de nombres vulgares y botánicos de árboles y arbustos propicios para repoblar los bosques de la República de Fernando Altamirano y José Ramírez a más de 100 años de su publicación. Rev. Mex. Biod. 76: 11-35. [ Links ]
García-Gallegos, E., G. Gómez, O. G. Vázquez y E. M. Zamora. 2009. Respuesta de Cassia tomentosa desarrollada en tepetate con inoculación micorrízica bajo condiciones de invernadero. Revista UDO Agrícola 9(4): 816-825. [ Links ]
García-Oliva, F. 2005. Algunas bases del enfoque ecosistémico para restauración. In : Sánchez, O., E. Peters, R. Márquez-Huitzil, E. Vega, G. Portales, M. Valdez y D. Azuara (Eds.). Temas sobre restauración ecológica. SEMARNAT-INE-U.S. Fish and Wildlife Service-Unidos para la Conservación A.C., México, D. F. México. pp. 101-112. [ Links ]
García-Sánchez, R., A. Monroy-Ata y E. Chimal. 2008. Hongos micorrizógenos arbusculares asociados a diferentes plantas y matorrales del Valle del Mezquital, Hidalgo, México. In : Montaño-Arias, N. M., S. L. Camargo-Ricalde, R. García-Sánchez y A. Monroy-Ata (Eds.): Micorrizas arbusculares en ecosistemas áridos y semiáridos. Mundi-Prensa S.A. de C.V., Instituto Nacional de Ecología, SEMARNAT, UAM-Iztapalapa. FES-Zaragoza-UNAM, México, D. F. México. pp. 123-136. [ Links ]
González K., V. and F. Camacho M. 2000. Test on growing media for Eysenhardtia polystachya , a promissing species for planting on degraded areas of Mexico. Seed. Sci. Tech. 28(2): 271-275. [ Links ]
González R., H. e I. Cantú. 2001. Adaptación a la sequía de plantas arbustivas de matorral espinoso tamaulipeco. Ciencia UANL 4(4): 454-461. [ Links ]
González-Monterrubio, C., A. Monroy-Ata, E. M. García-Amador y M. S. Orozco-Almanza. 2005. Influencia de hongos micorrizógenos arbusculares (HMA) en el desarrollo de plántulas de Opuntia streptacantha Lem. sometidas a sequía, en condiciones de invernadero. Rev. Esp. Ciencias Quím. Biól. 8(1): 5-10. [ Links ]
Haulon, M., G. Flores, A. Vera, G. Werner y G. Velásquez. 2005. Rehabilitación de tepetates y medición de erosión en Santiago Tlalpan. In : Flores, G., E. Fuentes, N. García, M. Haulon y A. Vera (Comps.). La edafología y la sostenibilidad. Guía de campo. XXIII Curso-Diplomado Internacional de Edafología "Nicolás Aguilera". Universidad Autónoma de Tlaxcala, Tlax. México. pp. 65-73. [ Links ]
Hernández-Cuevas, L. y R. García-Sánchez. 2008. Propagación por esporas: el caso de las micorrizas arbusculares In : Álvarez-Sánchez, J. y A. Monroy-Ata (Comps.): Técnicas de estudio de las asociaciones micorrízicas y sus implicaciones en la restauración. Las Prensas de Ciencias-UNAM, México, D. F. México. pp. 29-42. [ Links ]
Hernández-Cuevas, L., P. Guadarrama-Chávez, I. Sánchez-Gallén y J. A. Ramos-Zapata. 2008. Micorriza arbuscular. Colonización intrarradical y extracción de esporas del suelo. In : Álvarez-Sánchez, J. y A. Monroy-Ata (Comps.). Técnicas de estudio de las asociaciones micorrízicas y sus implicaciones en la restauración. Las Prensas de Ciencias-UNAM, México, D. F. México. pp. 1-16. [ Links ]
Magdaleno, M. L., E. García, J. I. Valdez-Hernández y V. De la Cruz. 2005. Evaluación del sistema agroforestal "árboles en terrenos de cultivo" en Vicente Guerrero, Tlaxcala, México. Rev. Fitotec. Mex. 28 (3): 203-212. [ Links ]
Monroy-Ata, A., J. Estevez, R. García-Sánchez y R. Ríos. 2007. Establecimiento de plantas mediante el uso de micorrizas y de islas de recursos en un matorral xerófilo deteriorado. Bol. Soc. Bot. Mex. 80 (Supl.): 49-57. [ Links ]
Montaño, N. M. y A. Monroy. 2000. Alternativas para la conservación ecológica de suelos en zonas áridas y semiáridas de México. Ciencia y Desarrollo 154: 26-37. [ Links ]
Morfin L., L. y F. Camacho M. 1987. El Palo dulce ( Eysenhardtia polystachya (Ortega) Sarg.) una alternativa para la explotación forrajera de áreas tepetatosas. In : Ruíz, F. F. (Ed.). Uso y manejo de tepetates para el desarrollo rural. Departamento de Suelos. Universidad Autónoma Chapingo, Texcoco, Edo. de México. México. pp. 192-198. [ Links ]
Programa Nacional de Reforestación PRONARE. 2000. Evaluación de la reforestación 1999 del Programa Nacional de Reforestación, SEMARNAP. México, D.F. México. s/p. [ Links ]
Rodríguez T., D. A. 2008. Indicadores de Calidad de Planta Forestal. Universidad Autónoma Chapingo-Mundi-Prensa, S.A. de C.V., México, D. F. México. 156 p. [ Links ]
Rondón, J. A. y R. Vidal. 2005. Establecimiento de la cubierta vegetal en áreas degradadas (principios y métodos). Rev. For. Lat. 38: 63-82. [ Links ]
Statistical Package for the Social Sciences.SPSS 1999. Base 10.0. Applications Guide. SPSS Inc. Chicago, IL. USA. 426 p. [ Links ]
Tarafdar, J. C. y J. Panwar. 2008. El papel de las micorrizas arbusculares en la nutrición por P en plantas de ambientes áridos y semiáridos. In : Montaño-Arias, N. M., S. L. Camargo-Ricalde, R. García-Sánchez y A. Monroy-Ata (Eds.): Micorrizas arbusculares en ecosistemas áridos y semiáridos. Mundi-Prensa S.A. de C.V., Instituto Nacional de Ecología, SEMARNAT, UAM-Iztapalapa. FES-Zaragoza-UNAM, México, D. F. México. pp. 41-53. [ Links ]
Terrones, R. T. del R. L., H. García, M. A. Hernández y C. Mejia, 2007. Potencial agroforestal con arbustivas nativas. Folleto técnico 1. INIFAP-Campo Experimental Bajío. Celaya, Gto. México. 36 p. [ Links ]
Varela, L. 2000. Estudio de la micorriza y hongos micorrizógenos en un matorral secundario establecido en el Cerro de Tepeticpac, Tlaxcala, México. Tesis de Doctorado, ENCB, IPN, México, D. F. México. 86 p. [ Links ]
Werner, G. 1992. Suelos volcánicos endurecidos (tepetates) en el estado de Tlaxcala: distribución, rehabilitación, manejo y conservación. Terra 10: 318-331. [ Links ]

