










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista mexicana de ciencias forestales
versão impressa ISSN 2007-1132
Rev. mex. de cienc. forestales vol.2 no.3 México Jan./Fev. 2011
Artículos
Análisis morfológico y de hábitat en el híbrido Quercus glabrescens x Q. rugosa
Morphologic and habitat analysis of Quercus glabrescens x Q. rugosa HYBRID
Sandra Marisol Núñez-Castillo1, José Guadalupe Álvarez-Moctezuma1, Fernando Zavala-Chávez1† y Policarpo Espinosa-Robles2
1 División de Ciencias Forestales. Universidad Autónoma Chapingo. Correo-e: jogualmo@correo.chapingo.mx
2 Departamento de Fitotecnia. Universidad Autónoma Chapingo.
Fecha de recepción: 25 de febrero de 2010.
Fecha de aceptación: 15 de febrero de 2011.
RESUMEN
Las especies del género Quercus tienden a formar híbridos con alta frecuencia en diversas partes del mundo. Se estudió el híbrido Quercus glabrescens x Q. rugosa , en función del hábitat (distribución geográfica y asociación de especies) y del índice de hibridación morfológico. Con la distribución de Q. glabrescens y Q. rugosa se obtuvo un modelo bioclimático de temperatura mínima media y porcentaje de precipitación invernal. En el Parque Nacional El Chico, Hidalgo, México se levantaron ocho parcelas y se calculó la frecuencia, cobertura, área basal, volumen del fuste e índice de dominancia de las especies arbóreas. Se valoraron 10 caracteres morfológicos en 50 individuos (supuestamente híbridos). El modelo bioclimático mostró que Q. glabrescens tiene mayor preferencia por zonas con temperaturas mínimas medias menores y porcentaje de precipitación invernal más alto que Q. rugosa . Se identificó una zona común de afinidades ecológicas. A través del índice de correlación de Pearson y del Análisis de Componentes Principales se detectaron variantes de hábitat con la asociación de especies y categoría de híbrido. Se presentaron individuos con tendencia a Q. glabrescens e intermedios. Los primeros manifestaron afinidades ecológicas similares a Q. glabrescens y los segundos expresaron flexibilidad, sobre el particular. Se aporta evidencia de la hibridación natural entre Q. glabrescens y Q. rugosa .
Palabras clave: Análisis de Componentes Principales, hábitat, hibridación natural, índice de hibridación morfológico, Quercus glabrescens, Quercus rugosa.
ABSTRACT
Quercus species are prone to form hybrids very frequently in different parts of the world. The Quercus glabrescens x Q. rugosa hybrid was studied in terms of habitat (geographic distribution and species association) and of the Morphologic Hybridization Index. With the distribution of Q. glabrescens and Q. rugosa was obtained a bioclimatic model of minimum average temperature and winter rainfall per cent. At El Chico National Park, Hidalgo State, Mexico, eight lots were surveyed and in each of them was determined frequency, cover, basal area, stem volume and dominance index of the tree species. Ten morphologic characters were assessed in 50 examples (alleged hybrids). The bioclimatic model showed that Q. glabrescens has a greater preference for zones where low minimum average temperatures prevail and with winter rainfall per cent higher than Q. rugosa . A common zone with ecologic affinities was found Through Pearson correlation and the Principal Component Analysis, diverse habitats were detected with species association and hybrid category. Trees with a tendency for Q. glabrescens or in-between were found. The first showed ecologic affinities similar to Q. glabrescens and the second expressed flexibility. Evidence is reported about the natural hybridization between Q. glabrescens and Q. rugosa.
Key words: Principal Component Analysis, habitat, natural hybridization, morphology hybridization index, Quercus glabrescens, Quercus rugosa.
INTRODUCCIÓN
El género Quercus L. cuenta con más de 500 especies distribuidas en el mundo y es característico de zonas templadas. Para México se reconocen entre 135 y 150, de las cuales 86 son endémicas (Nixon, 1993). Este taxón junto con Pinus cubren la mayor parte del territorio con clima templado y semihúmedo del país (Rzedowski, 1978).
La hibridación natural es un fenómeno frecuente entre plantas (Rieseberg, 1997). Consiste en el cruzamiento de diferentes taxa separados por aislamiento ecológico o reproductivo (Grant, 1989; Rieseberg y Carney, 1998). En la literatura se cita a Quercus con una tendencia a formar híbridos (Muller, 1952; Grant, 1989; Arnold, 1997), para Europa con una frecuencia del 31.7% (Bacilieri et al ., 1996) y en México no son la excepción (González-Rodríguez et al. , 2004; Tovar-Sánchez y Oyama, 2004). La hibridación en los encinos ha sido detectada a través de características morfológicas y marcadores moleculares por diversos autores (Tucker, 1968; Bruschi et al ., 2003; González-Rodríguez et al. , 2004; Tovar-Sánchez y Oyama, 2004; González-Rodríguez y Oyama, 2005; Albarrán-Lara et al., 2010).
Quercus glabrescens Benth. y Quercus rugosa Née tienen elementos contrastantes en su morfología foliar (Cuadro 1). Sin embargo, se ha observado que algunos individuos tienen hojas y corteza con rasgos intermedios entre ambos. Q. rugosa se considera como el taxón del subgénero Leucobalanus que presenta hibridación con un mayor número de especies (Zavala, 1995).
El índice morfológico de hibridación es una herramienta útil para determinar los híbridos (Long, 1959; Keim et al ., 1989; Cruzan y Arnold, 1993; Tovar-Sánchez y Oyama, 2004, 2006). Así mismo, se reconoce la importancia del hábitat en la definición de los taxa (Van Valen, 1976) y la presión de selección que ejerce sobre el fenotipo, no en el genotipo. Por lo tanto, es imposible entender la historia evolutiva de una especie si éste es excluido del análisis, por lo que es indispensable incluirlo en los estudios de los procesos de especiación e hibridación; en lo que corresponde a las dos especies mexicanas mencionadas, hay pocas investigaciones. Álvarez-Moctezuma et al . (1998) lograron diferenciar hábitats para cinco especies de Quercus con las variables altitud y precipitación invernal (de noviembre a abril). Generalmente a Q. rugosa se le observa desde zonas húmedas hasta regiones secas, con una amplia distribución (Rzedowski, 1978; Zavala, 1995), mientras que Q. glabrescens sólo se desarrolla en regiones húmedas o subhúmedas de la Faja Volcánica Transmexicana (Zavala, 1995) (Figura 1).
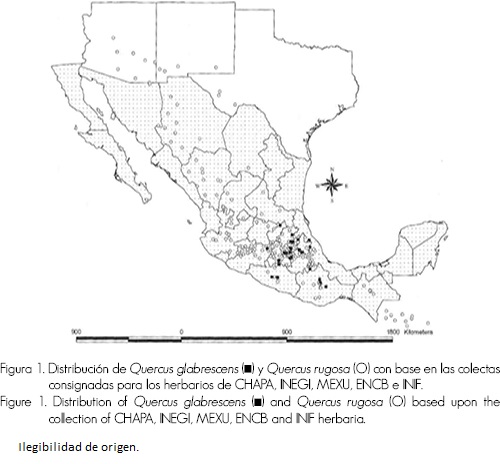
La alteración de los hábitats favorece el rompimiento de barreras reproductivas, al aumentar la frecuencia de híbridos, que en función de sus capacidades, podrán ocupar hábitats similares, intermedios o distintos a los parentales (Anderson, 1948; Cruzan y Arnold, 1993; Rieseberg y Carney, 1998).
La hibridación propicia la diversidad del acervo genético al originar genotipos nuevos en las poblaciones, y más aún cuando los híbridos logran establecerse y reproducirse, con la posibilidad de producir poblaciones introgresivas (Mettler y Gregg, 1982; Grant, 1989). La gran variación de fenotipos en tales individuos da lugar a problemas de nivel taxonómico, particularmente en Quercus , cuya complejidad taxonómica se incrementa con la hibridación (Zavala, 1990; Valencia, 2004).
Quercus glabrescens se distribuye en el bosque mesófilo de montaña, bosque de Pinus - Quercus y en el bosque de Abies-Quercus (Cuadro 2), entre los 2,500 y los 3,200 msnm, en suelos profundos (Zavala, 1995; Mejía et al ., 2004; Martínez et al ., 2004; CONANP, 2004). Q. rugosa es tolerante a muchos ambientes, en consecuencia, tiene una distribución amplia (Figura 1) y se asocia con un gran número de especies. Puede crecer en terrenos planos, laderas, cañadas, matorrales subtropicales hasta bosque mesófilo de montaña (Bello y Labat, 1987). Generalmente, se relaciona con el bosque de Pinus - Quercus , bosque de Quercus-Pinus , bosque de Abies , bosque mesófilo de montaña y bosque de Pseudotsuga , entre 1,000 y 3,200 msnm (Cuadro 2), (Bello y Labat, 1987; Zavala, 1995); en suelos de someros a profundos (Valencia, 1989). Q. rugosa se distribuye en el sur de Estados Unidos, México, Guatemala y El Salvador (Figura 1)(Lundell, 1955; Zavala, 1995; Álvarez-Moctezuma et al ., 1998; Fernández-Nava et al ., 1998; Ramírez-Marcial et al ., 1998; CONABIO, 2000; Cornejo-Tenorio et al ., 2003; García-Arévalo et al ., 2004; Mejía et al ., 2004; Martínez et al ., 2004; Standley y Steyermark, 1954; CONANP, 2004). Por otro lado, Zavala (1995) menciona que la presencia de Quercus rugosa en el bosque de Abies religiosa (HBK.) Schltdl. et Cham. está relacionada con el disturbio de dicha comunidad forestal.
El cruzamiento entre Q. glabrescens y Q. rugosa es favorecido por la similitud de sus afinidades ecológicas y por la coincidencia de sus periodos de floración; además de que los encinos sólo presentan hibridación con individuos del mismo subgénero. La distribución de Q. glabrescens y Q. rugosa se traslapa en el Eje Neovolcánico, región que recientemente fue identificada como una zona híbrida entre Q. crassifolia Humb. et Bonpl. y Q. crassipes Humb. et Bonpl. (Tovar-Sánchez y Oyama 2004, 2006). También ahí se localiza el Parque Nacional El Chico, Hidalgo, en donde se observa que la floración masculina de Q. glabrescens es más temprana en relación a la de Q. rugosa que florece hasta abril (la antesis ocurre en marzo).
En recorridos realizados en El Chico se han identificado árboles con características adecuadas para la producción forestal (rectitud del fuste, gran diámetro, altura, autopoda) y con rasgos intermedios entre Quercus rugosa y Q. glabrescens.
El objetivo de este trabajo fue caracterizar el hábitat y evaluar la hibridación en ejemplares de Q. glabrescens x Q. rugosa, previamente registrados en el Parque Nacional El Chico.
MATERIALES Y MÉTODOS
El estudio del hábitat se abordó por medio de datos de distribución geográfica y asociación de especies. Para la primera se consultaron ejemplares de Q. glabrescens y Q. rugosa del Herbario de la División de Ciencias Forestales de la Universidad Autónoma Chapingo (CHAPA), Herbario del Instituto de Estadística, Geografía e Informática (INEGI), Herbario Nacional (MEXU) del Instituto de Biología de la Universidad Nacional Autónoma de México, Herbario de la Escuela Nacional de Ciencias Biológicas del Instituto Politécnico Nacional (ENCB) y Herbario Nacional Forestal del Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIF). Se generó un modelo bioclimático con las variables temperatura mínima media (tmm) y porcentaje de precipitación invernal (ppinv), a partir de la importancia que ambos factores tienen en la definición del hábitat de Quercus (Muller, 1952), lo que coincide con el de muchas especies forestales (USDA, 2003).
Para el modelo bioclimático se utilizó la información contenida en los ejemplares de herbario ubicados en el Eje Neovolcánico. La determinación de la temperatura mínima media requirió del dato correspondiente a la altitud del lugar de colecta; para los ejemplares que carecían de dicha información se calculó mediante cartas topográficas de INEGI 1:50,000 (INEGI, 1984). Posteriormente, se localizaron las dos estaciones meteorológicas más cercanas a esos sitios de colecta mediante el Sistema de Información Climatológica (SICLIM) versión 1.0. (1990), y se obtuvieron los valores de temperatura mínima media y precipitación invernal (diciembre, enero y febrero, que en la zona de estudio corresponde a la temporada de sequía). Se incorporó todo el periodo disponible en la base de datos del programa. Además se ajustaron las temperaturas de acuerdo a la diferencia de altitud del sitio de colecta y de las estaciones meteorológicas. Con éstas y las registradas en las dos estaciones meteorológicas se generaron gradientes regionales estimados a partir de regresiones lineales. Por último, se elaboró un gráfico con la temperatura mínima media (eje de las abscisas) y la precipitación invernal (eje de las ordenadas).
Para documentar la asociación de las dos especies se hicieron diez recorridos de campo en el Parque Nacional El Chico, Hidalgo, en donde se identificaron presuntos híbridos entre Q. glabrescens y Q. rugosa con base en la corteza y morfología foliar (figuras 2 y 3). Una vez delimitada la zona de estudio, que correspondió a la parte del bosque de Abies-Quercus (Zavala, 1995), se dividió en ocho parcelas (20 m x 20 m). Cada una incluyó al menos un supuesto individuo híbrido. Tres se ubicaron en el paraje La Peña del Cuervo, dos en El Puente de Dios, dos en La Peñita y uno colindante al puente de La Peñita. Las coordenadas de El Chico son 20° 12' 26" de latitud norte y 98° 43' 52" de longitud oeste.


En la evaluación se consideraron todos los individuos arbóreos en pie mayores a 2 m de altura, a los cuales se les registraron los siguientes datos: especie, diámetro a la altura del pecho, altura total y diámetro de copa. Además se delimitó y fotografió una superficie de 20 cm por 20 cm en la corteza (generalmente arriba de 1.30 m) de los dos taxa estudiados y del supuesto híbrido Q. glabrescens x rugosa . Se obtuvieron 50 ejemplares, aparentemente, híbridos. De cada individuo se colectaron tres muestras, una se depositó en la colección del herbario CHAPA y dos se usaron para determinar las variables morfológicas y el índice de hibridación.
Con los datos dasométricos se calcularon: la frecuencia, diámetro de fuste y de copa (se tomaron dos mediciones: una de norte a sur y otra de este a oeste); área basal [Ab = π*(diámetro de fuste/ 2)2]; cobertura de copa [Cc = π*(diámetro mayor + diámetro menor/ 2)2]; volumen de fuste (Vf = Ab*altura) e índice de dominancia [Id=(Ab + Vf + Cc relativos)/ 3].
Los factores abióticos considerados fueron: pedregosidad superficial (%), número de días con neblina, exposición (grados), pendiente (%), cobertura arbórea (de todas las especies en porcentaje) y variables edáficas que se obtuvieron de un análisis del suelo. Para este proceso se tomaron cinco submuestras (una en el centro y las otras cuatro en los vértices de cada parcela, que se mezclaron); se tomaron en cuenta las siguientes variables con sus respectivos métodos: pH (potenciómetro relación suelo: agua 1:2), porcentaje de materia orgánica (Walkley y Black, 1934), nitrógeno (extraído con cloruro de potasio 2N y determinado por arrastre de vapor), fósforo (Olsen et al ., 1954), potasio (en acetato de amonio 1N, pH 7.0, relación 1:20 y determinado por espectrofotometría de emisión de flama), calcio y magnesio extraídos en acetato de amonio 1.0N, pH 7.0, relación 1:20 y determinado por espectrofotometría de absorción atómica; hierro, cobre, zinc y manganeso extraídos por DTPA, relación 1:4 y determinado por espectrofotometría de absorción atómica; boro extraído con CaCl2 1.0 M por fotocolorimetría de azometina-H; densidad aparente (método de la probeta) y textura (hidrómetro de Bouyoucos) (SEMARNAT, 2000).
En Índice de Hibridación Morfológico (IHM) se estimó en función de 10 caracteres contrastantes entre Q. rugosa y Q. glabrescens (Cuadro 1). Se asignaron valores de cero a las características exhibidas por Q. rugosa ; dos a las de Q. glabrescens , y a las intermedias en los híbridos se les dió el valor de uno (Grant, 1989). Los caracteres fueron:
- Corteza. En un área de 20 x 20 cm por arriba de 1.30 m del fuste, se midieron las escamas en largo y ancho y se calculó la relación entre ambos
- Longitud de la lámina foliar (cm). Se eligieron cinco de las hojas más grandes de cada árbol y se midió la longitud máxima de la lámina foliar
- Longitud del pecíolo (mm). Se midió de la base de la hoja a la ramilla
- Número de nervaduras. A partir de la nervadura principal se contaron las secundarias de ambos lados y se obtuvo un número promedio
- Pubescencia. A la ausencia de pubescencia se le adjudicó el valor de cero, a la pubescencia densa de dos, cuando fue escasa, o bien en las nervaduras principales, de uno
- Margen. Al margen revoluto se le asignó el valor de cero, al plano de dos y al intermedio de uno
- Número de dientes. Se contaron por cada lado de la hoja y se calculó el promedio
- Profundidad de la escotadura (mm). Se midió la escotadura del diente más grande, por cada lado de la hoja y se promedió
- Ancho de la hoja (mm) y el ángulo del ápice de la hoja en grados. Se tomó como eje la nervadura principal y con ayuda de un transportador de 180° se determinó el ángulo del ápice.
Para evaluar el IHM se estudiaron las variables morfológicas contrastantes de acuerdo con Zavala (1989), (Cuadro 1). Con base en el IHM se definieron cinco categorías de híbrido: 0 a 0.4, 0.4 a 0.8, 0.8 a 1.2, 1.2 a 1.6, 1.6 a 2.
Los datos se trabajaron mediante una correlación momento-producto de Pearson, el índice de similitud de Horn y el análisis de componentes principales (Krebs, 1989) con el programa SAS System versión 8 (SAS, 1985).
RESULTADOS Y DISCUSIÓN
Hábitat según la distribución geográfica
Con base en la información obtenida a partir de ejemplares herborizados y el modelo bioclimático se determinó que Q. glabrescens crece en regiones con temperatura mínima media (tmm) de -11°C a 3.4°C y un porcentaje de precipitación invernal (ppinv) entre 2 y 9%. Q. rugosa se ubicó en una tmm de -11°C a 8°C con ppinv de 1.6 a 6%. Además, se presentó una amplia zona común con tmm de -11°C a 0.3°C y un ppinv de 1.7 a 5.6%, que indica afinidad ecológica entre ambas. La mayor parte de los sitios de colecta se situaron dentro de la zona común, es decir, los dos taxa coexisten en localidades con los mismos intervalos de temperatura y precipitación. También pueden apreciarse áreas bien definidas de hábitats extremos en las dos especies (Figura 4). Lo anterior denota que Q. glabrescens ocupa zonas con precipitación invernal mayor y temperaturas mínimas medias más bajas, hecho que sugiere requerimientos de ambientes más húmedos y frescos, en relación a los propios de Q. rugosa .

Los resultados aquí consignados son similares a los registrados por Álvarez-Moctezuma et al . (1998), quienes definieron a las variables altitud y precipitación invernal como determinantes en la separación del hábitat de Quercus spp. en la Meseta Central de Chiapas.
Los híbridos exhibieron alta heterogeneidad en la fecha de antesis (desde marzo hasta junio), razón por la cual es posible que algunos individuos híbridos tuvieran mayor probabilidad de cruzarse con una de las especies parentales y otros entre ellos mismos.
Índice de hibridación morfológico
Conforme al IHM se identificaron seis individuos con más similitud a Q. glabrescens, valores de 1.6, 1.7 y 1.8; once con características intermedias entre Q. glabrescens y Q. rugosa correspondientes a valores de 0.8, 0.9, 1.0, 1.1 y 1.2, y sólo un ejemplar mostró características más cercanas a Q. rugosa , con un IHM de 0.7 (figuras 2 y 3).
Se determinó que la mayoría de los supuestos híbridos son fenotípicamente intermedios entre las especies parentales y otros tienen marcada tendencia unidireccional hacia Q. glabrescens . Lo anterior se explica por el hecho de que este tipo de organismos pueden expresar un mosaico de fenotipos parentales que depende de la interacción entre la expresión genética y el ambiente (Rieseberg y Carney, 1998). Así mismo, no fue posible identificar la generación a la que pertenecen los híbridos. En trabajos previos se ha mencionado que los de la generación F1 presentan características intermedias a los padres, pero que al retrocruzarse adquieren, en las generaciones subsecuentes, mayor número de características de uno de ellos, ya que su frecuencia y permanencia por afinidades ecológicas se potencializa (Anderson, 1948; Potts y Reid, 1985; Grant, 1989; Cruzan y Arnold, 1993 y Rieseberg y Carney, 1998).
La evidencia de hibridación mediante el IHM ha sido confirmada en Helianthus maximilianii Schrad. x H. grosseserratus Martens (Long, 1959), Eucalyptus risdonii Hook x E. amygdalina Labill., Populus fremontii S. Wats. x P. angustifolia James (Keim et al ., 1989), Iris fulva x I. brevicaulis (Cruzan y Arnold, 1993) y Q. crassifolia x Q. crassipes (Tovar-Sánchez y Oyama, 2004; Tovar-Sánchez y Oyama 2006), especies en las que se observó variación en el grado de hibridación de acuerdo con la forma de la hoja, supervivencia, vigor y adaptación en plántulas de retrocruzas.
El IHM promedio por parcela osciló entre 0.9 y 1.5, con una media aritmética de 1.23 (desviación estándar de 0.19). La presencia de los híbridos (en función de la cobertura arbórea) fue variable. En la parcela uno se registraron las categorías de híbridos con un IHM de 1.2 - 1.6, 0.8 - 1.2 y 0.4 - 0.8, y la 1.2 - 1.6 dominó en función del área basal, cobertura, volumen de fuste e índice de dominancia. La parcela tres reunió individuos de las categorías 1.2 - 1.6 y 0.8 - 1.2 y sobresalió la 0.8 - 1.2 en área basal, cobertura, volumen e índice de dominancia. En las parcelas cuatro y cinco se definieron las categorías 1.6 - 2 y 1.2 - 1.6; en ambas predominó la 1.2 - 1.6 en frecuencia, área basal, cobertura, volumen e índice de dominancia. Por último, en las parcelas dos, seis, siete y ocho se registraron las categorías 1.2 - 1.6 y 0.8 - 1.2, y en todas las parcelas predominó la categoría 1.2 - 1.6 (Figura 5).
Hábitat en función de la asociación de especies
El análisis de similitud entre parcelas mostró la existencia de tres grupos. De acuerdo con el índice de similitud de Horn (Krebs, 1989), el primero está formado por las parcelas uno, cinco y siete (unidas en un nivel de similitud de 0.63); el segundo por las seis, ocho y dos y el tercero por la tres y la cuatro (Figura 6). Estos resultados concuerdan con el análisis de componentes principales (Krebs, 1989), que organizó agrupamientos parecidos, a excepción del grupo uno, al que se sumó la parcela dos. Dicho patrón sugiere una cierta posibilidad de indicar hábitats variables en las parcelas (Figura 7).


En estudios florísticos, la similitud entre parcelas ha sido abordada con el coeficiente de Jaccard, que genera grupos con valores semejantes a los obtenidos en la presente investigación (José-Noguera et al., 2006). En el caso actual se establecieron dos grupos con coeficientes de 0.70 y 0.32, evidencia de la heterogeneidad de la flora entre las parcelas (Catalán-Heverástico et al ., 2003).
En el presente estudio se documentó que los supuestos híbridos se asocian (en todas las parcelas muestreadas) con Arbutus xalapensis Kunth, Garrya laurifolia Hartw. ex Benth., Juniperus monticola Mart., Prunus serotina Ehrh. , Ribes affine Kunth, Abies religiosa, Quercus laurina Bonpl. , Quercus crassifolia Humb. et Bonpl., Arctostaphylos arguta (Zucc.) DC. y Cercocarpus macrophyllus C. Schneid. En la parcela uno, 15% de la vegetación arbórea la domina Abies , y en la siete representa 48%; en las parcelas dos, cuatro, cinco, seis y ocho el género Quercus predominó con 66%, 54%, 67%, 46% y 70%, respectivamente, y en la tres sobresalió Juniperus monticola con 67% (Figura 8). La explicación de estos resultados es que la dominancia corresponde a un atributo de la comunidad, muy diferente a la riqueza florística.
La asociación de variables dependientes (categorías de IHM) e independientes (especies asociadas, características edafológicas, pedregosidad, pendiente, exposición y neblina), a través del análisis de correlación de Pearson (Krebs, 1989), mostró que la categoría de híbrido con IHM 1.6 - 2 está fuertemente correlacionada de manera positiva con Abies religiosa , con la pendiente del terreno y con la exposición (Cuadro 3). Puede asumirse que los híbridos de la categoría 1.6 - 2 de IHM tienen las mismas afinidades ecológicas que Abies religiosa . Por otra parte, en el área de estudio la pendiente del terreno varía de ligera hasta altamente pronunciada, con cañadas protegidas del viento y exposición hacia las laderas húmedas.
La categoría de híbrido con un IHM de 1.2 - 1.6 se asocia de manera positiva con Ribes affine, Arbutus xalapensis y Quercus crassifolia . Con respecto a las variables abióticas, parece ser que tiene afinidades ecológicas más específicas, ya que mostró una fuerte correlación positiva con nitrógeno inorgánico, zinc, manganeso y materia orgánica (Cuadro 3). Las correlaciones obtenidas evidencian preferencias edáficas de suelos húmedos, ricos en nutrimentos y materia orgánica. De manera coincidente, el manganeso es un elemento presente en suelos constantemente inundados (Ortega, 1981).
La categoría de híbrido con un IHM de 0.8 - 1.2 tiende a desarrollarse junto a Juniperus monticola, Abies religiosa, Garrya laurifolia y Quercus crassifolia , sobre suelos someros. Se le localiza en la parte intermedia de las afinidades del hábitat de los parentales ( Q. glabrescens y Q. rugosa ), lo que hace suponer que requiere condiciones intermedias para su establecimiento, al asociarse con especies que se ubican tanto en regiones húmedas, como secas (Cuadro 3). Este fenómeno se ha descrito para otros híbridos (Anderson, 1948; Grant, 1989). Sin embargo, esto no es general para todos; por ejemplo, en híbridos entre Iris hexagona Walt. x I. fulva Ker Gawl. (con tendencia bidireccional) se observó preferencia por hábitats intermedios, ya que ocupan áreas ecotonales entre los ámbitos de los taxa parentales (Arnold, 1992).
El análisis de componentes principales diferenció dos grupos de híbridos (Figura 9). El primero incluyó aquéllos con IHM de 1.6 - 2 y 1.2 - 1.6 (tendencia hacia Q. glabrescens ) relacionado con las variables manganeso, materia orgánica y nitrógeno inorgánico. El segundo conjunto correspondió a las de IHM de 0.8 - 1.2 y 0.4 - 0.8 (características intermedias e inclinación hacia Q. rugosa ); y se asoció con la cobertura arbórea de Abies religiosa y Arbutus xalapensis y con la cobertura absoluta de todas las especies. Esto coincide con el estudio de hibridación natural entre Nothofagus antarctica (G. Foster) Oerst. y N. pumilio (Poepp et Endl.) Krasser. en el que uno de los grupos segregó a N. pumilio y el otro integró a los individuos híbridos con N. antarctica (Quiroga et al ., 2005). A diferencia del análisis de correlación, con el de componentes principales se logró asociar la categoría de IHM 0.4 - 0.8 con otras variables. No obstante, en ambos métodos no existió ningún tipo de asociación para la categoría 0 - 0.4, por falta de datos.

Los dos análisis concordaron en los resultados, el análisis de correlación agrupó características entre la categoría de híbrido y las variables ecológicas que permiten profundizar en cuanto a las preferencias de hábitat de los híbridos. El análisis de componentes principales separó las necesidades entre las diferentes categorías de híbridos. Así, los híbridos con tendencia a Q. glabrescens demandan hábitats con características muy particulares, como la especie parental; y las de fenotipos intermedios tienen mayor flexibilidad.
Las primeras investigaciones sobre el hábitat en híbridos revelaban que sólo podían ocupar ambientes intermedios (Anderson, 1948; Muller, 1952), pero trabajos más recientes han evidenciado probabilidad de que se desarrollen en los mismos, similares o completamente distintos a los ocupados por los parentales (Cruzan y Arnold, 1993). El híbrido Quercus havardi Rydb. x Q. mohriana Buckley ex Rydb. presenta poca diferencia morfológica y causas parentales y se identificó conviviendo con ellas (Muller, 1952). Por otro lado, híbridos de Iris brevicaulis Raf. x fulva con tendencia hacia I. brevicaulis conviven con las mismas especies vegetales, y muestran preferencia por los hábitats de I. brevicaulis ; no obstante, los que tienen propensión hacia I. fulva no tienen afinidad por hábitats parentales, solamente se les observa en nuevos (Cruzan y Arnold, 1993).
De acuerdo con algunos estudios florísticos, en México, se identificó que Q. rugosa se asocia exclusivamente con 24 especies arbóreas, a diferencia de Q. glabrescens que lo hace exclusivamente con 20 (además de tener 20 taxa en común) (Cuadro 4). En función de la asociación de especies, las comunidades de Q. glabrescens y Q. rugosa pueden diferenciarse entre sí (Figura 1 y Cuadro 4).
Es necesario realizar estudios moleculares que corroboren los resultados aquí planteados.
CONCLUSIONES
Los resultados del índice de hibridación morfológico permitieron determinar la existencia de hibridación entre Q. glabrescens y Q. rugosa . Los híbridos con tendencia a Q. glabrescens mostraron una fuerte afinidad por hábitats semejantes a los de la especie parental (terrenos con alta pendiente, exposiciones húmedas, neblina frecuente, suelos con nitrógeno, zinc, manganeso y materia orgánica). Los híbridos intermedios exhibieron afinidades por hábitats con características combinadas entre los ocupados por las especies parentales. La ocurrencia de un mosaico de fenotipos puede sugerir introgresión híbrida en la zona.
AGRADECIMIENTOS
A la Comisión Nacional Forestal y Consejo Nacional de Ciencia y Tecnología por el apoyo financiero aportado a través del proyecto de investigación CONAFOR-2002-C01-6181. A la Universidad Autónoma Chapingo por el financiamiento proporcionado a través del proyecto 06100506.
REFERENCIAS
Albarrán-Lara, A. L., L. Mendoza-Cuenca, S. Valencia-Ávalos, A. González-Rodríguez and K. Oyama. 2010. Leaf fluctuating asymmetry increases with hybridization and introgression between Quercus magnoliifolia and Quercus resinosa (Fagaceae) through an altitudinal gradient in Mexico. International Journal of Plant Science 171: 310-322. [ Links ]
Álvarez-Moctezuma, J. G., S. Ochoa-Gaona, B. H. Jong, y M. L. Soto-Pinto, 1998. Hábitat y distribución de 5 especies de Quercus (Fagaceae) en la Meseta Central de Chiapas, México. Chiapas, México. Rev. Biol. Trop. 47(3):351-358. [ Links ]
Anderson, E. 1948. Hybridization of the habitat. Evolution. 2(1):1-9. [ Links ]
Arnold, M. L. 1992. Natural hybridization as an evolutionary process. Annual Review of Ecology and Systematics 23:237-261. [ Links ]
Arnold, M. L. 1997. Natural hybridization and evolution. Oxford University Press. Oxford, UK. 232 p. [ Links ]
Bacilieri, R., A. Ducousso, R. J Petit and A. Kremer. 1996. Mating system and asymmetric hybridization in a mixed stand of European oaks. Evolution 50: 900-908. [ Links ]
Bello G., M. y J. N. Labat. 1987. Los encinos ( Quercus ) del Estado de Michoacán, México. Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP). Secretaría de Agricultura y Recursos Hidráulicos. México, D. F. México 93 p. [ Links ]
Bruschi, P., G. Vendramin, F. G Bussotti and P. Grossoni. 2003. Morphological and molecular diversity among Italian populations of Quecus petraea (Fagaceae). Annals of Botany 91:707-716. [ Links ]
Catalán-Heverástico, C., L. López-Mata y T. Terrazas. 2003. Estructura, composición florísticas y diversidad de especies leñosas del bosque mesófilo de montaña de Guerrero, México. Anales del Instituto de Biología Serie Botánica 74(2): 209-230. [ Links ]
Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (CONABIO), 2000. Comisión Nacional de Biodiversidad. Quercus rugosa . Anales de Ciencias Naturales. http://www.conabio.gob.mx/conocimiento/info.../31-fagac10m.pdf (5 de septiembre de 2006). [ Links ]
Comisión Nacional Forestal (CONAFOR), 2000. Quercus rugosa. www.conafor.gob.mx/'portal/docs/secciones/bosquedes/Fichas%20Tecnicas/Quercus%20rugosa.pdf#search=%22%quercus%20rugosa%22 (5 de septiembre de 2006). [ Links ]
Comisión Nacional de Áreas Naturales Protegidas (CONANP), 2004. Programa de conservación y manejo Parque Nacional El Chico. http://www.ine.gob.mx/ueajei/publicaciones/libros/108/hgo.html (5 de septiembre de 2006). [ Links ]
Cornejo-Tenorio, G., A. Casas, B. Farfán, J. Villaseñor y G. Ibarra-Manríquez. 2003. Flora y vegetación de zonas núcleo de la Reserva de la Biosfera Mariposa Monarca, México. Bol. Soc. Bot. Méx. 73:43-62. [ Links ]
Cruzan, M. B. and M. L. Arnold. 1993. Ecological and genetic associations in an Iris hybrid zone. Evolution 47(5): 1432-1445. [ Links ]
Fernández-Nava., J. Rodríguez, S. Arreguín C. y A. Rodríguez J. 1998. Listado florístico de la cuenca del río Balsas, México. Polibotánica 9:1-151. [ Links ]
García-Arévalo, A., J. Mendoza-Contreras y J. Nocedal. 2004. Asociaciones vegetales de los bosques del municipio de Guanaceví, Durango. Madera y Bosques 10(1):21-34. [ Links ]
González-Rodríguez A., M. Arias, S. Valencia and K. Oyama. 2004. Morphological and RAPD analysis of hybridization between Quercus affinis and Quercus laurina (Fagaceae), two Mexican red oaks. American Journal of Botany 91:401-409. [ Links ]
González-Rodríguez, A. and K. Oyama. 2005. Leaf morphometric variation in Quercus affinis and Q. laurina (Fagaceae), two hybridizing Mexican red oaks. Botanical Journal of the Linnean Society 147: 427-435. [ Links ]
Grant, V. 1989. Especiacion vegetal. Columbia University Press. 2da. Edición. Edit. Limusa. México, D. F. México. 587 p. [ Links ]
Instituto Nacional de Estadística, Geografìa e Información (INEGI). 1984. Cartas topográficas 1:50000. E14-A56. México, D. F. México. s/p. [ Links ]
José-Noguera, A., G. Castro-Marín y B. González-Rivas 2006. Diversidad florística del bosque de galería en dos localidades del departamento de Carazo, Nicaragua. La Calera, Ciencias Forestales. http://www.bio-nica.info/biblioteca/NogueraBosqueDeGaleriaCarazo.pdf (2 septiembre de 2010). [ Links ]
Keim, P., K. N. Paige, T. G. Whitham and K. G. Lark. 1989. Genetic analysis of an interspecific hybrid swarm of Populus : Ocurrence of unidirectional introgression. Genetics Society of America 123: 557-565. [ Links ]
Krebs, Ch. J. 1989. Ecological methodology. Harper Collins. New York,NY. USA. 654 p. [ Links ]
Long, R. W. 1959. Natural and artificial hybrids of Helianthus maximiliani x H. grosseserratus. American Journal of Botany 46(10):687-692. [ Links ]
Lundell, C.L. 1955. Flora of Texas. Vo. I Texas Research Foundation. Renner, TX. USA. 1881 p. [ Links ]
Martínez, G., M., D. R. Cruz, J. Castrejón R., S. Valencia A., J. Jiménez R. y C. Ruíz-Jiménez. 2004. Flora vascular de la porción guerrerense de la Sierra de Taxco, Guerrero, México. Anales del Instituto de Biología, Universidad Nacional Autónoma de México, Serie Botánica 75(2):105-189. [ Links ]
Mejía D. N., J. Meave A. y C. Ruíz-Jiménez. 2004. Análisis estructural de un bosque mesófilo de montaña en el extremo oriental de la Sierra Madre del Sur (Oaxaca), México. Bol. Soc. Bot. Méx. 74:13-29. [ Links ]
Mettler, L. y T. Gregg. 1982. Genética de las poblaciones y evolución. Prentice Hall. 1a Edición. Englewood Clifts, NJ. USA. 245 p. [ Links ]
Muller, C. H. 1952. Ecological control of hybridization in Quercus : A Factor in the mechanism of evolution. Evolution 6(2):147-161. [ Links ]
Nixon, K.C. 1993. The genus Quercus in México. Chapter 16: 447-458. In : Ramamoorthy, T. P., R. Bye, A. Lot and J. Fa (Eds.). Biological Diversity of Mexico: Origins and distribution. Oxford University Press. NY. USA. 812 pp. [ Links ]
Secretaría de Medio Ambiente y Recursos Naturales (SEMARNAT) 2000. NOM-021-RECNAT-2000 Diario Oficial de la Federación. México, D.F. México. 73 p. [ Links ]
Olsen, R. S., V. C. Cole, F. S. Watanabe and L. A. Dean. 1954. Estimation of available phosphorus in soils by extraction with sodium bicarbonate. USDA. Washington, D.C. USA. Circular 939. 43 p. [ Links ]
Ortega, E. 1981. Química de suelos. Universidad Autónoma Chapingo. Departamento de Suelos. Texcoco. Edo. de Méx. México. 400-416 p. [ Links ]
Potts, B. M. and J. B. Reid. 1985. Analysis of a hybrid swarm between Eucalyptus risdonii Hook,f. and E. amygdalina Labill. Aust. J. Bot. 33:543-62. [ Links ]
Quiroga, P., R. Vidal R. y C. Premoli A. 2005. Evidencia morfológica e isoenzimática de hibridación natural entre Nothofagus antarctica y N. pumilio en el noroeste patagónico. Bosque 26(2):25-32. [ Links ]
Ramírez-Marcial, N., S. Ochoa-Gaona, M. González-Espinosa y P. Quintana-Ascencio. 1998. Análisis florístico y sucesional en la estación biológica cerro Huitepec, Chiapas, México. Acta Botánica Mexicana 44:59-85. [ Links ]
Rzedowski, J. 1978. Vegetación de México. Ed. Limusa. México, D.F. México. 432 p. [ Links ]
Rieseberg, L. H. 1997. Hybrid origins of plant species. Annual Review of Ecology and Systematics 28:359-389. [ Links ]
Rieseberg, L. H. and S. E. Carney. 1998. Plant hybridization. New Phytologist 140(4):599-624. [ Links ]
Statistical Analysis System (SAS) Institute 1985. SAS introductory guide. Raleigh, NC. USA. 99 p. [ Links ]
Sistema de Información Climatológica (SICLIM). Versión 1.0 y base de datos. 1990. Instituto Mexicano de Tecnología del Agua. Jiutepec, Mor. México. s / p. [ Links ]
Standley, P. C. and J. A. Steyermark 1954. Flora of Guatemala. Fieldana: Botany. Vol. 24. Part III. Chicago Natural History Museum. Chicago, IL. USA. p. 369-396. [ Links ]
Tovar-Sánchez, E. and K. Oyama. 2004. Natural hybridization and hybrid zones between Q. crassifolia and Q. crassipes (Fagaceae) in Mexico: morpho-logical and molecular evidence. American Journal of Botany 91:1352-1363. [ Links ]
Tovar-Sánchez, E. and K. Oyama. 2006. Community structure of canopy arthropods associated to Q. crassifolia X Q. crassipes complex. OIKOS 112:370-381. [ Links ]
Tucker, J. M. 1968. Identify of the oak tree al Live Oak Tanks, Joshua Tree National Monument, California. Madroño 19:256-266. [ Links ]
United States Departament Agriculture (USDA). 2003. USDA Plant Hardiness Zone Map. The United States National Arboretum. Agricultural Research Service, U.S. Department of Agriculture, Washington, D. C. www. National Arboretum - USDA Plant Hardiness Zone Map.htm. (20 de marzo de 2010). [ Links ]
Valencia A., S. 1989. Contribución al conocimiento del género Quercus en el estado de Guerrero, México. Universidad Nacional Autónoma de México. Tesis de Licenciatura. México, D.F. México.183 p. [ Links ]
Valencia, A. S. 2004. Diversidad del género Quercus (Fagaceae) en México. Bol. Soc. Bot. Méx. 75: 33-53. [ Links ]
Van Valen, L. 1976. Ecological species, multispecies and oaks. Taxon 25:233-9. [ Links ]
Vargas M., F. 1984. Parques nacionales de México y reservas equivalentes. Pasado, presente y futuro. Instituto de Investigaciones Económicas. Universidad Nacional Autónoma de México. México, D. F. México. 266 p. [ Links ]
Walkley, A. and A. Black. 1934. An examination of the Degthareff method for determining soil organic matter and a proposed modification of the chromic acid titration method. Soil. Sci. 27:29-38. [ Links ]
Zavala Ch., F. 1989. Identificación de encinos de México. División de Ciencias Forestales. Universidad Autónoma Chapingo. Chapingo, Texcoco, Edo. de Méx. México. 150 p. [ Links ]
Zavala Ch., F. 1990. Los encinos de México: un recurso desaprovechado. Ciencia y Desarrollo 16(95):43-51. [ Links ]
Zavala Ch., F. 1995. Encinos hidalguenses. División de Ciencias Forestales. Universidad Autónoma Chapingo. Chapingo. Texcoco, Edo. de Méx. México. 133 p. [ Links ]

