











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
Los agostaderos distribuidos alrededor del mundo cubren más del 50 % de la superficie terrestre, y aportan biomasa que brinda un servicio ecosistémico fundamental del cual dependen la fauna silvestre, la población y la ganadería1,2, la principal actividad económica en este ecosistema. En el último siglo, los agostaderos han sufrido degradación debido a episodios de sequía y sobrepastoreo por una carga animal excesiva3,4, por fallas en un aprovechamiento eficiente y sostenible del agostadero para cubrir los requerimientos de biomasa forrajera del hato ganadero5. Por lo anterior es importante contar con estimaciones confiables de la cantidad de forraje disponible para el ganado, que permitan hacer un uso adecuado del forraje, evitar el sobrepastoreo y satisfacer las necesidades de los animales6,7.
El método tradicional para estimar la producción de biomasa aérea es el de corte y pesado del pasto, aunque este método destructivo es preciso, suele ser costoso y tardado8,9. Además, la distribución aleatoria de la vegetación obliga a incrementar el número de muestras recomendada por sitio (15-20 muestras10), colectadas idealmente en cada estación de crecimiento del forraje8.
Los métodos indirectos (no destructivos, ya que no requieren cortar el pasto presente), surgen como alternativa al tradicional (destructivo, que implica cortar y pesar el pasto presente), para determinar la biomasa aérea de forraje en el agostadero, ya que tienen la ventaja de lograr estimaciones de la biomasa de extensas superficies de una forma más rápida11,12. Los métodos empíricos subjetivos de estimación visual tienen la desventaja de una alta variación en los resultados entre las personas que lo realizan en diferentes periodos de tiempo13. Platos o bastones graduados han sido utilizados en décadas recientes, para estimar la biomasa forrajera en praderas con vegetación homogénea14,15. Nuevas metodologías para la estimación de la biomasa han sido desarrolladas mediante la utilización de imágenes satelitales16, imágenes de radar17 y a través de vehículos no tripulados18, sin embargo, estas son realizadas principalmente en áreas de cultivo.
Los agostaderos presentan diversos tipos de vegetación, con distribución heterogénea. Por ejemplo, en el matorral espinoso tamaulipeco (MET) predominan las especies arbustivas y semiarbustivas, que constituyen en ocasiones más del 80 % de la composición botánica, mientras que las especies gramíneas y otras hierbas difícilmente superan el 10 y 6 % respectivamente19,20. Bajo estas condiciones los modelos alométricos desarrollados a partir de relacionar los datos de producción de biomasa obtenidos del método tradicional, con las mediciones hechas en características morfológicas del individuo, son una buena opción para estimar objetivamente la biomasa de los pastos. Una vez generado el modelo, la estimación de la biomasa se puede realizar solamente midiendo las variables vegetativas necesarias, sin necesidad de cortar las plantas5,9.
Estudios previos al presente trabajo han sido realizados en praderas bajo condiciones de riego y monocultivos15,21,22. Para condiciones de agostaderos, se han publicado resultados obtenidos en condiciones áridas de Arizona23 y de multiespecies en Argentina9. Existen reportes de ecuaciones generadas específicamente para la estimación de la biomasa de ciertas especies de pastos, tales como las reportadas a partir de 40 plantas de praderas de pasto de C. ciliaris en el sur de Arizona, relacionando la biomasa, con el diámetro basal y la altura de la planta24, así como las ecuaciones generadas para A. purpurea en un estudio previo25, utilizando el diámetro de la planta a diferentes alturas como variables.
En Europa se desarrolló una metodología de estimación indirecta para especies de pastos21, que utilizó una medida denominada volumen mínimo, obtenido al juntar todos los tallos de la planta aplicando una fuerza subjetiva, no estandarizada, hasta formar un volumen mínimo.
En el presente estudio se planteó el objetivo de generar nuevas ecuaciones para estimar a partir de la medición de sus variables morfológicas, la biomasa aérea de cinco especies de pastos presentes en agostaderos de Nuevo León, México, que pudiera servir como alternativa para reemplazar la necesidad de cortar pasto como en el método tradicional.
Material y métodos
El estudio se realizó en un área de 132 ha de agostaderos en el municipio de Marín, Nuevo León (25°52'28"N; 100°03'24"O), en la que la precipitación varía entre 400 y 600 mm anuales, y la temperatura media varía de 20 a 22 °C26. El tipo de vegetación principal es de matorral espinoso tamaulipeco (MET), además de contar con áreas de pastizal inducido y agrícolas.
En Enero de 2021 se establecieron de forma aleatoria dentro del área de estudio con vegetación propia del matorral espinoso tamaulipeco, 31 parcelas de muestreo de 100 metros cuadrados (10 m x 10 m), y dentro de cada parcela se delimitaron cinco subparcelas de 1 m2, en las cuales se realizó la colecta y se registró el peso (base materia seca) de todos los individuos de pastos presentes en cada una de las 155 subparcelas evaluadas, en un diseño similar al utilizado previamente en áreas de agostadero27. Se colocó un cerco perimetral alrededor de cada parcela, para evitar disturbios. Todas las parcelas se cortaron una altura de 3 a 5 cm, al inicio del estudio y en cada muestreo. El primer muestreo se llevó a cabo del 15 de junio al 15 de julio del 2021, y el segundo muestreo se realizó en otoño, del 18 de octubre al 8 de noviembre del 2021. En ambos casos el muestreo se llevó a cabo 30 a 40 días después de presentarse una precipitación superior a 150 mm (en el primer muestreo) y de 231 mm (en el caso del segundo muestreo), ya que de acuerdo a estudios previos28, la floración del pasto Cenchrus ciliaris ocurre entre 25 y 35 días después del rebrote a consecuencia de precipitación de 150 mm, lo que se considera como el umbral para la productividad de la especie Cenchrus ciliaris29 durante el verano y el otoño.
En cada subparcela se muestrearon e identificaron individualmente a nivel de género y especie, todas las plantas de las especies de gramíneas presentes. Se utilizó un flexómetro marca Truper® modelo pro-Lock FX-5M, para medir las variables descritas a continuación y mostradas en la Figura 1A:
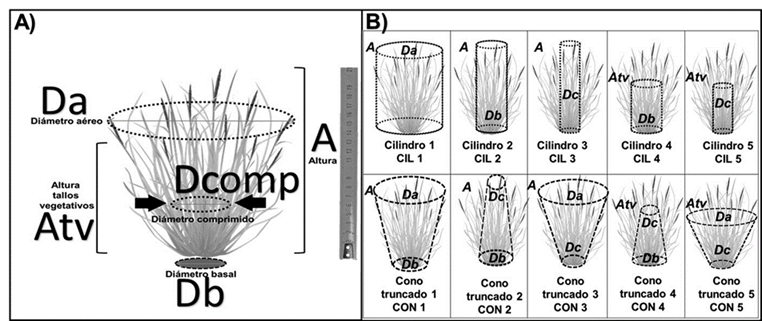
A= altura; Atv= altura de tallos vegetativos; Db= diámetro basal; Dc= diámetro comprimido; Da= diámetro aéreo.
Figura 1 Variables medidas en las plantas de pastos (A) y formas de volúmenes estimados (B)
Altura máxima (A): distancia entre el suelo y la parte más alta de los tallos y hojas.
Altura de los tallos vegetativos (Atv): distancia entre el suelo y la mayor parte de las hojas vegetativas, generalmente aquellas que no tienen espiga.
Diámetro basal (Db): de la circunferencia de la base de la planta.
Diámetro aéreo (Da): a la altura de los tallos vegetativos.
Utilizando un calibrador digital marca Traceable® modelo 6˝, se midió el diámetro comprimido (Dcomp, Figura 1) a la mitad de la altura del pasto, utilizando un prototipo experimental que, por medio de una banda retráctil, aplica una presión uniforme graduada de 2 kg en los tallos vegetativos.
Utilizando las variables morfológicas de la Figura 1A, se calculó la cobertura aérea en forma circular (COBAC), cobertura aérea en forma elipsoidal (COBAE), cobertura basal circular (COBBC) y cobertura basal elipsoidal (COBBE). Los volúmenes de cilindro (en sus modalidades CIL 1 a CIL5), y cono (en modalidades CON 1 a CON5), señalados en la Figura 1B, se basaron en propuestas previas30.
Cada una de las 745 plantas muestreadas, fue identificada, medida, y cortada con tijeras de mano a nivel del suelo, para registrar su peso verde (g) en campo, y se almacenó en bolsa de papel Kraft. Posteriormente las muestras se llevaron a laboratorio y secadas en una estufa de aire forzado a 60 °C hasta llegar a peso constante, con el fin de obtener su peso seco (g), utilizando una báscula con capacidad de 500 g con división mínima de 0.1 g (marca Torrey, modelo Lab-500).
Se identificaron, midieron y cortaron los 745 individuos encontrados en las subparcelas de muestreo. Se colectaron las plantas de las cinco especies de pastos presentes en el área de estudio: Cenchrus ciliaris (n= 424 individuos), Pappophorum bicolor (n= 125 individuos), Aristida purpurea (n= 107 individuos), Tridens texanus (n= 59 individuos) y Paspalum pubiflorum (n= 30 individuos), para la generación de ecuaciones.
Un total de 19 variables independientes (diámetro aéreo, diámetro basal, diámetro comprimido, altura, altura de los tallos vegetativos, cobertura basal circular, cobertura basal elíptica, cobertura aérea circular, cobertura aérea elíptica, volumen cilindro 1, volumen cilindro 2, volumen cilindro 3, volumen cilindro 4, volumen cilindro 5, volumen cono 1, volumen cono 2, volumen cono 3, volumen cono 4, volumen cono 5) y una variable dependiente (biomasa medida [g MS] por individuo de las especies de pastos) fueron sometidas a análisis de regresión lineal, de pasos sucesivos9 y regresión no lineal (logarítmico, inverso, cuadrático, cubico, potencia, exponencial) en el software IBM SPSS, para generar ecuaciones de predicción de biomasa tanto de forma general y de forma específica (por cada una de las cinco especies de pastos encontrados en los muestreos).
Para evaluar la calidad de la estimación de biomasa lograda con las nuevas ecuaciones generales y específicas, se realizaron comparaciones de cada uno de los valores registrados en el muestreo destructivo, con los valores pronosticados de cada una de las ecuaciones. Se procedió a calcular el coeficiente de determinación de la regresión (R2)27, el error estándar (EE)30, la correlación de Pearson (r)18, la raíz del error cuadrático medio normalizado (NRMSE)21 y el criterio de información de Akaike (AIC)27.
Se seleccionaron las seis ecuaciones generales desarrolladas con los mejores ajustes estadísticos de AIC, NRMSE, R2, EE, r, para la estimación del total de las especies colectadas, y una ecuación específica para cada una de las cinco especies de pasto registradas.
Resultados
Los pastos Cenchrus ciliaris y Paspalum pubiflorum presentaron valores superiores de cobertura aérea y basal, diámetro aéreo y basal, peso verde y seco (P<0.05) al resto de las especies (Cuadro 1). Cenchrus ciliaris registró un mayor valor promedio de peso seco por individuo al de Aristida purpurea y Tridens texanus (P<0.05), mientras que Paspalum pubiflorum y Pappophorum bicolor obtuvieron valores intermedios.
Cuadro 1 Valores promedio de cobertura aérea, cobertura basal, diámetro aéreo, diámetro basal, diámetro comprimido, altura máxima, altura de los tallos vegetativos, peso verde y peso seco por individuo de cada especie en las subparcelas experimentales de 1m2
| Especie | Cobertura | Diámetro | Altura | Peso | |||||
|---|---|---|---|---|---|---|---|---|---|
| Aérea (cm2) |
Basal (cm2) |
Aéreo (cm) |
Basal (cm) |
Comprimido (mm) |
Máxima (cm) |
Tallos vegetativos (cm) |
Verde (g) |
Seco (g) |
|
| Cenchrus ciliaris | 729ab | 190a | 28a | 14a | 19a | 52a | 27a | 53a | 22a |
| Pappophorum bicolor | 355bc | 52bc | 20bc | 8bc | 13abc | 52a | 23ab | 15bcd | 10ab |
| Aristida purpurea | 277bc | 29c | 17cd | 6c | 9bcd | 45ab | 20bc | 7cd | 5b |
| Tridens texanus | 208c | 37bc | 16cd | 6c | 6d | 34cd | 16c | 5d | 3b |
| Paspalum pubiflorum | 799a | 104abc | 30a | 11ab | 13ab | 40bc | 23ab | 48ab | 13ab |
abcd Diferentes letras dentro de la misma columna indican diferencia significativa (P<0.05).
Las estimaciones de biomasa aérea calculadas con las seis nuevas ecuaciones generales generadas en el presente estudio, tuvieron coeficientes de determinación (R2) que variaron entre 0.77 y 0.90, mientras que el coeficiente de correlación de Pearson (r) de 0.88 a 0.94. La raíz del error cuadrático medio normalizado (NRMSE) varió de 0.68 a 0.48 y el criterio de información de Akaike (AIC) tomo valores de 3553 a 3079 (Cuadro 2).
Cuadro 2 Ecuaciones alométricas generales y específicas generadas a partir de variables morfológicas para estimar indirectamente la biomasa aérea de los pastos (g MS-1)
| Id | Ecuación | R2 | EE | r | NRM SE |
AIC |
|---|---|---|---|---|---|---|
| Ecuaciones generales | ||||||
| I | Y= 0.9648 + 0.0026CON5 | 0.77 | 10.1 | 0.88 | 0.64 | 3469 |
| II | Y= 2.5343 + 0.0027CON5 + 0.0139CIL3 - 0.0295COBAC +0.8253Dcomp - 0.2595Atv + 0.0002CIL1 | 0.87 | 7.8 | 0.93 | 0.49 | 3108 |
| III | Y= 1.2159 + 0.0032CON5 + 0.0447CIL3 - 0.0421COBAC + 0.8939Dcomp - 0.3478Atv + 0.0003CIL1 - 0.0253CON2 + 0.5790Db + 0.0084CIL2 | 0.88 | 7.5 | 0.94 | 0.48 | 3079 |
| IV | Y= 0.1213 * Dcomp 1.6818 | 0.86 | 0.4 | 0.88 | 0.67 | 3530 |
| V | Y= 0.4473 * CIL3 0.7288 | 0.89 | 0.4 | 0.88 | 0.68 | 3553 |
| VI | Y= 0.8084 * CIL5 0.7078 | 0.90 | 0.4 | 0.88 | 0.67 | 3530 |
| Ecuaciones específicas | ||||||
| C.c. | Y= 0.2862 + 0.0032CON5 - 0.0753COBAC + 1.4623Dcomp - 0.0767CON4 + 0.0279CIL4+ 0.0902COBBC - 0.3257Atv + 0.0022CIL1- 0.0032CON1 + 0.0931CIL5 | 0.88 | 9.0 | 0.94 | 0.40 | 1603 |
| P.b. | Y= 2.1060 + 0.0490CIL3 + 0.0050CON4 + 0.0002CON3 - 0.0050COBBE | 0.86 | 3.0 | 0.92 | 0.29 | 287 |
| A.p. | Y= -0.6641 + 0.1138CIL5 + 0.1257Da + 0.0046CON2 - 0.0816COBBC + 0.1951Dcomp | 0.92 | 1.5 | 0.96 | 0.27 | 87 |
| T.t. | Y= 0.5474 + (0.1672 * CIL3) + (-0.0012 * CIL3) + (1.18 × 10 -5 * CIL3) | 0.91 | 0.9 | 0.96 | 0.30 | -18 |
| P.p. | Y= -0.0371 +(0.2744 * CIL3) + (-0.0020 * CIL3) + (6 × 10-6 * CIL3) | 0.93 | 2.3 | 0.97 | 0.24 | 57 |
Ecuaciones de modelo lineal= I, II, III, C.c., P.b., y A.p.; Ecuaciones de modelo potencial= IV, V y VI; Ecuaciones de modelo cúbico= T.t. y P.p.; Y= Biomasa aérea (g MS-1); R2= Coeficiente de determinación; EE=Error estándar; r= Coeficiente de correlación de Pearson; NRMSE= Raíz cuadrada del error cuadrático medio normalizado; AIC= Criterio de Información de Akaike; C.c.=Cenchrus ciliaris; P.b.= Pappophorum bicolor; A.p.= Aristida purpurea; T.t.= Tridens texanus; P.p.= Paspalum pubiflorum. Ver Figura 1 para Db, Da, Dcomp, A, Atv, COBBE, COBBC, COBAC, CIL1, CIL2, CIL3, CIL5, CON1, CON2, CON3, CON4, CON5. Todos los coeficientes de regresión fueron significativos (P<0.05).
La variable cilindro 3, calculada a partir de la altura de la planta y el diámetro comprimido, fue la base para generar las ecuaciones específicas para Tridens texanus y Paspalum pubiflorum en modelo cúbico, cuyas estimaciones tuvieron un ajuste de R2 de 0.91, r= 0.96, NRMSE= 0.30, AIC= -18 para Tridens texanus (Figura 2F) y de R2= 0.93, r= 0.97, NRMSE= 0.24, AIC= 57 para P. pubiflorum.
La ecuación I es un modelo lineal, que incorpora la medida del cono 5 (Figura 1B), el cual es calculado a partir de tres variables directas (diámetro aéreo, diámetro comprimido y altura de los tallos vegetativos), y cuyas estimaciones tienen un R2 de 0.77, r= 0.88, NRMSE= 0.64, AIC= 3469. Las estimaciones calculadas con la ecuación II (lineal) a partir de los datos generados por las variables diámetro aéreo, diámetro comprimido, altura y altura de los tallos vegetativos, tienen un R2 de 0.87, r= 0.93, NRMSE= 0.49, AIC= 3108. La ecuación III (lineal) incorpora los datos de las cinco variables medidas para calcular estimaciones que tienen un R2 de 0.88, r= 0.94, NRMSE= 0.48, AIC= 3079 (Figura 2A).

A) Ecuación general III (Cuadro 2). B) Ecuación general IV (diámetro comprimido). C) Ecuación específica para Cenchrus ciliaris. D) Ecuación específica para Pappophorum bicolor (Cuadro 2). E) Ecuación específica de Aristida purpurea. F) Ecuación específica de Tridens texanus (Cuadro 2). Todas las gráficas fueron ajustadas a cero.
Figura 2 Regresión de los valores pronosticados y los valores observados
Las ecuaciones no lineales IV (R2=0.86, r= 0.88, NRMSE= 0.67, AIC= 3530; Figura 2B), V (R2=0.89, r= 0.88, NRMSE= 0.68, AIC= 3553) y VI (0.90, r= 0.88, NRMSE= 0.67, AIC= 3530), son del modelo de potencia, y utilizan menor cantidad de variables. Las estimaciones calculadas utilizando la ecuación IV, que utiliza el diámetro comprimido como única variable tienen un R2=0.86, (Figura 2B). La ecuación V utiliza el cilindro 3, calculado a partir del diámetro comprimido y de la altura de la planta, para estimar biomasa aérea de pastos con R2=0.89. La ecuación VI utiliza el cilindro 5 (Figura 1B), calculado a partir del diámetro comprimido y la altura de los tallos vegetativos para estimar biomasa aérea con R2=0.90 (Cuadro 2).
En el caso de las ecuaciones generadas específicamente para cada una de las especies de pastos, las comparaciones entre los valores estimados y los resultados registrados directamente de biomasa aérea arrojaron valores de R2 de 0.86 hasta 0.93. Los valores de r(Pearson) variaron de 0.92 a 0.97. Para NRMSE se registraron valores de 0.40 a 0.24 y AIC tomó valores de 1603 a -18 (Cuadro 2). En el caso específico de la ecuación generada para Cenchrus ciliaris se logró un buen ajuste de los valores estimados de biomasa aérea, con R2 de 0.88, r= 0.94, NRMSE= 0.40, AIC= 1603 utilizando las cinco variables medidas (Figura 2C).
Los resultados estimados utilizando las cinco variables con la ecuación específica para la especie Pappophorum bicolor (Cuadro 2) tuvieron un ajuste de (R2 = 0.86, r = 0.92, NRMSE = 0.29, AIC = 287) (Figura 2D).
La información recopilada de 107 individuos de Aristida purpurea, permitió generar una ecuación específica para esta especie de pasto (Cuadro 2), cuyas estimaciones a partir de las cinco variables medidas tuvieron un ajuste con un coeficiente de determinación de 0.92, r= 0.96, NRMSE = 0.27, AIC = 87. (Figura 2E).
Discusión
Las ecuaciones lineares y polinómicas generadas en el presente estudio a partir del muestreo de medidas vegetativas y de registros de peso de biomasa de gramíneas presentes en agostadero, permitieron estimar la biomasa aérea de gramíneas presentes en el agostadero, con alto grado de precisión.
Las nuevas ecuaciones alométricas generales establecidas en el presente estudio, tuvieron valores de R2 (0.77 a 0.93) superiores a los reportados previamente (R2 de 0.25 a 0.85) para estimaciones de ecuaciones generales para dos gramíneas y dos pseudogramíneas en los Andes Peruanos27. Ecuaciones alométricas generadas en Chubut, Argentina23, evaluando 50 individuos de tres especies de gramíneas, tuvieron valores de R2 (de entre 0.72 y 0.86), similares a las obtenidas en el presente estudio.
El número de parcelas establecidas en el presente estudio fue similar al utilizado previamente27, sin embargo, en el presente estudio se tuvieron parcelas de 100 m2, mientras que en el estudio previo27, tuvieron parcelas de 4 m2. Además, en cada una de las 31 parcelas del presente estudio, se registraron los valores de composición botánica, medidas vegetativas y biomasa de las gramíneas presentes en cinco subparcelas. Con ello se tuvo mayor confiabilidad de las mediciones registradas y de los promedios calculados, para la generación de las ecuaciones.
La especie C. ciliaris tuvo presencia importante en la vegetación de gramíneas observada en el agostadero evaluado, ya que representó el 57 % del número total (n=745) de individuos colectados y su biomasa, calculada tomando en cuenta el número de individuos y el peso promedio de los individuos de la especie mostrados en el Cuadro 1, representó el 80 % de la biomasa registrada en el agostadero evaluado. En las áreas evaluadas en el presente estudio, el pasto C. ciliaris presente se estableció por dispersión natural, dejando en evidencia su alto potencial para establecerse en los agostaderos en México31. La especie nativa con mayor presencia en el presente estudio fue Pappophorum bicolor, que tuvo registro de 125 individuos, es decir el 17 % del total, cuya biomasa representó solo 11 % de la biomasa total (Cuadro 1).
El diámetro comprimido fue la variable incluida en el 100 % en las ecuaciones alométricas generales y en el 97 % de las ecuaciones específicas, tanto para el modelo lineal como no lineal, generadas en el presente estudio para cada especie de pasto. La altura de los tallos vegetativos fue incluida en el 87 % de las ecuaciones generales. La variable altura de los tallos vegetativos y la altura de la planta fueron incluidas en el 71 % de las ecuaciones específicas. Estas variables han sido directamente relacionadas con la densidad del forraje32. Algunos autores30 reportaron que en praderas con condiciones óptimas para su desarrollo, la variable cobertura vegetal es la que mejor puede estimar indirectamente la biomasa. Mahood et al22, determinaron que la cobertura vegetal es un buen predictor para la estimación de biomasa, con R2 de hasta 0.89 en comunidades vegetales de Bromus tectorum.
Una metodología de estimación indirecta implementada para especies de pastos del oeste de Europa21, utilizó como variable una medida denominada volumen mínimo, consistente en juntar manualmente todos los tallos de la planta hasta formar un volumen mínimo; sin embargo, la fuerza aplicada fue subjetiva, variando de acuerdo con la persona que realizara la medición e incluso podría variar cuando la misma persona ejerce diferente presión por cansancio al hacer repetidamente el muestreo. El dispositivo de compactación utilizado en el presente trabajo aplicando la misma fuerza de compresión generó mayor certidumbre de los resultados obtenidos.
Los valores de NRMSE calculados en cada una de las nuevas ecuaciones generadas en el presente estudio determinan la dispersión de los datos estimados respecto a los datos observados, siendo 0 el ajuste ideal33. En contraste con ello, el criterio de Akaike compara y selecciona de entre un grupo de modelos de predicción que utilizan los mismos datos experimentales, el más adecuado para pronosticar los valores esperados en comparación con los valores observados, que en este caso debería ser el modelo con el menor valor AIC34,35. Los valores de NRMSE y AIC son de gran utilidad en la selección de los mejores modelos.
La ecuación específica generada en el presente estudio para Cenchrus ciliaris tuvo valor de R2= 0.87, y con ello, R2 similar al reportado en un estudio (R2 de 0.8224) para generar ecuaciones alométricas relacionando la biomasa con la medición del diámetro basal y la altura de 40 plantas de praderas de pasto de C. ciliaris en el sur de Arizona. La ecuación generada en el presente estudio para A. purpurea resultó con un R2= 0.89, similar a los reportados (R2 de 0.82 a 0.90) en un estudio previo25, para la especie A. purpurea, utilizando el diámetro de la planta a diferentes alturas. Algunos autores30 generaron ecuaciones para estimar biomasa aérea a partir del análisis de 93 plantas de Agropyron desertorum y reportaron coeficientes de variación (R2 de 0.76 a 0.88) ligeramente inferiores a los del presente estudio. Las nuevas ecuaciones generadas en este estudio son candidatas potenciales para sustituir la fase de corte secado y pesado que se realiza en el método tradicional36.
El desarrollo de ecuaciones alométricas con aplicación en agostadero es de suma importancia para lograr un manejo sustentable del ecosistema27. Estos muestreos de vegetación no destructivos tienen ventajas con relación al tiempo y presupuesto respecto al muestreo tradicional, y permiten además el monitoreo del crecimiento a nivel de individuo21,37.
Conclusiones e implicaciones
Se evaluó la importancia de 19 variables relacionadas con las características morfológicas de pastos presentes en el agostadero para estimar la producción de biomasa aérea por planta. Con esas informaciones se generaron 6 nuevas ecuaciones generales para cinco especies de pasto presentes en el área de estudio, y 5 nuevas ecuaciones específicas (una para cada especie de pasto Cenchrus ciliaris, Pappophorum bicolor, Aristida purpurea, Tridens texanus y Paspalum pubiflorum), que estiman de manera confiable y práctica la producción de biomasa sin destruir (cortar) la planta. Las nuevas ecuaciones que se generaron en este estudio son una opción viable para sustituir la fase de corte de las muestras en el método tradicional de estimación de la producción de biomasa aérea de pastos en el agostadero. El desarrollo de ecuaciones alométricas generales (multiespecies) y específicas (para una especie de pasto en particular) son una alternativa confiable para estimar indirectamente la biomasa aérea de los pastos de los agostaderos del noreste de México, de forma más rápida y menos costosa que el método tradicional.

