










Serviços Personalizados
Journal
Artigo
 texto em
texto em  Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista mexicana de ciencias pecuarias
versão On-line ISSN 2448-6698versão impressa ISSN 2007-1124
Rev. mex. de cienc. pecuarias vol.14 no.3 Mérida Jul./Set. 2023 Epub 08-Set-2023
https://doi.org/10.22319/rmcp.v14i3.6251
Articles
Genetic parameters, correlations and trends of reproductive traits in Holstein cattle from Mexico
a Universidad Nacional Autónoma de México. Posgrado en Ciencias Biológicas, Circuito de Posgrados, Edificio D, 1er Piso, Ciudad Universitaria, Alcaldía Coyoacán. 04510, Ciudad de México, México.
b Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias. Centro Nacional de Investigación Disciplinaria en Fisiología y Mejoramiento Animal. Querétaro, México.
c Universidad Nacional Autónoma de México. Facultad de Medicina Veterinaria y Zootecnia. Ciudad de México, México.
d Universidad Nacional Autónoma de México. Instituto de Ecología, Departamento de Ecología Evolutiva. Ciudad de México, México.
The objective was to calculate the parameters, correlations and describe the genetic trends of reproductive traits in Holstein cattle from Mexico in three different periods of time, in females born between 2006 and 2019, using records of the Mexican Holstein association. The reproductive parameters calculated were: calving to first insemination interval (CFI), number of services per conception (NSC), days open (DO), interval between calving (IBC), and age at first calving (AFC). The components of variance were estimated using restricted maximum likelihood in an animal model, to calculate genetic parameters (narrow-sense heritability (h2), repeatability (r) and genetic correlations) and phenotypic correlations. Additive genetic variances and genetic values were also estimated in three periods of time (P1: 2006-2009, P2: 2010-2013 and P3: 2014-2017). The calculated heritabilities were from 4 to 9 % and the repeatabilities from 8 to 9 %, values close to previous reports for fertility traits. Phenotypic correlations were positive for almost all reproductive parameters and genetic correlations were positive over a wide range (0.13-0.99). Analyses by period showed changes, possibly derived from the influence of the import and use of germplasm from foreign bulls (mainly from the United States of America and Canada) which implement genomic selection and include fertility traits. The present research has made it possible to update the information on reproductive and genetic parameters in fertility traits that can be incorporated into national genetic evaluations.
Keywords Reproductive traits; Holstein cattle; Genetic parameters; Heritability; Repeatability
El objetivo fue calcular los parámetros, correlaciones y describir las tendencias genéticas de caracteres reproductivos en ganado Holstein de México en tres periodos diferentes de tiempo, en hembras nacidas entre 2006 y 2019, usando registros de la asociación Holstein de México. Los parámetros reproductivos calculados fueron: intervalo parto a primer servicio (IPPS), el número de servicios por concepción (NSC), los días abiertos (DAB), el intervalo entre partos (IEP) y la edad al primer parto (EPP). Los componentes de la varianza fueron estimados utilizando máxima verosimilitud restringida en un modelo animal, para calcular parámetros genéticos (heredabilidad en sentido estricto (h2), repetibilidad (r) y correlaciones genéticas) y correlaciones fenotípicas. También se estimaron varianzas genéticas aditivas y valores genéticos en tres periodos de tiempo (P1: 2006-2009, P2: 2010-2013 y P3: 2014-2017). Las heredabilidades calculadas fueron de 4 a 9 % y las repetibilidades del 8 al 9 %, valores cercanos a reportes previos para características de fertilidad. Las correlaciones fenotípicas fueron positivas para casi todos los parámetros reproductivos y las genéticas fueron positivas en un rango amplio (0.13-0.99). Los análisis por periodos mostraron cambios, posiblemente derivados de la influencia de la importación y uso de germoplasma de toros extranjeros (principalmente de los Estados Unido de América y Canadá) que implementan selección genómica e incluyen características fertilidad. La presente investigación ha permitido actualizar la información de parámetros reproductivos y genéticos en caracteres de fertilidad que podrán ser incorporados a las evaluaciones genéticas nacionales.
Palabras clave Caracteres reproductivos; Ganado Holstein; Parámetros genéticos; Heredabilidad; Repetibilidad
Introduction
In domestic animals such as cattle, the study of reproduction is based on records of the reproductive events of herds, including dates of birth, insemination and calving, with which it is possible to calculate reproductive parameters (RPs)1,2. Examples of RPs are the days from calving to first service/insemination, the percentage of conception at first service, the interval between calvings, the days from calving to conception, the number of services per conception and the age at first calving, which may be correlated with each other3,4. The reproductive parameters that are used for the study of cow fertility differ between countries3,5.
In dairy cattle of the Holstein breed, there are several reports published worldwide on a reduction in fertility from the 80s to the early 2000s6,7, apparently due to the genetic improvement of milk production7,8, which is related to a negative genetic correlation reported between fertility and milk production9,10.
For dairy farmers, a reduction in fertility causes significant losses and economic impacts10,11. By requiring a greater number of inseminations for a cow to become pregnant, insemination costs increase and the interval between calvings lengthens7,9, decreasing the average milk production of the herd, as well as the number of calves per cow10,11. Likewise, fewer cows will become pregnant within the required period, which increases the “involuntary disposal” or slaughter of the cow due to low fertility10,11, increasing costs due to an additional replacement animal is bought or raised in the farm11,12.
Therefore, the fertility and the beginning of the productive life of the cow are priority factors in the lifetime productivity of dairy cattle2,12. Their study is important for the future profitability of production systems and reduction of slaughter generated by deficient fertilities9,11.
To increase milk production in cattle, genetic selection has been used, with methodologies developed and implemented from phenotypic and genealogical records13,14. For a little over 20 yr, the use of genome-wide genetic markers, such as single-nucleotide polymorphisms (SNPs), has been proposed to aid selection, through genomic selection (GS)15, first used in Holstein cattle from the United States in late 200916.
For several generations in countries such as the United States of America and Canada, fertility traits in Holstein cattle have been evaluated and improved16,17, and the use of GS resulted in an increase in the rate of improvement, despite being generally low heritable traits16,17 and the possible impact that this selection has had on the genetic parameters and values of populations that import germplasm from these countries, such as Mexico, is not known. On the other hand, in Mexico there are few works with the aim of studying genetic aspects of fertility traits18,19,20, despite their high economic and functional impact on Mexican herds.
Therefore, the objective of the present study was to calculate the parameters, correlations and describe the genetic trends of reproductive traits in Holstein cattle from Mexico in three different periods of time.
Material and methods
Data and reproductive parameters analyzed
This study included 415,859 reproductive records of Holstein cows (Bos taurus taurus) that had information on milk production, insemination events (artificial (AI) or natural mounting (NM)) and calving, occurred between January 2006 and December 2019; information provided by the Holstein association of Mexico.
The RPs calculated and included in the present study were: calving to first insemination interval (CFI), measured as the days elapsed between calving and the first service/insemination; number of services per conception (NSC), it corresponds to the number of services/inseminations until gestation; days open (DO), are the days elapsed between the calving and the new gestation of the cow, the interval between calvings (IBC), measured as the number of months between two consecutive calvings, and finally, the age at first calving (AFC), which corresponds to the recorded age on which each cow has its first calving. The first four RPs together indicate the ability of cows to conceive and restore the estrous cycle; and the ability of farmers to detect estrus and insemination5. The last RP (age at first calving) is a measure of the female calf’s ability to grow and conceive at an early age.
To ensure the quality of the data of the RPs studied, biologically feasible limits were applied to prevent the use of misinformation (outliers), considering the values reported by the International Bull Evaluation Service (INTERBULL)21. Thus, the following records were excluded: days from calving to first service < 20 or > 365 d, when the number of services per conception was greater than 10, when the days open were <20 or > 365, when the intervals between calving were outside the range of 290 to 762 d and when the age at first calving was <18 or > 40 mo, the descriptive statistics of the variables studied are shown in Table 1. After quality control, the dataset consisted of 202,545 records for CFI and NSC; 194,816 records for DO; 139,901 for IBC and 103,467 for AFC. The pedigree files included between 103,467 and 202,545 animals, depending on the trait evaluated and included up to three generations of ancestors of animals with reproductive records.
Estimation of variance components
The variance components for CFI, NSC, DO and IBC were estimated with a repeatability animal model, separately for each characteristic to take advantage of as much information as possible. In the animal model, the availability of individual records allows the prediction of genetic values for all individuals in the population (even if they do not have phenotypic information but genealogical information). The genetic values are calculated with the best linear unbiased predictor, with a mixed equation model (Equation 1)13, where the random effect refers to the animal that has records and its ancestors, and the fixed effects are all those environmental effects, such as the year of birth14.
In order to control sources of variation other than those studied in the models of the RPs, the following were included as fixed effects: the subclasses of herd/year/season of calving or herd-year season of birth for AFC, with four seasons defined according to the month of calving (January to March, April to June, July to September and October to December); the level of production (classified into four levels for all farms from 1 to 4, seeking to assign cows to the quartile that describes their level of production within their herd/year/season of calving, with 1 being the one with the lowest production and 4 being the one with the highest production); the number of calving (grouping cows with four lactations or more in a single class) and age at calving. The animal and the cow’s permanent environment were included as random effects. In the case of AFC, there was no permanent environment.
The general animal model used was:
Where:
y=variable of interest (vector of records of: CFI, NSC, DO, IBC, AFC);
b= vector of fixed effects (herd-year-season of calving (hys) or herd-year season of birth (hysb) for AFC, production level (pl), number of calving (noc) and age at calving (ac));
u= vector of random effects (effect of animal and permanent environment when applicable);
X= incidence matrix that relates observations to fixed effects;
Z= incidence matrix that relates observations to random effects (animal and permanent environment);
e= vector of the effects of the error or residuals.
No transformations of the studied variables were performed because previous studies12 showed that they are not necessary. Estimates of variance components were made by means of restricted maximum likelihood22, using the BLUPF90 software 23.
Multivariate analyses
In order to estimate the genetic covariances and correlations for each pair of traits, bivariate analyses were performed using the models described above.
The general model used was:
where subindices 1 and 2 identify the pair of traits to be evaluated (CFI, NSC, DO, IBC, AFC) with their respective fixed and random effects.
Genetic parameters and correlations
Narrow-sense heritabilities (h2), repeatability (r) and genetic correlations24 between the RPs were obtained based on the estimates of the variance components using the BLUPF90 software 23. Standard deviations of the estimated parameters were approximated based on their expectations using the REMLF90 software 23.
Analysis by periods (additive genetic variance and genetic values)
Taking into consideration that the import of germplasm from different countries worldwide, especially from the United States and Canada, has been increasing in recent years, representing almost 80 % of the bulls used in the population studied, as well as changes in the values of the reproductive parameters of these countries (due to the introduction of genomic information in the evaluation and selection processes since 2009), data were grouped into three periods, each of four years (P1: 2006-2009, P2: 2010-2013 and P3: 2014-2017). In the first period, selection was considered to be based on pedigree, which uses information from the records of cows and their relatives. In the second period, the incorporation of genomic information into the selection began. In the third period, it is considered that genomic selection is established, and with it the possibility of increasing the accuracy of evaluations of low heritability traits.
Results and discussion
Reproductive parameters
The average for CFI was 71.5 d, which could indicate that the first inseminations are being carried out between 10 and 30 d after the restoration of the estrous cycle (21 d) and the subsequent voluntary waiting period (usually 40 d). In comparison, measurements made in different countries reported averages ranging from 70 to 93 d (Table 1).
Table 1: Descriptive statistics of reproductive parameters in Holstein populations, including those of the present study
| Author | Country | CFI | NSC | DO | IBC | AFC |
|---|---|---|---|---|---|---|
|
|
|
|
|
|
||
| 29 | USA | 87.6 ± 26 | - | 109.2 ± 38 | - | 33.6 ± 12.1 |
| 6 | USA | - | - | 131.5 | - | - |
| 26 | CAN | 80.2 | 1.66 | 108.4 | - | - |
| 34 | USA | - | - | - | - | 27.2 ± 3.3 |
| 36 | USA | 70.5 ± 26 | - | - | - | - |
| 36 | USA | 91.5 ± 40 | - | - | - | - |
| 31 | ITA | 84.5 ± 37 | 1.7 ± 0.1 | - | 413.5 ± 82 | - |
| 12 | ESP | 81 ± 28 | 1.9 ± 1.2 | 117 ± 57.1 | 400 ± 60 | - |
| 33 | MEX | - | - | - | 406 ± 67 | 27.6 ± 2.9 |
| 33 | USA | - | - | - | 401 ± 62 | 27.1 ± 3.1 |
| 28 | DNK | 81.3 ± 40 | 2.2 ± 1.5 | 133.3 ± 76 | 413.1 ± 76 | - |
| 18 | MEX | 73 ± 45 | 2.0 ± 1.3 | 101 ± 54 | 389 ± 57 | - |
| 25 | TUN | 93.2 ± 80 | 2.6 ± 1.7 | 150.9 ± 76 | 444.2 ± 102 | - |
| 19 | MEX | - | - | - | 418.6 ± 89 | - |
| 30 | IRN | 72.9 ± 35 | 2.1 ± 1.4 | 117.7 ± 64 | 393.9 ± 63 | - |
| 27 | COL | - | 1.6 ± 1.0 | 127.2 ± 77 | 410.3 ± 78 | - |
| 32 | CZE | - | - | - | 400 ± 59 | 25.3 ± 2.5 |
| Present study | MEX | 71.5 ± 23 | 2.7 ± 1.9 | 131.6 ± 74 | 413.6 ± 85 | 24.2 ± 2.7 |
CFI = calving to first insemination interval in days, NSC= number of services per conception, DO = days open, IBC= interval between calvings in days, AFC= age at first calving in months,
The results for NSC were 2.7 on average (Table 1), similar to the 2.55 reported in a population of this same breed (Holstein) from Tunisia25. Lower averages from 1.58 and up to 2 services per conception have been reported in Holstein populations of Spain12, Mexico18, Canada26 and Colombia27. The difference can be caused by the upper limits of 7 services established by the aforementioned authors, with respect to the 10 services considered in the estimates of this work.
For DO, the results showed 131.6 d, in accordance with the 131.5 d reported in a population of the United States6, of Denmark with 133.3 d28; or in Colombia with 127.2 d27 (Table 1). While lower averages were obtained in other studies from 101 d to 117 d18,29,30. In contrast, a value of 150 d was reported in Tunisia25. As in the traits discussed above, the differences between this study and those cited here could be due to the variation of the limits used for each population, as well as to postpartum health problems specific to each population studied29.
The average of IBC of 413 d coincides with other populations28,31 (Table 1), as well as reports ranging from 400 to 416 d19,27,32. On the other hand, lower averages of 389 d were reported in 2010 in a Holstein and Brown Swiss Mexican population under subtropical conditions18.
For AFC, the results were 24.15 mo (Table 1), similar to that reported in several studies, such as the 25.34 mo mentioned in the Czech Republic32, 27.1 in populations of the United States33,34, as well as 27.6 mo in another Mexican population33 of the Holstein association of Mexico that includes data from 1971 to 1995. In contrast, 33.6 mo were reported in a population of the United States29. This parameter shows about the female calf’s ability to grow and conceive at an early age5, which may be different for each population, since there is a cost-benefit ratio in achieving earlier ages at first calving11. That is, it is a voluntary and economic decision of the administrators of each population, because the age at first insemination determines AFC and the first is almost always an administrative decision, since the cost associated with developing the female calf faster can be important in some environments (countries and production systems)11,35.
The differences obtained in the RPs evaluated and those mentioned above illustrate the difficulty in estimating them and the lack of consensus on acceptable biological limits in the measurements of these traits3,5, which generates possible biases to the estimates and difficulties for comparison. Other issues such as the dependence of these traits on the conditions of each country26,27,30, the economic purposes of each farm12, as well as the conditions and management that vary from one herd to another35, may be generating the changes observed between countries.
Genetic parameters
Heritability ranged from 4 % to 9 % for the RPs studied, being higher for AFC (9 %), of 5 % for NSC, DO and IBC, and the lowest for CFI with 4 % (Table 2), similar to that previously reported for fertility traits12,26,27.
Table 2 Narrow-sense heritability and repeatability in Holstein populations, including those of the present study
| Author | Country | CFI | NSC | DO | IBC | AFC | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| h2 | r | h2 | r | h2 | r | h2 | r | h2 | ||
| 6 | USA | - | - | - | - | 0.05 | - | - | - | 0.05 |
| 26 | CAN | 0.03 | 0.08 | 0.03 | 0.07 | 0.05 | 0.10 | - | - | - |
| 31 | ITA | 0.06 | 0.14 | 0.03 | - | - | - | 0.07 | - | - |
| 12 | ESP | 0.05 | - | 0.02 | - | 0.04 | - | 0.04 | - | - |
| 28 | DNK | 0.09 | - | 0.03 | - | 0.07 | - | 0.07 | - | - |
| 25 | TUN | 0.03 | 0.13 | 0.03 | 0.03 | 0.04 | 0.14 | 0.06 | 0.15 | - |
| 19 | MEX | - | - | - | - | - | - | 0.01 | - | 0.03 |
| 30 | IRN | 0.06 | - | 0.05 | - | 0.08 | - | 0.07 | - | - |
| 27 | COL | - | - | 0.04 | 0.07 | 0.08 | 0.1 | 0.09 | 0.09 | - |
| 32 | CZE | - | - | - | - | - | - | 0.03 | 0.09 | 0.03 |
| PS | MEX | 0.04 (0.003) | 0.09 (0.003) | 0.05 (0.004) | 0.08 (0.003) | 0.05 (0.004) | 0.08 (0.003) | 0.05 (0.004) | 0.08 (0.003) | 0.09 (0.007) |
RP: reproductive parameter, CFI: calving to first insemination interval in days, NSC: number of services per conception, DO: days open, IBC: interval between calvings in days, AFC: age at first calving in months, h2: heritability, r: repeatability, standard error in parentheses, PS: present study.
The lowest heritability obtained was for CFI, with 4 %, similar to the 3 % reported in populations of Tunisia25 and Canada26, as well as 5 % of a Spanish population12. However, the results of the present study were lower than the 6 % reported by Italy31 and Iran30, as well as the 9 % estimated in Denmark28.
The estimated heritability for NSC was 5 %, consistent with that reported in an Iranian population30, in a Colombian27 with 4 %, and the 3 % reported in populations of Spain12, Tunisia25 and Canada26.
Regarding DO, the heritability is consistent with that reported in Canada26 and the United States6 of 5 %, as well as those estimated in Spain12 and Tunisia25 with 4 %; on the other hand, 7 % was reported in Denmark28 and 8 % estimated in populations of Colombia and Iran27,30.
For IBC, a heritability of 5 % was estimated, which was also similar to previous works done in Tunisia (6 %)25, Spain (4 %)12, Czech Republic (3 %)32 and lower than the 7 % in Italy31, Denmark28 and Iran30. The maximum heritability value reported for this reproductive parameter is 9 % in Colombia27, and the lowest value reported is 1 %, in the same study population over a period of time from 1998 to 200319.
The highest heritability obtained in the present study was for AFC, with 9 %, higher than the 5 % reported in one population of the United States6, or the 3 % reported in another Mexican population19 and in the Czech Republic32. Therefore, it would be expected that, in our population, the response to selection for this trait would have a better result, due to differences in heritabilities.
The repeatability estimate for NSC and DO was the same, slightly higher for CFI and IBC with 9 % (Table 2). Other values reported for the Holstein breed are similar to those obtained in the present study26,27,32.
The genetic parameters of the reproductive traits included in this study have relatively low values (heritability < 10 % and repeatability < 15 %)12,19,26, so, in addition to genetics, the environment has an important effect as a source of variation36. That is, other factors such as health29,36, nutritional aspects36, as well as administrative management issues (voluntary waiting period) by producers impact these traits6,26,29.
It is important to highlight that, although the heritabilities found in this study are low, genetic improvement can allow positive, permanent and cumulative benefits to be obtained in populations11,16,17.
The estimated phenotypic and genetic correlations were positive for almost all reproductive parameters (Table 3). The highest genetic correlations were found between DO and IBC (0.99), as well as high values of 0.84 and 0.88 for NSC with DO and IBC, respectively. For CFI with DO and IBC, an intermediate correlation (0.55 and 0.51) was found. For the other combinations of fertility traits, low correlations (from 0.09 to 0.22) were found.
Table 3 Genetic correlations above the diagonal (standard error), phenotypic correlations below the diagonal (number of records for each pair of traits)
| CFI | NSC | DO | IBC | AFC | |
|---|---|---|---|---|---|
| CFI | - | 0.13 (0.06) | 0.55 (0.05) | 0.51 (0.06) | 0.09 (0.07) |
| NSC | -0.05 (202,545) | - | 0.84 (0.02) | 0.88 (0.02) | 0.22 (0.08) |
| DO | 0.26 (192,527) | 0.77 (194,816) | - | 0.99 (0.002) | 0.09 (0.08) |
| IBC | 0.23 (138,277) | 0.78 (139,901) | 0.98 (137,158) | - | 0.12 (0.11) |
| AFC | 0.08 (103,467) | 0.01 (84,675) | 0.06 (80,467) | 0.07 (62,999) | - |
CFI = calving to first insemination interval in days; NSC= number of services per conception; DO = days open; IBC= interval between calvings in days; AFC= age at first calving in months.
All phenotypic correlations had a P<0.001.
In particular, the lowest estimated genetic correlation was for the association between CFI and NSC (0.13), similar to the 0.12 reported for these parameters in an Italian population30, and it was also the lowest estimated for another Mexican population18 with a value of 0.25. CFI and DO had a positive and moderate genetic association of 0.55, in contrast, in another population of Spain12 and Mexico18, a value of 0.82 was obtained for this association. The correlation estimator for CFI and IBC indicates that these two reproductive traits show a positive and moderate genetic association (0.51), and lower than another Mexican population (0.89)18.
The correlation between NSC and DO was positive and high (0.84), although lower than that reported in Spain12, Mexico18 and Colombia27 (0.94, 0.97, 0.98, respectively). A similar correlation value of 0.88 was found between NSC and IBC, close to the 0.89 reported in Spain12, lower than the 0.95 and 0.97 in Colombia27 and Mexico18, respectively; unlike the 0.61 reported in Italy30. High values of genetic correlation indicate the possibility of selecting for a trait, because as one can be decreased or increased, the same effect on the other is expected. In addition, the close relationship and dependence of the estrus detection rate and pregnancy between these measurements may explain these results, as well as the fact that the same genetic regions are encoding these traits29,37.
Between DO and IBC, high genetic correlation values (0.99) were found, identical to those reported in Spain12 and Iran30 and similar to the 0.98 reported in Colombia27. The results found are to be expected, because the IBC includes DO and the length of gestation29, as a consequence, any variation of days open should result in changes in the interval between calvings. Additionally, these two traits, being almost equivalent, suggest that they are controlled by the same genes or genetic regions, which is known as the pleiotropic effect37.
The age at first calving is influenced by the environment and management practices common to female calves and heifers born in the same period (year or year-season)6, which was corroborated by the low correlations found between it and the other RPs (Table 3), so these results suggest genetic and phenotypic independence among them.
Additive genetic variance
The results of the estimates of variances by period can be divided into two groups of comparisons P1 vs P2 and P2 vs P3 (Table 4). In P1 vs P2, for additive genetic variances, was observed similar values in CFI (12.48-11.17), NSC (0.15-0.16), DO (229.8-214.6) and IBC (478-466.7), except for AFC (1.23-0.19) which showed a significant decrease, possibly due to the selection on milk production during this period, because as a cow becomes pregnant, it will begin its productive life.
Table 4 Additive genetic variance and heritability for five reproductive parameters in three periods of time (P1: 2006-2009, P2: 2010-2013 and P3: 2014-2017)
| Reproductive parameter | Period | Number of records | σ2 a ± SE | h 2 |
|---|---|---|---|---|
| CFI | P1:06-09 | 71,667 | 12.48 ± 2.2 | 0.03 |
| P2:10-13 | 63,952 | 11.17 ± 2.0 | 0.03 | |
| P3:14-17 | 43,233 | 19.88 ± 3.0 | 0.06 | |
| NSC | P1:06-09 | 71,667 | 0.15 ± 0.02 | 0.05 |
| P2:10-13 | 63,952 | 0.16 ± 0.02 | 0.05 | |
| P3:14-17 | 43,233 | 0.10 ± 0.02 | 0.04 | |
| DO | P1:06-09 | 69,417 | 229.8 ± 35.32 | 0.04 |
| P2:10-13 | 62,783 | 214.6 ± 33.87 | 0.04 | |
| P3:14-17 | 42,790 | 194.8 ± 25.66 | 0.05 | |
| IBC | P1:06-09 | 54,005 | 478.0 ± 51.1 | 0.07 |
| P2:10-13 | 48,388 | 466.7 ± 52.5 | 0.07 | |
| P3:14-17 | 25,801 | 266.8 ± 53.9 | 0.05 | |
| AFC | P1:06-09 | 31,542 | 1.23 ± 0.12 | 0.22 |
| P2:10-13 | 34,094 | 0.19 ± 0.04 | 0.04 | |
| P3:14-17 | 31,222 | 0.15 ± 0.03 | 0.04 |
CFI = calving to first insemination interval in days, NSC= number of services per conception, DO = days open, IBC= interval between calving in days, AFC= age at first calving in months, σ2 a= additive genetic variance, SE= standard error, h2= narrow-sense heritability.
In P2 vs P3, an increase in CFI (11.17-19.88) was observed in additive genetic variance, which suggests genetic diversity and the larger it is, the greater the opportunity for response to selection38. In contrast, decreases were observed for NSC (0.16-0.10), DO (214.6-194.8) and IBC (466.7-266.8), while AFC was similar to the previous period (0.19-0.15). The decrease in the additive genetic variances of NSC, DO and IBC may be due to a selective pressure of these traits.
Genetic values
The changes over time between groups P1 vs P2, and P2 vs P3 are shown in Figures 1a and 1b, where it can be seen how, in general, genetic trends were similar between P1 and P2, decreasing in P3. These results are in line with the farmers’ criteria for choosing semen, who, during the first two periods, emphasized the volume of milk, which decreased in P3, while in case of reproductive traits, there were no changes in P1 and P2, but a positive change was observed in P3 [Durán C., 2022, unpublished data].
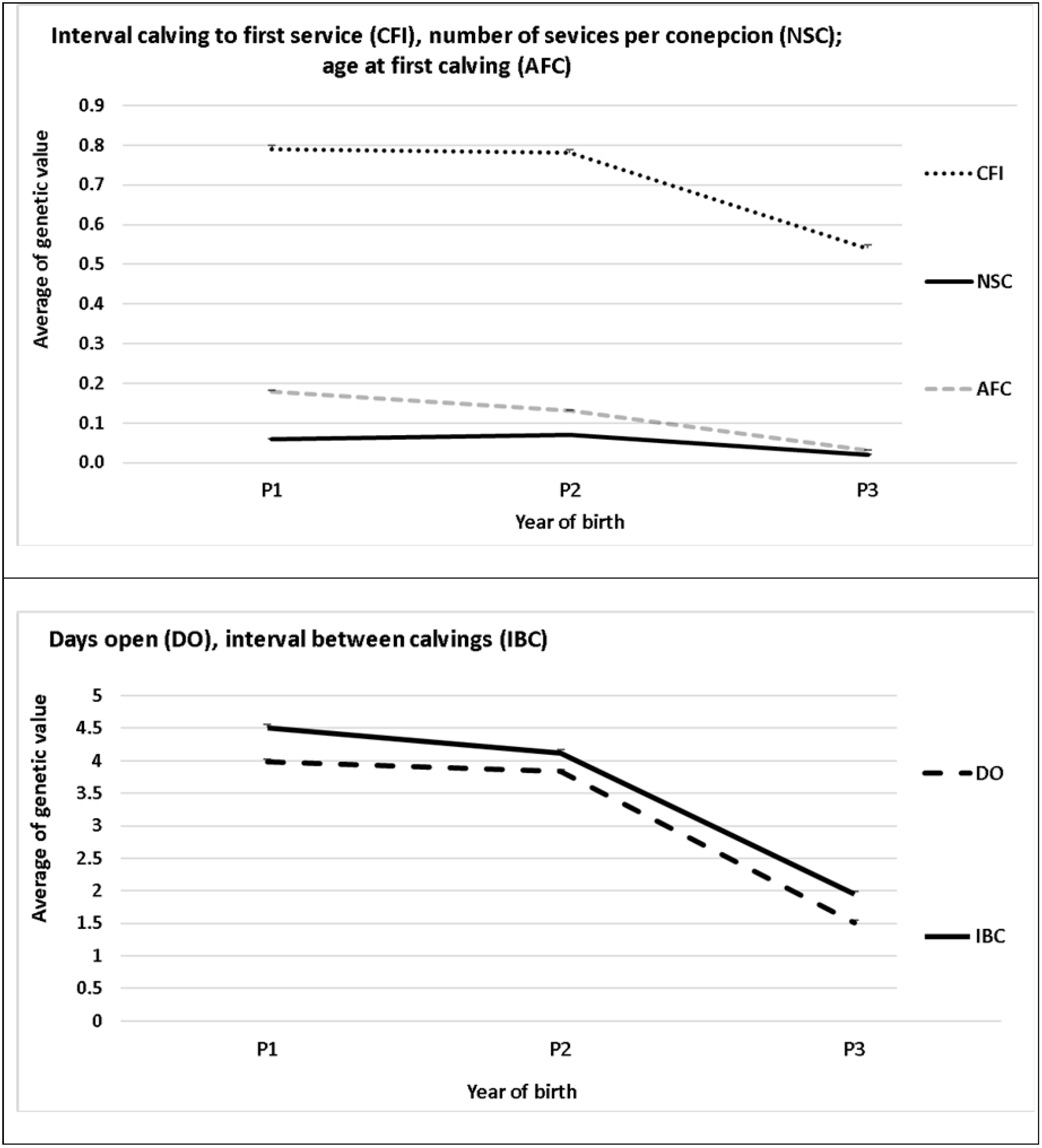
Figure 1 Genetic trends for CFI (=IPPS), NSC and AFC (=EPP) (a) and for DO (=DAB) and IBC (=IEP) (b) in three periods of time (P1: 2006-2009, P2: 2010-2013 and P3: 2014-2017)
For group II (P2 vs P3), a decrease in average genetic values can be observed for CFI, DO, IBC and AFC, but not for NSC. This reduction in genetic values is desirable because what is sought in these fertility traits studied is the decrease.
These changes in fertility traits could possibly be attributed to the import and use of germplasm from the United States since 195039, which has improved fertility traits since 200317 and uses genomic selection since 200916.
For NSC, no changes were observed in genetic values, probably because this characteristic depends significantly on insemination techniques, which do not depend on the population of origin of the germplasm.
The results of this study show how the incorporation of genomic information in the selection processes of the countries of origin of imported germplasm has apparently had a positive effect on the population (Figures 1a and 1b), regardless of the low heritability and difficulty of measuring fertility traits16,17.
Conclusions and implications
The estimates of heritability and repeatability of the reproductive parameters obtained in the present study were low and similar to previous reports in the breed. However, the existing additive genetic variation is sufficient to expect favorable and cumulative (although slow) results in the selection of these traits. The estimated correlations between the PRs were positive, highlighting that they were high between NSC, DO and IBC, in particular the correlation between DO and IBC, with a correlation of 0.99, which suggests that it would be advisable to select only for one parameter, since it could be considered that its effects are controlled by the same genetic regions. The results of the estimates of additive genetic variances and genetic values in different periods show the existence of changes in the population over time, possibly derived from the implementation of genomic selection in germplasm supplier countries, such as Canada and the United States.
Acknowledgements and conflict of interest
This paper is part of the requirements for obtaining a Doctoral degree of CDA at the Posgrado en Ciencias Biológicas, of the Universidad Nacional Autónoma de México (UNAM). Special thanks to M. Sc. Enoc Mejía, to M. Sc. José Cortés and to MVZ Marisol Morales for their advice on the management of databases, as well as to the Holstein Association of Mexico for the information provided. The study was funded by the Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias with the project No. SIGI: 15352034772. The Consejo Nacional de Ciencia y Tecnología is also thanked for scholarship no. 631475 provided to CDA. The authors declare no conflict of interest.
REFERENCES
1. Flaws JA, Spencer TE. Female reproduction, content and volume overview. In: Academic Press. Encyclopedia of reproduction. 2nd ed. USA: Academic Press Books - Elsevier; 2018: V2:1-2. [ Links ]
2. Walsh SW, Williams EJ, Evans ACO. A review of the causes of poor fertility in high milk producing dairy cows. Anim Reprod Sci 2011;123(3-4):127-138. [ Links ]
3. Banos G. Survey on genetic evaluation procedures for functional traits in cattle in various countries. Interbull Bulletin 1996;(12). [ Links ]
4. VanRaden PM, Sanders AH, Tooker ME, Miller RH, Norman HD, Kuhn MT, et al. Development of a national genetic evaluation for cow fertility. J Dairy Sci 2004;87(7): 2285-2292. [ Links ]
5. Jorjani H. International genetic evaluation for female fertility traits. Interbull Bulletin 2006;(34):57. [ Links ]
6. Seykora AJ, McDaniel BT. Heritabilities and correlations of lactation yields and fertility for Holsteins. J Dairy Sci 1983;66(7):1486-1493. [ Links ]
7. Hageman WHD, Shook GE, Taylor WJ. Reproductive performance in genetic lines selected for high or average milk yield. J Dairy Sci 1991;74:4366-4376. [ Links ]
8. Peñagaricano F. Genetics and genomics of dairy cattle. In: Academic Press. Animal Agriculture. Texas, USA: Academic Press Books - Elsevier; 2020:101-119. [ Links ]
9. Van Arendonk JA, Hovenier R, De Boer W. Phenotypic and genetic association between fertility and production in dairy cows. Livest Prod Sci 1989;21(1):1-12. [ Links ]
10. Vollema AR. Selection for longevity in dairy cattle [doctoral thesis]. Wageningen, The Netherlands: Wageningen University and Research; 1998. [ Links ]
11. Pritchard T, Coffey M, Mrode R, Wall E. Genetic parameters for production, health, fertility and longevity traits in dairy cows. Animal 2012;7(1):34-46. [ Links ]
12. González-Recio O, Alenda R. Genetic parameters for female fertility traits and a fertility index in Spanish dairy cattle. J Dairy Sci 2005;88:3282-3289. [ Links ]
13. Henderson CR. Applications of linear models in animal breeding. Ontario, Canada: University of Guelph, Guelph; 1984. [ Links ]
14. Mrode RA. Linear models for the prediction of animal breeding values. SRUC UK: Cabi; 2005. [ Links ]
15. Meuwissen TH, Hayes BJ, Goddard ME. Prediction of total genetic value using genome-wide dense marker maps. Genetics 2001;157:1819-1829. [ Links ]
16. Wiggans GR, Cole JB, Hubbard SM, Sonstegard TS. Genomic selection in dairy cattle: the USDA experience. Annu Rev Anim Biosci 2017;5:309-327. [ Links ]
17. García-Ruiz A, Cole JB, VanRaden PM, Wiggans GR, Ruiz-López FJ, Van Tassell CP. Changes in genetic selection differentials and generation intervals in US Holstein dairy cattle as a result of genomic selection. Proc Natl Acad Sci 2016;113(28):E3995-E4004. [ Links ]
18. Ríos-Utrera A, Calderón-Robles R, Rosete-Fernández J, Lagunes-Lagunes J. Análisis genético de características reproductivas de vacas Holstein criadas en un ambiente subtropical. Agronomía Mesoamericana 2010;21:245-253. [ Links ]
19. Montaldo HH, Castillo-Juárez H, Valencia-Posadas M, Cienfuegos-Rivas EG, Ruiz-López FJ. Genetic and environmental parameters for milk production, udder health, and fertility traits in Mexican Holstein cows. J Dairy Sci 2010;93(5):2168-2175. [ Links ]
20. Montaldo HH, Pelcastre-Cruz A, Castillo-Juárez H, Ruiz-López FJ, Miglior F. Genotype×environment interaction for fertility and milk yield traits in Canadian, Mexican and US Holstein cattle. Span J Agric Res 2017;15(2):e0402-e0402. [ Links ]
21. INTERBULL. International Bull Evaluation Service. National genetic evaluation forms provided by countries. 2020. https://interbull.org/ib/geforms Accessed Sep 1, 2020. [ Links ]
22. Patterson HD, Thompson R. Recovery of interblock information when block sizes are unequal. Biometrika 1971;58:545-554. [ Links ]
23. Misztal I, Aguilar I, Legarra A, Vitezica Z, Tsuruta S, Lourenco D. Manual for BLUPF90 family of programs. 2014. http://nce.ads.uga.edu/wiki/lib/exe/fetch.php? media=blupf90_all7.pdf Accessed Nov 1, 2020. [ Links ]
24. Falconer DS. Introduction to quantitative genetics. 2nd ed. New York, USA: Longman Scientific & Technical; 1981. [ Links ]
25. M’hamdi N, Aloulou R, Brar SK, Bouallegue M, Ben Hamouda M. Phenotypic and genetic parameters of reproductive traits in Tunisian Holstein cows. Biotechnol Anim Husb 2010;26:297-307. [ Links ]
26. Hayes JF, Cue RI, Monardes HG. Estimates of repeatability of reproductive measures in Canadian Holsteins. J Dairy Sci 1992;75(6):1701-1706. [ Links ]
27. Zambrano JC, Echeverri J. Genetic and environmental variance and covariance parameters for some reproductive traits of Holstein and Jersey cattle in Antioquia (Colombia). Rev Bras Zootec 2014;43(3):132-139 [ Links ]
28. Sun C, Madsen P, Lund M, Zhang Y, Nielsen US, Su G. Improvement in genetic evaluation of female fertility in dairy cattle using multiple-trait models including dairy yield traits. J Anim Sci 2010;88:871-878. [ Links ]
29. Fonseca FA, Britt JH, McDaniel BT, Wilk JC, Rakes AH. Reproductive traits of Holsteins and Jerseys. Effects of age, milk yield, and clinical abnormalities on involution of cervix and uterus, ovulation, estrous cycles, detection of estrus, conception rate, and days open. J Dairy Sci 1983;66(5):1128-1147. [ Links ]
30. Ghiasi H, Pakdel A, Nejati-Javaremi A, Mehrabani-Yeganeh H, Honarvar M, González-Recio O, et al. Genetic variance components for female fertility in Iranian Holstein cows. Livest Sci 2011;139(3):277-280. [ Links ]
31. Biffani S, Canavesi R, Samore AB. Estimates of genetic parameters for fertility traits of Italian Holstein Friesian cattle. Stocarstvo 2005;59:145-153. [ Links ]
32. Brzáková M, Zavadilová L, Přibyl J, Pešek P, Kašná E, Kranjčevičová A. Estimation of genetic parameters for female fertility traits in the Czech Holstein population. Czech J Anim Sci 2019;64(5):199-206. [ Links ]
33. Cienfuegos-Rivas EG, Blake RW, Oltenacu PA, Castillo-Juarez H. Fertility responses of Mexican Holstein cows to US sire selection. J Dairy Sci 2006;89(7):2755-2760. [ Links ]
34. Castillo-Juarez H, Oltenacu PA, Blake RW, McCulloch CE, Cienfuegos-Rivas EG. Effect of herd environment on the genetic and phenotypic relationships among milk yield, conception rate, and somatic cell score in Holstein cattle. J Dairy Sci 2000;83(4): 807-814. [ Links ]
35. Garcia-Peniche TB, Cassell BG, Pearson RE, Misztal I. Comparisons of Holsteins with Brown Swiss and Jersey cows on the same farm for age at first calving and first calving interval. J Dairy Sci 2005;88(2):790-796. [ Links ]
36. Weigel KA, Rekaya R. Genetic parameters for reproductive traits of Holstein cattle in California and Minnesota. J Dairy Sci 2000;83(5):1072-1080. [ Links ]
37. Bulmer MG. The mathematical theory of quantitative genetics. New York, USA: Clarendon Press, Oxford; 1980. [ Links ]
38. Woolliams JA, Oldenbroek JK. Genetic diversity issues in animal populations in the genomic era. In Oldenbroek JK editor. Genomic management of animal genetic diversity. Wageningen, The Netherlands: Wageningen Academic Publishers; 2017:13-45. [ Links ]
39. Cienfuegos-Rivas EG, Oltenacu PA, Blake RW, Schwager SJ, Castillo-Juarez H, Ruiz FJ. Interaction between milk yield of Holstein cows in Mexico and the United States. J Dairy Sci 1999;82(10):2218-2223. [ Links ]
Received: May 30, 2022; Accepted: January 23, 2023
 Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
