










Services on Demand
Journal
Article
 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista mexicana de ciencias pecuarias
On-line version ISSN 2448-6698Print version ISSN 2007-1124
Rev. mex. de cienc. pecuarias vol.14 n.1 Mérida Jan./Mar. 2023 Epub Mar 24, 2023
https://doi.org/10.22319/rmcp.v14i1.6200
Notas de investigación
Caracterización del virus de la diarrea viral bovino subtipo 1b aislado de un caso de la enfermedad de las mucosas
aComisión México-Estados Unidos para la prevención de fiebre Aftosa y otras enfermedades exóticas de los animales, Carretera México-Toluca Km 15.5 Piso 4 Col. Palo Alto. Cuajimalpa de Morelos. 05110. Ciudad de México. México.
bCentro Nacional de Servicios de Constatación en Salud Animal (CENAPA), Morelos, México.
El virus de la diarrea viral bovina (VDVB) genera pérdidas significativas en la producción de bovinos. Se reporta sobre un caso fatal de la enfermedad de las mucosas en un toro de dos años. Para la detección del agente causal, se analizaron muestras de lesiones, de sangre completa y de heces mediante el RT-PCR, PCR, ELISA y aislamiento viral. Se obtuvo amplificación positiva por RT-PCR en muestras de sangre para el virus de la diarrea viral bovina (VDVB). El aislamiento viral de las muestras de las lesiones confirmó que el VDVB fue el agente causal de las manifestaciones clínicas. Una caracterización genética basada en el análisis filogenético de tres secuencias parciales identificó la presencia del subgenotipo 1b del VDVB en las muestras analizadas. El animal muestreado manifestaba signos clínicos indicando que tenía la enfermedad de las mucosas, lo cual sugiere que sufría de una infección persistente (IP) por VDVB. Este hallazgo resalta la importancia de establecer programas de control de VDVB basados en probar la presencia de IP en el ganado de México.
Palabras clave Virus diarrea viral bovina; Ganado; Enfermedad de las mucosas; infección persistente; México
This report describes a fatal case of mucosal disease in a two-year-old bull. For causal agent detection, scab, whole blood, and feces samples were tested by RT-PCR, PCR, ELISA, and viral isolation. RT-PCR positive amplification was obtained in blood samples for bovine viral diarrhea virus (BVDV). Viral isolation from the scab samples confirmed BVDV as the causative agent of the clinical manifestations. Subsequently, genetic characterization based on phylogenetic analysis of three partial sequences revealed the presence of BVDV subgenotype 1b in analyzed samples. Due to the development of clinical manifestation named mucosal disease, these findings suggest the detection of BVDV persistently infected (PI) bull; therefore, these results demonstrate the importance of establishing BVDV control programs that rely on testing the presence of PI in cattle from Mexico.
Key words Bovine viral diarrhea virus; Cattle; Mucosal disease; Persistent infection; Mexico
La diarrea viral bovina (DVB) sigue siendo una de las enfermedades endémicas más comunes en el ganado bovino y otros rumiantes en todo el mundo. Esta enfermedad provoca impactos económicos significativos en la industria ganadera debido a sus efectos negativos sobre la reproducción y las condiciones de salud en el ganado1,2. La causa de la DVB es un virus de ARN monocatenario y de cadena positiva denominado virus de la diarrea viral bovina (VDVB); pertenece a la familia Flaviviridae dentro del género Pestivirus. En la actualidad, el VDVB se clasifica en tres especies: Pestivirus A (virus de la diarrea viral bovina 1, VDVB-1); Pestivirus B (virus de la diarrea viral bovina 2, VDVB-2); y Pestivirus H (pestivirus tipo HoBi). Cada especie se segrega en subgenotipos3. El Pestivirus A se subdivide en hasta 21 subgenotipos (del 1a hasta el 1u), y el Pestivirus B y el Pestivirus H se subdividen en cuatro subgenotipos cada uno (del a hasta el d)4.
También las cepas de VDVB se clasifican en biotipos citopáticos (CP) y no citopáticos (NCP) según su efecto sobre la replicación y los cambios morfológicos que inducen en el cultivo celular. Esta clasificación es relevante porque la citopatogenicidad in vitro no se relaciona con la citopatogenicidad in vivo. Las cepas NCP son predominantes en el campo y están involucradas en la mayoría de los casos de infección natural y de infecciones persistentes. Las cepas CP son poco comunes y se aíslan casi exclusivamente de una forma mortal de DVB llamada enfermedad de las mucosas (EM)5.
La infección por VDVB se caracteriza por manifestaciones clínicas que incluyen trastornos respiratorios, gastrointestinales y reproductivos. Las fallas reproductivas como los abortos, la momificación, la muerte fetal, los defectos congénitos y el nacimiento de animales persistentemente infectados (PI) generan grandes pérdidas económicas6.
Los animales PI son de particular preocupación porque pueden, por medio de la infección transplacentaria con una cepa de VDVB NCP, infectar a un embrión durante los primeros 125 días de gestación. Los animales nacidos de una madre con una PI adquieren una tolerancia inmunológica hacia la cepa de VDVB infectante y desarrollan una infección persistente. Por lo tanto, en un ternero con una PI congénita no se inducirá una respuesta inmune por anticuerpos o células T contra el virus7. Los animales PI son una fuente permanente del VDVB, ya que eliminan de por vida al virus en secreciones corporales como las nasales, orales, leche, orina, heces y en semen. Por lo tanto, juegan un papel esencial en la patogénesis y epidemiología de la DVB8.
Los terneros nacidos con una PI parecen animales normales, aunque a veces son débiles. Sin embargo, se caracterizan por tasas de crecimiento reducidas, y sufren de la inmunosupresión y de tasas altas de mortalidad2. Tanto la tasa de mortalidad como la de morbilidad incrementan en los animales con una PI debido a un aumento en su susceptibilidad a otras enfermedades. Mueren comúnmente por la neumonía o la EM. La mayoría de los terneros PI mueren por la EM entre los 6 y los 24 meses de edad9,10. Sin embargo, hay reportes de bovinos PI que alcanzan los 3, 5 y hasta 7 años, lo que implica un período largo de diseminación viral2,11,12.
La EM es una condición fatal esporádica restringida al ganado PI. Ocurre cuando el VDVB NCP causante de la PI muta a una CP como resultado de un evento de recombinación, o cuando el animal PI tiene una coinfección con una cepa CP del VDVB antigénicamente homóloga (13,14. Por lo tanto, se pueden encontrar ambos biotipos de manera consistente en animales con EM15,16. Un cuadro de EM resulta en la muerte del animal dentro de las dos semanas posteriores al inicio de los signos clínicos. Las principales lesiones que se encuentran en casos de EM son erosiones y ulceraciones extensas a lo largo del tracto gastrointestinal17. Sin embargo, existen reportes de animales con EM que manifestaron un inicio tardío (varios meses después de la infección inicial) de los signos18. Otros signos clínicos incluyen la anorexia, la fiebre, la deshidratación, la diarrea, la dermatitis, la necrosis del tejido linfoide, y un mal estado corporal19.
El presente reporte describe el inicio de EM en un toro de dos años de edad con signos clínicos severos; es posible que este sea la primera descripción de una IP en ganado en México. Se recibió la notificación del animal en junio del 2021. Al momento del reporte, el curso del cuadro tenía 15 días e involucraba signos clínicos como la anorexia, la depresión, el ptialismo, diarrea hemorrágica severa, la deshidratación, secreción nasal y ulceración profunda y extensa en el hocico, las narinas, los labios, las encías y el paladar duro (Figuras 1, 2 y 3). El animal afectado pertenecía a una granja tradicional de traspatio ubicada en Texcoco, Estado de México, México. La granja tenía cuatro bovinos, cuatro caballos, seis perros y tres cerdos, aparentemente sanos al momento del informe. Antes del evento, no se había registrado manifestaciones clínicas similares en bovinos, o cualquier otro animal doméstico, en otras fincas en el área circundante. Según el propietario, no hubo movilización de animales entre las fincas cercanas, y no se introdujeron nuevos animales en la granja antes que el animal comenzara con la signología clínica.

Figura 1 Toro de dos años de edad con enfermedad de las mucosas en donde se muestran lesiones erosivas en la descarga nasal, y ulcerizacíon extensiva en el hocico y las narinas
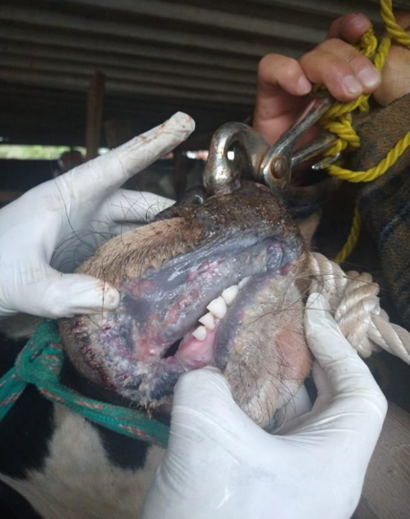
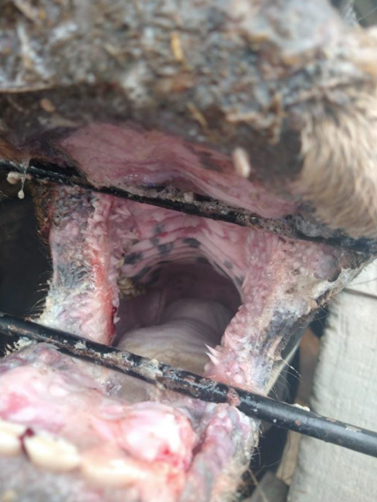
Se colectaron muestras de las lesiones cutáneas, muestras de sangre y de heces. Estas se remitieron para diagnóstico al Laboratorio de Inmunología, Biología Celular y Molecular de la Comisión México-Estados Unidos para la Prevención de la Fiebre Aftosa y Otras Enfermedades Exóticas de los Animales (CPA). El reporte de caso fue identificado con el número CPA-0861-21. Para el diagnóstico diferencial se consideraron las principales enfermedades vesiculares del ganado bovino, como son la fiebre aftosa (FA), la estomatitis vesicular (EV), la fiebre catarral maligna (FCM) y la DVB. No se detectó ni la FA ni la EV mediante RT-PCR, ELISA o el aislamiento del virus por cultivo celular. De igual forma, el virus del FCM no fue detectado por PCR.
El VDVB se aisló de las muestras de las lesiones y se obtuvo una amplificación positiva de las muestras de sangre completa mediante RT-PCR. El aislado de VDVB fue remitido al Laboratorio de Biología Molecular del Centro Nacional de Servicios de Diagnóstico en Salud Animal (CENASA) para su secuenciación parcial. Se secuenciaron la 5'UTR, Npro y E2 del VDVB, las cuales se depositaron en GenBank (números de acceso OM812936, OM812937 y OM812938, respectivamente). Se realizó un análisis filogenético basado en las regiones 5'UTR, Npro y E2. Las secuencias parciales de 5'UTR (360 pb), Npro (504 pb) y E2 (1,482 pb) obtenidas en este estudio se compararon con las cepas de referencia de VDVB para caracterizar el aislado de VDVB. Para las secuencias 5'UTR y Npro se infirió la historia evolutiva utilizando el método de máxima verosimilitud con un modelo de sustitución de dos parámetros de Kimura20, mientras para la E2 se usó un modelo de sustitución de tres parámetros de Tamura21. Ambos modelos se implementaron con el MEGA7 software usando 1,000 “bootstraps” de arranque cada una (Figura 4). Se modelaron las diferencias de velocidad evolutiva entre los sitios con una distribución gamma discreta con dos categorías; algunos sitios no mostraron variación evolutiva para las secuencias Npro y E2.
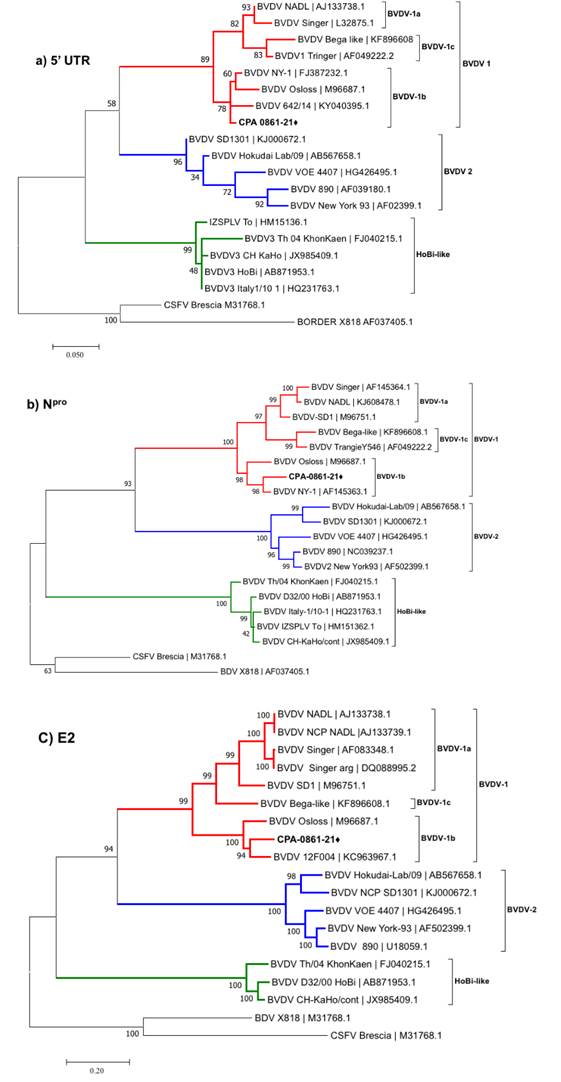
Se llevó a cabo la inferencia filogenética por medio del programa MEGA 7 siguiendo el método de verisimilitud máxima. Se hicieron 1,000 réplicas de “bootstraps”. Las secuencias de referencias se identifican por su número de acceso de GenBank. Las secuencias de nucleótidos identificadas en el presente estudio se identifican con el símbolo "⧫"
Figura 4 Árbol filogenético basado en las secuencias de las regiones 5'UTR (a), Npro (b) y E2 (c).
La DVB continúa siendo una enfermedad de preocupación significativa para la industria ganadera por su impacto económico asociado principalmente con los trastornos reproductivos2. Dependiendo de la etapa de gestación en la que se adquiere la infección por una cepa NCP del VDVB puede resultar en el nacimiento de terneros PI inmunotolerante. Estos animales son constantemente virémicos, y el VDVB se propaga a través de la mayoría de los órganos del animal pero sin que se desarrollen lesiones aparentes22. En consecuencia, el ganado PI mantiene la replicación y excreción viral de por vida en todas las secreciones corporales23. Por lo tanto, el ganado PI representa la principal fuente de transmisión y mantenimiento del VDVB dentro y entre hatos. Los VDVB NCP también pueden transmitirse por medio de ganado con una infección aguda y por fómites como material quirúrgico y de manejo contaminado, la revisión rectal, los sueros bovinos utilizados en la transferencia de embriones y producción de vacunas, el semen infectado y las vacunas contaminadas24-27.
Las infecciones por VDVB afectan de manera directa la fertilidad de los animales PI. Los toros PI pueden producir semen de calidad aceptable, pero de fertilidad baja asociada con anomalías en los espermatozoides y baja motilidad28. En hembras PI la función ovárica disminuye por medio de la hipoplasia y la reducción de las ovulaciones29. Así mismo, toros y vacas PI pueden engendrar crías PI aparentemente normales, los cuales recirculan el VDVB en animales susceptibles en el hato30.
La exposición continua de animales sanos al VDVB que viene de un animal con una IP puede perpetuar las infecciones por VDVB31. Como consecuencia, puede surgir la infertilidad del rebaño, la inmunosupresión y la generación de terneros PI32. Las infecciones agudas con los VDVB NCP comprometen la fertilidad del rebaño al producir un crecimiento folicular retardado y reducido33, la ovitis intersticial difusa 34, y fallas en la concepción al impedir la implantación del embrión22. También puede generar la muerte embrionaria antes del día 79 de gestación en vacas preñadas, o malformaciones congénitas entre los días 79 y 15035.
En áreas donde se implementan medidas adecuadas para el control del VDVB, la prevalencia estimada de animales con una IP está entre el 1-2%1. El presente caso es de importancia ya que no hay reportes de brotes de EM o su presencia en la población bovina mexicana. Además, se desconoce la proporción actual de terneros PI en el país. Actualmente, información limitada sobre caracterización genética y prevalencia del VDVB en México se ha comenzado a analizar (36.
El presente caso describe la presentación de EM causado por el VDVB-1b en un toro de carne en el que predominaron las lesiones ulcerativas en el tracto gastrointestinal. El VDVB-1b se define actualmente como la cepa más común en el campo. De hecho se considera el subgenotipo predominante a nivel mundial, seguido por el 1a y 1c4. Al VDVB-1b lo han descrito como la cepa más prevalente en terneros PI37. La presente identificación del subgenotipo 1b del VDVB coincide con un estudio previo que describe el VDVB-1b como un virus endémico que circula en el ganado bovino mexicano, junto con el 1a, 1c y 2a38. En conjunto con el presente caso de estudio, estos estudios representan un acercamiento inicial al VDVB en México.
El DVB sigue siendo una enfermedad no regulada en el país y no existen estrategias de control o medidas de prevención al nivel oficial. En consecuencia, los protocolos de vacunación se basan en procedimientos voluntarios y las medidas de seguimiento y de bioseguridad se implementan en función al conocimiento del DVB de los productores. Se confirmó que el toro evaluado en este caso clínico estaba infectado con el VDBV-1b además de manifestar signos sugiriendo un caso de EM. El toro detectado como positivo a VDVB pertenece a una granja donde se aplican escasas medidas sanitarias, las cuales no incluyen un protocolo de vacunación contra el VDVB.
Escenarios como la anterior son comunes en México, lo cual resalta la falta de medidas de control contra el DVB en el país. El presente caso de estudio confirmó la presencia del VDVB-1b en bovinos mexicanos y coincide con estudios anteriores38. El animal estudiado mostraba un cuadro grave de EM, destacando la importancia por subdiagnóstico de animales PI y, desde luego, el estado epidemiológico del VDVB. Informes sobre casos de DVB impulsarán el desarrollo de estrategias de control que permitan a los productores detectar el VDVB y eliminar a los terneros PI de sus rebaños. La vacunación es un elemento vital dentro de una estrategia de control del VDVB; sin embargo, se debe de escoger la vacuna más adecuada para brindar protección contra el VDVB circulante. Por ejemplo, en México ya se agregó el VDVB-1b como antígeno en una vacuna comercial. Aunque la vacunación es importante en el control del DVB, debe de formar parte de programas amplios para el control de DVB. El hallazgo de VDVB-1b en un toro no vacunado demuestra el papel crucial de la bioseguridad y la vigilancia de enfermedades para mitigar los efectos de las infecciones por VDVB en las poblaciones de ganado.
Literature cited
1. Houe H. Epidemiological features and economical importance of bovine virus diarrhoea virus (BVDV) infections. Vet Microbiol 1999;64(2-3):89-107. doi: 10.1016/s0378-1135(98)00262-4. Erratum in: Vet Microbiol 2003;93(3):275-6. [ Links ]
2. Houe H. Economic impact of BVDV infection in dairies. Biologicals 2003;31(2):137-43. doi: 10.1016/s1045-1056(03)00030-7. [ Links ]
3. Smith DB, Meyers G, Bukh GJ, Gould EA, Monath T, Scott-Muerhoff A, et al. Proposed revision to the taxonomy of the genus Pestivirus, family Flaviviridae. J General Virol 2017;98(8), 2106-2112. https://doi.org/10.1099/jgv.0.000873. [ Links ]
4. Yesilbag K, Alpay G, Becher P. Variability and global distribution of subgenotypes of bovine viral diarrhea virus. Viruses 2017;9(6):128. doi: 10.3390/v9060128. [ Links ]
5. Neill J. Molecular biology of bovine viral diarrhea virus. Biologicals 2013;41:2-7. doi: 10.1016/j.biologicals.2012.07.002. [ Links ]
6. Houe H, Lindberg A, Moenning V. Test strategies in bovine viral diarrhea virus control and eradication campaigns in Europe. J Vet Diagn Invest 2006;18(5):427-36. doi: 10.1177/104063870601800501. [ Links ]
7. Meyers G, Thiel HJ. Molecular characterization of pestiviruses. Adv Virus Res 1996; 47:53-118. doi: 10.1016/s0065-3527(08)60734-4. [ Links ]
8. Brodersen BW. Bovine viral diarrhea virus infections: manifestations of infection and recent advances in understanding pathogenesis and control. Vet Pathol 2014;51(2):453-64. doi: 10.1177/0300985813520250. [ Links ]
9. Odeón AC, Leunda MR, Faverín C, Boynak N, Vena MM, Zabal O. In vitro amplification of BVDV field strains isolated in Argentina: effect of cell line and culture conditions. Rev Argent Microbiol 2009;41:79-85. [ Links ]
10. Uzal FA, Platnner BL, Hostetter JM. Alimentary system in pathology of domestic animals. In: Maxie, MG, editor. Jubb, Keneddy and Palmers pathology of domestic animals.;6th ed. St. Louis, Missouri: Academic Press Inc; 2016:122-130. [ Links ]
11. Brock KV, Grooms DL, Ridpath JF, Bolin SR. Changes in levels of viremia in cattle persistently infected with bovine viral diarrhea virus. J Vet Diagn Invest 1998;10:22-26. [ Links ]
12. Bedekovic T, Lemo N, Lojkic I, Cvetnicz Z, Cac Z, Madic J. Bovine viral diarrhoea: A seven year old persistently infected cow - a case report. Veterinarski Arch 2012;82 (6):637-643. [ Links ]
13. Brownlie J. The pathways for bovine virus diarrhoea virus biotypes in the pathogenesis of disease. Arch Virol Suppl 1991;3:79-96. doi: 10.1007/978-3-7091-9153-8_10. [ Links ]
14. Tautz N, Thiel HJ. Cytopathogenicity of pestiviruses: cleavage of bovine viral diarrhea virus NS2-3 has to occur at a defined position to allow viral replication. Arch Virol 2003;148(7):1405-12. doi: 10.1007/s00705-003-0106-9. [ Links ]
15. Bolin SR, McClurkin AW, Cutlip RC, Coria MF. Severe clinical disease induced in cattle persistently infected with noncytopathic bovine viral diarrhea virus by superinfection with cytopathic bovine viral diarrhea virus. Am J Vet Res 1985;46(3):573-6. [ Links ]
16. Kummerer B, Tautz MN, Becher P, Thiel H, Meyers G. The genetic basis for cytopathogenicity of pestiviruses. Vet Microbiol 2000;77(1-2):117-28. doi: 10.1016/s0378-1135(00)00268-6. [ Links ]
17. Baker JC. The clinical manifestations of bovine viral diarrhea infection. Vet Clin North Am Food Anim Pract 1995;11(3):425-45. doi: 10.1016/s0749-0720(15)30460-6. [ Links ]
18. Fritzemeier J, Haas L, Liebler E, Moennig V, Greiser-Wilke I. The development of early vs. late onset mucosal disease is a consequence of two different pathogenic mechanisms. Arch Virol 1997;142(7):1335-50. doi: 10.1007/s007050050164. [ Links ]
19. Wilhelmsen CL, Bolin SR, Ridpath JF, Cheville NF, Kluge JP. Lesions and localization of viral antigen in tissues of cattle with experimentally induced or naturally acquired mucosal disease, or with naturally acquired chronic bovine viral diarrhea. Am J Vet Res 1991;52(2):269-75. [ Links ]
20. Kimura M. A simple method for estimating evolutionary rate of base substitutions through comparative studies of nucleotide sequences. J Molec Evol 1980;16:111-120. [ Links ]
21. Tamura K. Estimation of the number of nucleotide substitutions when there are strong transition-transversion and G + C-content biases. Molec Biol Evol 1992;9:678-687. [ Links ]
22. Fray MD, Paton DJ, Alenius S. The effects of bovine viral diarrhoea virus on cattle reproduction in relation to disease control. Anim Reprod Sci 2000;60-61:615-27. doi: 10.1016/s0378-4320(00)00082-8. [ Links ]
23. Kameyama K, Konishi M, Tsutsui T, Yamamoto T. Survey for detecting persistently infected cattle with bovine viral diarrhea in Japan J Vet Med Sci 2016;78(8):1329-31. [ Links ]
24. Gunn HM. Role of fomites and flies in the transmission of bovine viral diarrhoea virus. Vet Rec 1993;132(23):584-5. doi: 10.1136/vr.132.23.584. [ Links ]
25. Brock KV, Redman DR, Vickers ML, Irvine NE. Quantitation of bovine viral diarrhea virus in embryo transfer flush fluids collected from a persistently infected heifer. J Vet Diagn Invest 1991;3(1):99-100. doi: 10.1177/104063879100300127. [ Links ]
26. Lang-Ree JR, Vatn T, Kommisrud E, Loken T. Transmission of bovine viral diarrhoea virus by rectal examination. Vet Rec 1994;135(17):412-3. doi: 10.1136/vr.135.17.412. [ Links ]
27. Gómez-Romero N, Velázquez-Salinas L, Ridpath JF, Verdugo-Rodríguez A, Basurto-Alcántara FJ. Detection and genotyping of bovine viral diarrhea virus found contaminating commercial veterinary vaccines, cell lines, and fetal bovine serum lots originating in Mexico. Arch Virol 2021;166(7):1999-2003. doi: 10.1007/s00705-021-05089-9. [ Links ]
28. Revell SC, Chasey D, Drew TD, Edwards S. Some observations on the semen of bulls persistently infected with bovine virus diarrhoea virus. Vet Rec 1988;123(5):122-5. doi: 10.1136/vr.123.5.122. [ Links ]
29. Grooms DL, Ward LA, Brock KV. Morphologic changes and immunohistochemical detection of viral antigen in ovaries from cattle persistently infected with bovine viral diarrhea virus. Am J Vet Res 1996;57(6):830-3. [ Links ]
30. Meyling A, Jensen AM Transmission of bovine virus diarrhoea virus (BVDV) by artificial insemination (AI) with semen from a persistently-infected bull. Vet Microbiol 1988;17(2):97-105. doi: 10.1016/0378-1135(88)90001-6. [ Links ]
31. Roeder PL, Harkness JW. BVD virus infection: prospects for control. Vet Rec 1986;118(6):143-7. doi: 10.1136/vr.119.6.143. [ Links ]
32. Hamers C, Lecomte C, Kulcsar G , Lambot M, Pastoret PP. Persistently infected cattle stabilise bovine viral diarrhea virus leading to herd specific strains. Vet Microbiol 1998;61(3):177-82. doi: 10.1016/s0378-1135(98)00185-0. [ Links ]
33. Grooms DL, Brock KV, Pate JL, Day ML. Changes in ovarian follicles following acute infection with bovine viral diarrhea virus. Theriogenology. 1998;49(3):595-605. doi: 10.1016/s0093-691x(98)00010-7. [ Links ]
34. Ssentongo YK, Johnson RH, Smith JR. Association of bovine viral diarrhoea-mucosal disease virus with ovaritis in cattle. Aust Vet J 1980;56(6):272-3. doi: 10.1111/j.1751-0813.1980.tb05722.x. [ Links ]
35. Windsor P. Abnormalities of development and pregnancy. Noakes ED, et al, editors. England, Vet Rep Obst (Tenth ed), W.B. Saunders, 2019; ISBN 9780702072338, https://doi.org/10.1016/B978-0-7020-7233-8.00009-4. [ Links ]
36. Gómez-Romero N, Ridpath JF, Basurto-Alcántara FJ, Verdugo-Rodríguez A. Bovine viral diarrhea virus in cattle from Mexico: Current Status. Front Vet Sci 2021;8:673577. doi: 10.3389/fvets.2021.673577. [ Links ]
37. Fulton RW, Whitley EM, Johnson BJ, Ridpath JF, Kapil S, Burge LJ, Cook BJ, Confer AJ. Prevalence of bovine viral diarrhea virus (BVDV) in persistently infected cattle and BVDV subtypes in affected cattle in beef herds in south central United States. Can J Vet Res 2009;73(4):283-91. [ Links ]
38. Gómez-Romero N, Basurto-Alcántara FJ, Verdugo-Rodríguez A, Bauermann FV, Ridpath JF, Genetic diversity of bovine viral diarrhea virus in cattle from Mexico. J Vet Diagn Invest 2017;29(3):362-365. doi: 10.1177/1040638717690187. [ Links ]
Recibido: 18 de Abril de 2022; Aprobado: 18 de Julio de 2022
 This is an open-access article distributed under the terms of the Creative Commons Attribution License
This is an open-access article distributed under the terms of the Creative Commons Attribution License
