











 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroducción
La salmonelosis transmitida por alimentos es una preocupación de salud pública a escala global1. La carne de diferentes especies, entre ellas la de bovino, funciona como reservorio de su principal agente causal: Salmonella enterica subsp. enterica, de aquí en adelante referida como Salmonella2. Particularmente, la carne de res molida se ha vinculado con brotes recientes de salmonelosis en América del Norte3, por lo cual se considera uno de los principales vehículos de exposición humana a Salmonella. En México se han reportado porcentajes de muestras positivas de entre 16 y 68 % en carne molida en puntos de venta4,5, por lo cual la investigación en esta área resulta relevante desde el punto de vista de salud pública.
Evidencia experimental reciente documenta el aislamiento de Salmonella a partir de nódulos linfáticos de bovinos aparentemente sanos, en frecuencias variables de entre <10 y hasta >90 %6,7. Asimismo, se ha demostrado que los linfonodos superficiales presentan mayores tasas de contaminación que los profundos; mientras que el número de animales con linfonodos contaminados es mucho mayor en ganado de engorda comercial que en los de desecho7,8. No obstante, los resultados suelen variar ostensiblemente según la zona geográfica y la época del año, un fenómeno determinado por mecanismos aun sin descifrar.
En trabajos con cepas de Salmonella obtenidas de animales de desecho, la tipificación de aislamientos, por electroforesis en gel de campos pulsados, demostró clonalidad entre cepas de linfonodos y las presentes en carne molida producida a partir de estos animales9. Sin embargo, este tipo de estudios no se ha realizado en animales de engorda comercial.
A pesar de las altas tasas de positividad a Salmonella reportadas en muestras bovinas en México4-6, la contribución de los linfonodos a este fenómeno prácticamente no ha sido abordada. Por tanto, el objetivo del presente estudio es estimar la frecuencia de contaminación y la diversidad de serotipos de Salmonella presentes en nódulos linfáticos, así como en carne y grasa asociada con estos últimos, en diferentes épocas del año.
Material y métodos
Diseño del estudio y determinación del tamaño de muestra
El tamaño de muestra se calculó mediante la fórmula estadística para estimar una proporción de una población, cuando no se conoce el número de elementos en esa población10:
Donde: n= tamaño de muestra; Zα 2= Valor de Z en una distribución normal Zα= 1.96 cuando α= 0.05; p= proporción de la población con la característica estudiada (si se desconoce se emplea 0.5, como en este caso); q= proporción de la población sin la característica estudiada(1-p); d= error o precisión deseada, fijado en 10% (0.1).
Bajo estos supuestos, se obtuvo un tamaño de muestra de 96, el cual se redondeó a 100. El muestreo se replicó dos veces por año, durante dos años consecutivos, con dos etapas en cada año. Las muestras tomadas entre abril y julio se designaron como muestras de estación “cálida” y las recolectadas entre septiembre y diciembre como muestras de estación “fría”.
Las canales provenían de bovinos machos enteros, cruzas de Bos indicus, con una edad promedio de 24 a 36 meses, procesadas en un rastro Tipo Inspección Federal del estado de Veracruz y trasportadas en refrigeración (<4 ºC), durante aproximadamente 8 h, hasta un punto de venta de la Ciudad de México. Una vez allí, las canales se mantuvieron en refrigeración durante dos días hasta la recolección de las muestras (72 a 96 h post mortem). El punto de venta se visitó los lunes y martes de cada semana, hasta completar entre 5 y 10 canales por semana, según la cantidad de canales disponibles.
Toma de muestras
De cada canal se extrajeron linfonodos superficiales (LNS, cervical superficial y subilíaco) y profundos (LNP, axilar propio y celíaco). La selección de linfonodos se hizo tomando en cuenta la probabilidad de que fueran incluidos en el proceso de molienda, por su ubicación anatómica. Además de los linfonodos, se tomaron aproximadamente 200 g de carne magra (CM, 50 % del diezmillo y 50 % del sirloin, por ser los cortes más usados para producir carne molida) y de carne con grasa (CG), de las zonas circundantes de los LNS y LNP (aproximadamente 50 % de cada una). Previo a los análisis, las porciones individuales de cada tipo de muestra se combinaron para formar una sola muestra compuesta. En algunas ocasiones, ciertas partes de la canal estaban comprometidas para la venta y no estuvieron disponibles para el muestreo. Por ello, no fue posible obtener todos los tipos de muestra del 100 % de las canales. Por consiguiente, la unidad de muestreo se definió como la muestra compuesta de LNS, LNP, CM y CG. En total, se recolectaron 1,545 muestras de todas las fuentes en los dos años de duración del estudio (Cuadro 1).
Cuadro 1 Distribución de las 1,545 muestras de carne y de linfonodos analizadas por estación y año entre el 3 de abril de 2017 y el 14 de diciembre de 2018
| Tipo de muestra | Estación cálida | Estación fría | |||||
|---|---|---|---|---|---|---|---|
| 2017 | 2018 | Total | 2017 | 2018 | Total | ||
| LNS | 168 | 98 | 266 | 33 | 102 | 135 | |
| LNP | 166 | 98 | 264 | 33 | 102 | 135 | |
| CM | 130 | 98 | 228 | 33 | 102 | 135 | |
| CG | 149 | 98 | 247 | 33 | 102 | 135 | |
| Total | 1,005 | 540 | |||||
Estación cálida: abril-julio, estación fría: septiembre-diciembre.
LNS= linfonodos superficiales, LNP= linfonodos profundos, CM= carne magra, CG= carne con grasa.
Las porciones individuales de cada tipo de muestra se colocaron en bolsas plásticas estériles, previamente identificadas y se mantuvieron en hieleras con geles refrigerantes (aproximadamente a 4º C), durante su traslado al laboratorio (duración de dos horas como máximo).
Análisis microbiológicos
La preparación de las muestras de linfonodos se realizó según métodos descritos previamente11, con algunas modificaciones. Los linfonodos se pesaron y, posteriormente, se sumergieron en agua hirviendo durante 5 seg, con el fin de esterilizar su superficie. A continuación, se les agregó la mitad del agua peptonada tamponada (APT) necesaria para alcanzar una dilución aproximada de 1:10 (8 g de LNP en 80 ml de APT y 25 g de LNS en 225 ml de APT) y se molieron durante 3 seg en una licuadora Oster previamente esterilizada. Las muestras molidas se vaciaron en una bolsa Stomacher®, previamente identificada, y con el resto del APT se arrastró el remanente contenido en el vaso de la licuadora, asegurando la transferencia de toda la muestra a la bolsa Stomacher®, realizando posteriormente la homogenización de la mezcla durante 1 min.
Para el análisis de las muestras de carne magra (CM), se tomaron 25 g y se molieron en una licuadora Oster estéril durante 30 seg. Posteriormente, el contenido se colocó en una bolsa de Stomacher® previamente identificada y se agregaron 225 ml de APT, homogenizando la mezcla durante 1 min. Por último, las muestras de carne con grasa (CG) se prepararon moliendo de manera conjunta en una licuadora Oster estéril, durante 30 seg, aproximadamente 1/3 de grasa y 2/3 de carne de las zonas circundantes de LNS y LNP (50 % proveniente de cada tipo de linfonodo). Después de la molienda se pesaron asépticamente 25 g y se siguió el mismo procedimiento descrito para la carne magra.
Los homogenatos se dejaron reposar durante 2 h a temperatura ambiente, antes de seguir los procedimientos de pre-enriquecimiento, enriquecimiento selectivo, aislamiento y confirmación bioquímica de Salmonella spp., establecidos en la norma oficial mexicana vigente12. También se realizó la confirmación molecular de los aislamientos presuntamente positivos a Salmonella spp., mediante PCR, utilizando el gen invA (284 pb), según métodos descritos previamente13. El ADN se extrajo con el kit “Ez-10 Spin Column Bacterial Genomic DNA Miniprep Kit” (BioBasic, Inc., Canadá), siguiendo las instrucciones del proveedor, a partir de las cepas puras, previamente refrescadas en caldo tripticasa soya (MCD Lab®, PRONADISA-CONDA®, España) por 18-24 h. Se utilizaron los cebadores sentido (CGCCATGGTATGGATTTGTC) y contrasentido (GTGGTAAAGCTCATCAAGCG), en reacciones de PCR con un volumen total de 10 μl, empleando los reactivos del MyTaqTM Mix (Bioline, U. K.) con las siguientes concentraciones finales: 5 μl de MyTaqTM Mix, 0.2 μl de cada dNTP y 2.1 μl de agua libre de nucleasas. Las condiciones de termociclado fueron: 94 ºC/3 min de desnaturalización inicial; posteriormente, 35 ciclos de desnaturalización, alineación y extensión (95ºC/45 seg, 62ºC/30 seg, 72ºC/45 seg, respectivamente) y 72ºC/2 min de extensión final. Los productos de amplificación por PCR se procesaron por electroforesis en gel de agarosa (SeaKem® LE Agarose, Lonza, EEUU) al 2%. Los geles se corrieron en buffer tris/borato/EDTA (TBE 1x), a 80 V por 50 min, utilizando SYBR Safe DNA Gel Stain (Invitrogen, USA) para revelar los fragmentos de ADN. La visualización y digitalización de imágenes se realizó en un fotodocumentador Gel Logic 2200 (Kodak, USA) con el software Care Stream® (Carestream Health, Inc., USA). En cada corrida se incluyó una cepa de S. enterica subsp. enterica ser. Typhimurium, del propio laboratorio, previamente confirmada por métodos bioquímicos, PCR y secuenciación completa del genoma. Los aislamientos confirmados se conservaron de dos modos. En el primero, se elaboraron inóculos de 1 ml tomando colonias frescas y mezclándolas en caldo infusión-cerebro-corazón (Merck, Germany) con 10% de glicerol y se mantuvieron a - 70 °C en un ultracongelador. Por otra parte, se realizó un respaldo de los aislamientos en agar soya tripticaseina (TSA, PRONADISA-CONDA®, España), almacenado a temperatura ambiente.
Predicción de serotipos y tipificación por multi-locus (MLST)
El serotipo de las cepas obtenidas se predijo a partir de los datos de secuenciación completa del genoma (lecturas crudas). El ADN genómico se extrajo a partir de colonias refrescadas en caldo TSA con agitación a 37 ºC durante aproximadamente 18 h. Transcurrido este tiempo, se tomó 1 ml de caldo TSA y se centrifugó a 5,000 xg durante 10 min para obtener un pellet de células. Posteriormente, se utilizó el kit de extracción “High Pure PCR Template Preparation Kit” (Roche Molecular Systems, Inc., Switzerland), según instrucciones del fabricante, para obtener el ADN genómico.
La secuenciación se realizó en un equipo Illumina NextSeq (Illumina, USA), utilizando el kit Nextera XT versión 3 (Illumina, USA) para preparar la biblioteca de ADN, con un inserto de 150 pb y una profundidad estimada mínima de 30X. Las lecturas crudas así obtenidas se utilizaron para predecir el serotipo, mediante análisis in silico, con ayuda del programa SeqSero14. Por último, se realizó la tipificación por multilocus (MLST), basada en siete genes esenciales (aroC, dnaN, hemD, hisD, purE, sucA)15, en el servidor del Centro de Epidemiología Genómica16. Como la tipificación por MLST se ha empleado por décadas y existe una base de datos de acceso público17, es posible estimar la importancia epidemiológica de los aislamientos, mediante la comparación con las ST reportadas previamente en muestras clínicas de humanos y de animales. Además, el perfil de alelos se utilizó para generar un árbol filogenético de mínima expansión, con ayuda del programa GrapeTree18, para analizar la diversidad de ST en la muestra bajo estudio.
Análisis estadístico
Se utilizó la prueba de Ji-cuadrada para determinar si existía asociación entre el tipo de muestra, la época del año y el serotipo de Salmonella con la frecuencia de contaminación. En caso de observar asociación significativa, se utilizó la razón de probabilidades para estimar los factores con mayor influencia en la tasa de contaminación de las diferentes matrices estudiadas. Los datos se analizaron con el programa Statgraphics Centurion XV (StatPoint, Technologies, USA).
Resultados
En general, se observó una frecuencia de contaminación con
Salmonella spp. del 5.0 %, con 78 aislamientos obtenidos a
partir de las 1,545 muestras analizadas en los dos años (Figura 1). Se evidenció una fuerte asociación entre el tipo de
muestra y la positividad al patógeno (
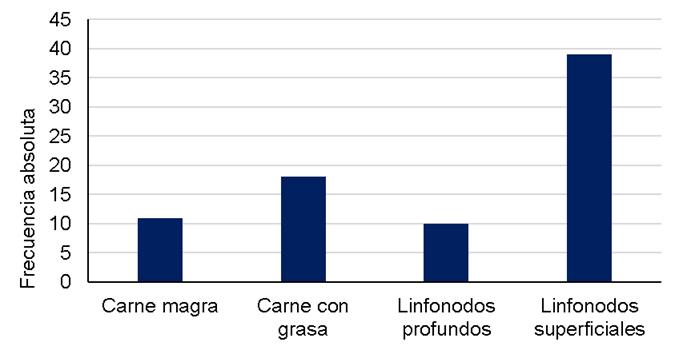
Figura 1 Frecuencia de contaminación con Salmonella spp. en muestras de carne magra (n=363), carne con grasa (n=382), linfonodos profundos (n=399) y linfonodos superficiales (n=401) de bovino, recolectadas entre abril de 2017 y diciembre de 2018
También se observó una asociación significativa entre la frecuencia de contaminación
y la época del año (
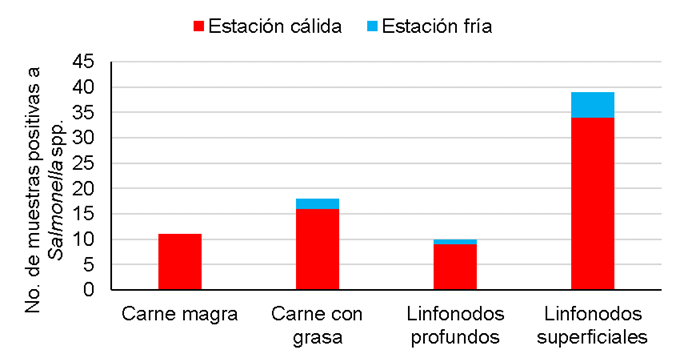
Figura 2 Frecuencia de contaminación con Salmonella spp. en muestras de carne magra (n=363), carne con grasa (n=382), linfonodos profundos (n=399) y linfonodos superficiales (n=401) de bovino, recolectadas entre abril de 2017 y diciembre de 2018
El serotipo de Salmonella también estuvo asociado con el tipo de
muestra (

Figura 3 Número de aislamientos de S. enterica subsp. enterica por serotipo y fuente de aislamiento en las estaciones cálida (a) y fría (b)
La tipificación por MLST mostró que los aislamientos de cada serotipo correspondieron a una misma ST (Figura 4). La excepción fue Salmonella typhimurium, la cual presentó dos ST (19 y 34). Sin embargo, ambos ST sólo se diferenciaron en el alelo dnaN. Por tanto, satisfacen el criterio para ser considerados como complejo clonal, al coincidir en seis de los siete alelos incluidos en el esquema de tipificación por MLST15.

Figura 4 Árbol de mínima expansión generado a partir del perfil de MLST de 78 aislamientos de S. enterica subsp. enterica.
Cada círculo corresponde a un ST y las divisiones dentro de un mismo círculo corresponden a un aislamiento. Los números en las ramas del árbol indican la cantidad de alelos con secuencias distintas entre ST. Los serotipos se indican mediante código de colores y la fuente de aislamiento se señala al interior o adyacente a cada círculo (en texto rojo, si provienen de estación cálida; o en azul, si son de estación fría).
Discusión
Las frecuencias de positividad al patógeno aquí observadas (2.5 a 9.7 %), son inferiores a las reportadas en otros trabajos con carne molida (16-68 %)4,5 y con linfonodos (50-94 %)6,19,20. No obstante, la variabilidad de este fenómeno entre zonas geográficas y épocas del año está bien documentada20,21. En general, el estudio confirma la importancia de los bovinos aparentemente sanos como reservorio de diversos serotipos de Salmonella de importancia epidemiológica. Así lo demuestra la detección de cepas del serotipo Typhimurium, ST 19 y 34, las cuales se asocian con casos clínicos humanos y con la cepa DT104, ampliamente distribuida a escala global22. De igual forma, los aislamientos del serotipo Kentucky (ST 198) se han asociado con infecciones en humanos y animales en Estados Unidos23. Estos hallazgos resaltan la necesidad de continuar investigando las poblaciones de Salmonella de origen no clínico, asociadas con producción animal, por su papel como reservorio de infecciones humanas.
Los resultados apoyan también observaciones previas sobre las mayores tasas de positividad al patógeno en linfonodos superficiales, especialmente en condiciones de clima cálido7,8. Aunque no se han descifrado los factores ambientales causantes de esta variabilidad, la mayor incidencia de picaduras de moscas y de otros insectos en el verano se ha sugerido como un factor condicionante de esta variación estacional19,24. No obstante, la escasa evidencia experimental relacionada con este factor no proviene de contextos naturales sino de estudios de desafío con moscas artificialmente infectadas con Salmonella.
Los esfuerzos realizados hasta el momento para prevenir las infecciones asintomáticas con Salmonella en el ganado han sido infructuosos. El uso de vacunas basadas en genes involucrados en la captación de hierro, mineral con un papel central en el proceso infeccioso, no tuvo efecto alguno sobre la frecuencia de contaminación en linfonodos de bovinos de engorda25. Tal hallazgo es poco sorprendente, considerando la redundancia funcional de Salmonella, la cual posee multiplicidad de genes en materia de captación y transporte de hierro (ej. iroBCDE, fepBCDEG, fhuBCD, exbBD, sitD y tonB)26.
Por otra parte, la supervivencia intracelular de la bacteria, internalizada en vacuolas de células eucariotas27, como los macrófagos, sugiere que la suplementación con antibióticos es una estrategia poco factible. En este sentido, la administración de concentraciones crecientes de tilosina en la dieta de ganado Holstein, previamente inoculado con el patógeno, no mostró efecto alguno, pues se continuó recuperando Salmonella a partir de los linfonodos de los animales tratados28.
Aparentemente, la redundancia funcional de Salmonella y sus mecanismos de supervivencia intracelular indican que la eventual eliminación del patógeno dependerá en última instancia del sistema inmune del hospedero. En animales inoculados experimentalmente con cepas del serotipo Montevideo, la eliminación total de la bacteria tomó cerca de un mes29. En este contexto, el monitoreo de infecciones subclínicas por Salmonella en granjas de engorda, una medida escasamente aplicada, podría funcionar como método para segregar animales portadores y, de esta forma, limitar la diseminación del patógeno. Adicionalmente, la presencia de cepas de un mismo serotipo y ST en carne molida y en linfonodos, observada en este estudio, sugiere a la remoción de los linfonodos como una estrategia para reducir drásticamente la frecuencia de contaminación con Salmonella en carne de res molida. Esta medida es relativamente fácil de realizar a escala industrial, aunque sólo para los linfonodos superficiales, no para los profundos. No obstante, son precisamente los linfonodos superficiales los de mayor relevancia epidemiológica. Por tanto, el establecimiento de esta medida como obligatoria en la normatividad nacional podría funcionar como estrategia para mitigar los riesgos asociados con la presencia de Salmonella en carne de res molida.
Por otra parte, resulta interesante analizar por qué algunos serotipos sólo se presentaron en muestras de carne (ej. Typhimurium), mientras que otros se detectaron en todas las matrices (ej. Anatum y Reading). Particularmente, el serotipo Anatum se ha reportado previamente como cepa predominante en muestras de origen no clínico, sobre todo en linfonodos19,20. Estas evidencias llevan a razonar sobre la posibilidad de que algunas cepas de Salmonella estén mejor adaptadas para colonizar y sobrevivir en ciertos nichos ecológicos. En el contexto del presente trabajo, sin embargo, es difícil determinar si la representación relativa de serotipos en los linfonodos depende de factores genéticos específicos. Además, se requiere emplear análisis de mayor poder discriminatorio que MLST, para explorar con mayor precisión las relaciones filogéneticas intra e inter-serotipo, así como la dinámica evolutiva de estas poblaciones. Este tema será materia de futuras contribuciones en el área de genómica comparativa.
Conclusiones e implicaciones
El estudio demuestra que los linfonodos y la carne de res molida, provenientes de animales aprobados para la matanza, funcionan como reservorios de cepas de Salmonella enterica de importancia clínica en humanos. Por tanto, es preciso establecer medidas de control para prevenir la diseminación de este patógeno a lo largo de la cadena productiva.

