











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
La rentabilidad de los sistemas de producción de carne bovina depende fundamentalmente de la eficiencia reproductiva de la hembra. Se reconoce que las tasas reproductivas altas influyen en forma significativa sobre la eficiencia biológica y económica de los sistemas de producción vaca-cría1. El impacto económico por mejorar el comportamiento reproductivo puede ser hasta cuatro veces mayor que el logrado por mejorar características de crecimiento y calidad de la canal2. Los valores de heredabilidad estimados para características reproductivas son bajos, e indican que los efectos ambientales son los principales responsables de la expresión de la fertilidad en vacas y vaquillas3.
Son pocos los programas nacionales que incluyen evaluaciones genéticas de características reproductivas alrededor del mundo (Estados Unidos, Brasil, Reino Unido y Francia) y pocas las características que se evalúan dentro de cada programa; las principales son preñez de vaquillas, edad al primer parto y permanencia productiva3,4. Sólo en algunos de estos países las diferencias esperadas en la progenie (DEP), como estimadores del mérito genético de la fertilidad de vaquillas, están disponibles para algunas razas, y se han asociado positivamente con la fertilidad en la fase adulta4,5.
Resultados recientes muestran la posibilidad de lograr progreso genético en características reproductivas a través de selección. Heredabilidad de 0.49 ± 0.03 obtenida para tasa de gestación de vaquillas en ganado Nelore, indica que variables como ésta se podrían utilizar como criterio de selección en programas nacionales para mejorar la fertilidad del hato6. Asimismo, Berry y Evans7 encontraron en una población de animales cruzados Bos taurus que la variación genética en características reproductivas era suficiente para asegurar un programa exitoso de mejora genética del comportamiento reproductivo.
En México, se han realizado evaluaciones genéticas nacionales para la Asociación Mexicana Simmental-Simbrah, A.C. y la Charolais Charbray Herd Book de México, A.C., desde el 2001 y 2002, respectivamente. Estas evaluaciones se han hecho para características de crecimiento, circunferencia escrotal, talla y permanencia productiva, utilizando procedimientos multirraciales8, pero no se ha realizado para fertilidad de vaquillas. El objetivo del presente estudio fue desarrollar un prototipo de evaluación genética multirracial para la fertilidad de vaquillas de las poblaciones Simmental-Simbrah y Charolais-Charbray.
MATERIAL Y MÉTODOS
Población de estudio
Se utilizó la información de 49,943 hembras registradas en la Asociación Mexicana Simmental-Simbrah A.C., nacidas entre 1984 y 2011 (EvSim), y la de 49,871 hembras registradas en la Charolais Charbray Herd Book de México A.C., nacidas entre 1990 y 2011 (EvChar). Los registros de las asociaciones indicaron que no se realizó cambio de propietario de las hembras incluidas en las presentes evaluaciones genéticas, por lo que se consideró que permanecieron en el mismo grupo contemporáneo desde el nacimiento hasta los 1,281 días de edad (42 meses). Se definieron cuatro épocas en función de la distribución de los nacimientos de las vaquillas: enero-marzo, abril-junio, julio-septiembre y octubre-diciembre. El grupo contemporáneo (GC) se integró por hato, año y época de nacimiento de la vaquilla. Los GC con menos de cuatro registros no fueron considerados en los análisis. La edad de la madre de la vaquilla al parto se calculó en días. Los Cuadros 1 y 2 resumen la estructura de las bases de datos utilizadas.
Cuadro 1 Estructura general de los datos y de los pedigríes utilizados para evaluar la fertilidad de vaquillas en poblaciones Simmental-Simbrah y Charolais-Charbray
| Simmental-Simbrah | Charolais-Charbray | |
|---|---|---|
| Animales en el pedigrí | 75,926 | 72,486 |
| Animales con registros productivos | 49,943 | 49,871 |
| Sementales en el pedigrí | 4,976 | 5,741 |
| Sementales con progenie con registros productivos | 4,335 | 3,592 |
| Madres en el pedigrí | 37,756 | 41,467 |
| Madres con registros | 33,890 | 30,979 |
Cuadro 2 Medias, mínimos y máximos de la información utilizada para evaluar la fertilidad de vaquillas en poblaciones Simmental-Simbrah y Charolais-Charbray
| Media | Min | Max | |
|---|---|---|---|
| Simmental-Simbrah: | |||
| Crías por semental | 11.52 | 1 | 315 |
| Sementales por año y hato | 3.52 | 1 | 59 |
| Años activos por semental | 2.51 | 1 | 19 |
| Vacas por semental | 10.87 | 1 | 256 |
| Crías por vaca | 1.47 | 1 | 9 |
| Charolais-Charbray: | |||
| Crías por semental | 13.88 | 1 | 346 |
| Sementales por año y hato | 5.19 | 1 | 31 |
| Años activos por semental | 2.61 | 1 | 16 |
| Vacas por semental | 13.03 | 1 | 336 |
| Crías por vaca | 1.61 | 1 | 8 |
Variable de respuesta
La característica evaluada fue la fertilidad de vaquillas (FERT); para cada vaquilla se generó una variable en función de su desempeño reproductivo; a las que parieron antes de los 1,281 días de edad se les asignó un uno y en caso contrario un cero.
Modelos estadísticos
Para determinar si los efectos fijos incluidos en el modelo lineal mixto definitivo explicaban variación significativa, se ajustaron los modelos lineales preliminares9,10 con el procedimiento GLM del paquete estadístico SAS11. De esta manera se decidió incluir la edad de la madre al parto como covariable lineal y cuadrática. El modelo lineal mixto definitivo, empleado en ambas evaluaciones genéticas, quedó estructurado como se muestra a continuación:

Donde, y es el vector de observaciones de FERT; b es el vector de efectos fijos para GC (4,938 y 4,528 niveles para EvSim y EvChar, respectivamente), edad de la madre de la vaquilla al parto, en días, como covariable lineal y cuadrática (EM y EM2, respectivamente), porcentaje de genes Simmental o Charolais (PG), heterocigosis (HET) y pérdidas por recombinación (PR) como covariables relacionadas con el genotipo de la vaquilla12,13; a es un vector aleatorio de efectos genéticos aditivos directos (ANIM); e es un vector aleatorio de efectos residuales; X y Z son matrices de incidencia que asocian los registros de FERT con los correspondientes efectos. Se asumió que los efectos aleatorios siguieron una distribución normal multivariada, con medias iguales a cero y con la estructura de (co)varianzas siguiente:
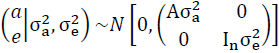
Donde, A es la matriz de parentescos entre los animales que conformaron el
pedigrí (de tamaño 75,926 x 75,926 para EvSim, y 74,486 x 74,486 para EvChar),
Análisis estadísticos
Los estimadores de los componentes de varianza, los valores del logaritmo de la verosimilitud (log L) y las DEP se obtuvieron ajustando un modelo lineal mixto para una sola característica. En la estimación se asumió que FERT se distribuyó binomialmente. Los análisis se realizaron con el paquete estadístico ASREML, utilizando el algoritmo de información promedio de máxima verosimilitud restringida16. Se asumió convergencia cuando el logaritmo de la verosimilitud cambió menos de 0.002 y el estimador del parámetro de la varianza menos del 1 %.
La fórmula utilizada para el cálculo de la heredabilidad (h2) fue:

La DEP es el mejor estimador del mérito genético de un individuo, como esperanza del verdadero valor genético y debe tener un valor de confiabilidad asociado, que tiene una relación directa con la cantidad de información disponible para cada individuo. La confiabilidad se calculó con la siguiente fórmula17:

Con la finalidad de evaluar el cambio de FERT a través del tiempo en las poblaciones estudiadas, se obtuvieron los promedios de las DEP por año de nacimiento de las vaquillas entre 1984 y 2011. Las tendencias genéticas se estimaron mediante la regresión lineal simple de estas medias en los años correspondientes, y la significancia de los coeficientes de regresión obtenidos se probó con el estadístico t 18, usando el procedimiento REG de SAS11.
Se compararon los promedios de las DEP y de las confiabilidades, de la totalidad de sementales y de subconjuntos con el 20, 10, 5 y 1 % superior de los sementales.
RESULTADOS Y DISCUSIÓN
Componentes de varianza y heredabilidad
Los estimadores de los componentes de varianza y heredabilidad para la fertilidad de vaquillas se presentan en el Cuadro 3. Los estimadores de heredabilidad para ambas evaluaciones fueron similares, 0.07 ± 0.008 (EvSim) y 0.06 ± 0.008 (EvChar). Estos valores están dentro del intervalo de estimadores reportados por otros investigadores que utilizaron modelos lineales. En una población multirracial, se estimó que la heredabilidad de la tasa de parto fue 0.06 ± 0.0215). En un programa de selección a largo plazo para características de crecimiento, con líneas Hereford, también se obtuvo una heredabilidad de 0.06 ± 0.06 para tasa de preñez19. Resultados similares se obtuvieron para Hereford y cruzas de Shorthorn y Hereford con Cebú (0.079 y 0.081, respectivamente); sin embargo, heredabilidades para tasa de parto muy próximas a cero se calcularon para Angus, 0.01520. En un estudio realizado con Angus, la heredabilidad del porcentaje de preñez de vaquillas de dos años fue de 0.08 ± 0.06, similar a la encontrada en este trabajo21. En contraste, en la raza Simmental, se estimaron heredabilidades para fertilidad de vaquillas menores a las del presente trabajo, 0.02 ± 0.003 cuando se usó monta natural y 0.016 ± 0.0025 cuando se usó inseminación artificial5.
Cuadro 3 Estimadores de componentes de varianza y heredabilidad de la fertilidad de vaquillas
| Simmental-Simbrah | Charolais-Charbray | |
| Varianza aditiva | 0.0103 (0.0012) | 0.0079 (0.0011) |
| Varianza del error | 0.1326 (0.0013) | 0.1348 (0.0013) |
| Varianza fenotípica | 0.1429 (0.0009) | 0.1428 (0.0009) |
| Heredabilidad | 0.0721 (0.0082) | 0.0594 (0.0077) |
| log L | 6,223.74 | 6,644.90 |
Errores estándar entre paréntesis.
Un estimador ligeramente mayor (0.11 ± 0.03) que el obtenido en el presente estudio, se encontró en una población Nelore en clima tropical, sujeta a selección para peso al año; los autores concluyeron que el avance genético de la fertilidad no se vio comprometido por la mejora en la característica de crecimiento23.
En contraste, Silva et al22 encontraron heredabilidades para preñez de 0.12 en vaquillas que parieron antes de los 46 meses y de 0.52 para las que parieron antes de los 31 meses. Diferencias en heredabilidad de la fertilidad se esperan entre poblaciones Bos taurus y Nelore, dado que estas últimas han sido sometidas a menor intensidad de selección para mejorar la fertilidad24. Por otro lado, estimadores de heredabilidad obtenidos con modelos umbrales son mayores, de 0.21 a 0.5324-28.
Los estimadores de heredabilidad del presente trabajo indican que la expresión de FERT dependió en gran proporción de factores ambientales. Sin embargo, aunque las heredabilidades estimadas en este estudio son bajas, el intervalo observado de las DEP (Cuadro 4) muestra que puede haber respuesta genética de FERT a la selección. Amplitudes ligeramente mayores entre las DEP de sementales se observaron en una población multirracial (-11.3 a 12.2)15 y en una población Angus (−11.94 a 10.01)26; los autores opinaron que estas variaciones son suficientes para seleccionar con el objetivo de aumentar la tasa reproductiva.
Cuadro 4 Diferencias esperadas en la progenie (DEP) y confiabilidades estimadas para grupos de sementales superiores para las evaluaciones Simmental-Simbrah y Charolais-Charbray
| n | DEP (%) | DE | Min | Max | Confiabilidad | |
|---|---|---|---|---|---|---|
| Sementales Simmental | ||||||
| 1% superior | 11 | 6.13 | 1.22 | 5.08 | 8.72 | 0.14 |
| 5% superior | 53 | 3.76 | 1.48 | 2.2 | 8.72 | 0.10 |
| 10% superior | 105 | 2.81 | 1.43 | 1.6 | 8.72 | 0.08 |
| 20% superior | 209 | 2.01 | 1.29 | 0.89 | 8.72 | 0.06 |
| 100% | 1,052 | -0.18 | 1.62 | -5.79 | 8.72 | 0.05 |
| Sementales Simbrah | ||||||
| 1% superior | 9 | 6.94 | 1.05 | 5.64 | 8.84 | 0.18 |
| 5% superior | 46 | 4.41 | 1.48 | 2.88 | 8.84 | 0.13 |
| 10% superior | 92 | 3.34 | 1.51 | 1.84 | 8.84 | 0.10 |
| 20% superior | 185 | 2.37 | 1.44 | 1.08 | 8.84 | 0.09 |
| 100% | 924 | -0.06 | 1.78 | -9.56 | 8.84 | 0.06 |
| Sementales Charolais | ||||||
| 1% superior | 27 | 5.07 | 0.86 | 4.27 | 8.22 | 0.13 |
| 5% superior | 136 | 3.38 | 1.04 | 2.26 | 8.22 | 0.10 |
| 10% superior | 272 | 2.59 | 1.09 | 1.41 | 8.22 | 0.09 |
| 20% superior | 545 | 1.80 | 1.11 | 0.72 | 8.22 | 0.07 |
| 100% | 2,717 | -0.07 | 1.39 | -7.94 | 8.22 | 0.05 |
| Sementales Charbray | ||||||
| 1% superior | 2 | 5.59 | 0.76 | 5.06 | 6.13 | 0.17 |
| 5% superior | 5 | 4.51 | 1.07 | 3.59 | 6.13 | 0.13 |
| 10% superior | 11 | 3.51 | 1.20 | 2.10 | 6.13 | 0.10 |
| 20% superior | 21 | 2.59 | 1.32 | 1.17 | 6.13 | 0.09 |
| 100% | 104 | 0.04 | 1.84 | -7.29 | 6.14 | 0.07 |
n= número de sementales; DE= desviación estándar; MIN = valor mínimo de las DEP; MAX = valor máximo de las DEP.
Grupos contemporáneos
Los efectos no genéticos han sido incluidos en los modelos de predicción para remover sesgos en las evaluaciones genéticas. El grupo contemporáneo se incluye para minimizar la variación que ocasionan los cambios en las condiciones ambientales del hato, que se dan por las asociaciones entre los grupos contemporáneos y el nivel genético de los animales29. La varianza del error de predicción es afectada por el tamaño de los grupos contemporáneos, así como por el tamaño de las familias utilizadas en la evaluación. La disminución del tamaño de los grupos contemporáneos aumenta el número de grupos contemporáneos que deben ser estimados y los grados de libertad perdidos en el proceso. En la Figura 1 se presenta la distribución del número de hembras en grupos contemporáneos de EvSim y EvChar, respectivamente. Las medianas del número de hembras por grupo contemporáneo de EvSim y EvChar fueron similares, con 10, en un rango intercuartil de 5 a 20. En el presente trabajo se encontró, en ambas poblaciones, que la confiabilidad de los valores genéticos disminuyó a medida que decreció el tamaño del grupo contemporáneo. Los valores de confiabilidad más bajos de los sementales estuvieron por debajo de 0.12 y 0.08 para EvSim y EvChar, respectivamente, cuando el tamaño del grupo contemporáneo fue menor de 10. Ugarte et al29 encontraron que la cantidad efectiva de información para la estimación de valores genéticos disminuyó con el tamaño del grupo contemporáneo, lo que aumentó el error de predicción, aun cuando el número de hijas por semental fue constante y los mayores valores de estos fueron de grupos contemporáneos menores de 9.
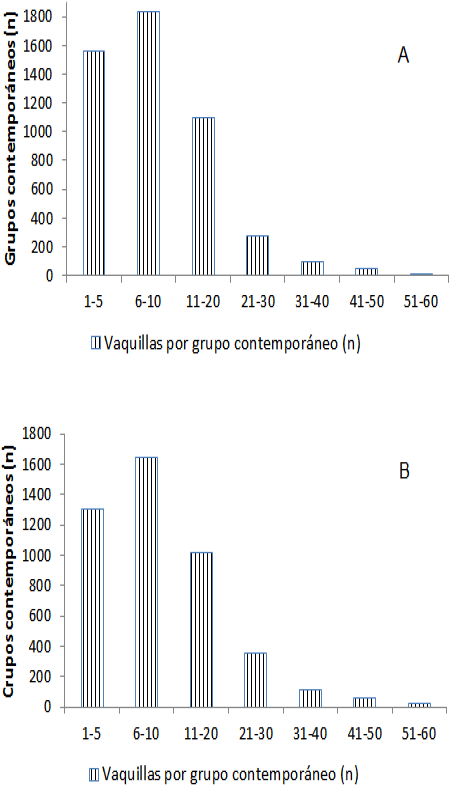
Grupos familiares
El número de sementales utilizados en el presente estudio fue 4,335 y 3,592 para EvSim y EvChar, respectivamente. La distribución del tamaño de las familias paternas de las dos evaluaciones se presenta en la Figura 2. La relación entre el tamaño de los grupos contemporáneos y el de las familias de medias hermanas utilizadas en la evaluación genética determina los niveles de confiabilidad y el sesgo de los valores genéticos obtenidos. El tamaño óptimo de las familias de medios hermanos para la estimación de la heredabilidad está determinado por la relación 4/h230,31. Bajos estimadores de heredabilidad se obtuvieron en ambas evaluaciones, 0.07 y 0.06 para EvSim y EvChar, respectivamente. La relación entre el número de hijas y la confiabilidad de las DEP de los sementales se presenta en la Figura 3. En ésta se puede observar que para alcanzar niveles de confiabilidad superiores a 0.3 se requiere que un semental tenga más de 100 hijas. En un estudio de reproducción sostenida exitosa, se encontró media y mediana de tamaño de familia de 7 y 11, respectivamente; también estimaron que para alcanzar una confiabilidad de 0.5 un semental necesitaría 90 hijas, considerando la heredabilidad de 0.05; por lo que los investigadores determinaron que para mejorar la confiabilidad se requiere que el tamaño de las familias aumente sustancialmente31. Como alternativa para evitar el descarte de información proveniente de GC pequeños, en especial de hijas de toros jóvenes, investigadores proponen la utilización combinada GC fijos y aleatorios selectivos, en donde han encontrado que su utilización interfiere poco en el ordenamiento de los valores genéticos, la estimación de los parámetros genéticos y las tendencias genéticas32.
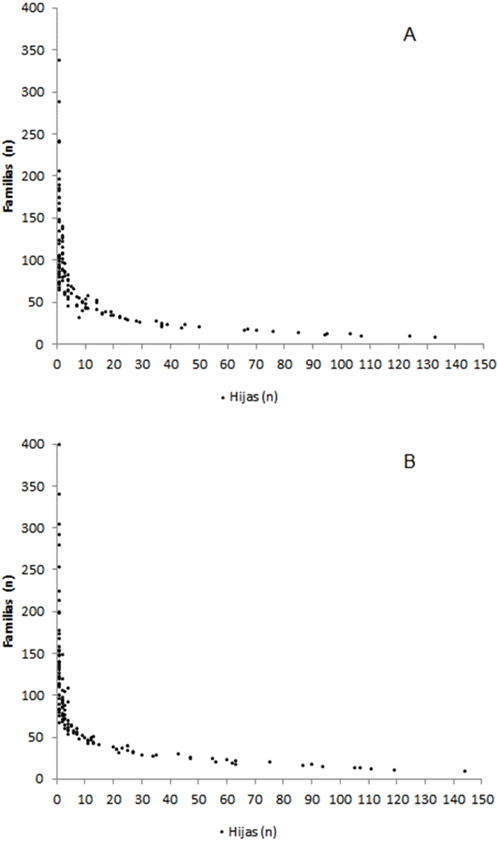
Figura 2 Distribución del tamaño de las familias paternas de las vaquillas en la evaluación Simmental-Simbrah (A) y Charolais-Charbray (B)

Tendencias genéticas
La tendencia genética anual en la fertilidad de vaquillas es el cambio en el mérito genético aditivo en la población para esta característica en el tiempo. En la Figura 4 se presentan las tendencias de las DEP para FERT, a través de los años, de las razas Simmental, Simbrah, Charolais y Charbray. Tanto para EvSim como para EvChar las DEP fueron ajustadas por el promedio de las DEP del año 2001; este año se consideró como la base genética en ambas poblaciones. El cambio genético anual para Charolais, Simmental y Simbrah, entre los años de 1984 y 2011, se estimó en 0.012 ± 0.001 (P<0.001), 0.008 ± 0.004 (P<0.05) y 0.002 ± 0.003 puntos porcentuales (P>0.05), y las medias generales ajustadas de las DEP para este período fueron 0.063 ± 0.069, 0.028 ± 0.166 y 0.09 ± 0.134, respectivamente. En 1984, la DEP promedio de FERT fue -0.063, -0.36 y -0.16, para las poblaciones Charolais, Simmental y Simbrah, respectivamente. A partir de este año, el promedio de las DEP incrementó de manera uniforme hasta llegar, en el 2011 a 0.29 en Charolais, 0.25 en Simmental y 0.19 en Simbrah. También se observa la tendencia genética de la población Charbray con un patrón errático, que coincide con los años de menor cantidad de información disponible de esta raza. El cambio genético anual observado en Charbray entre 1988 y 2011 fue el mayor de entre las cuatro razas (0.1334 ± 0.006 (P<0.05); el promedio de las DEP fue 0.041 ± 0.219. A partir del 2001, la tendencia de FERT para la población Charbray siguió una pauta similar a las de las demás razas evaluadas. Es importante resaltar que las tendencias genéticas estimadas en ambas poblaciones muestran que existió una respuesta genética favorable de la fertilidad de las vaquillas de las cuatro razas evaluadas, durante el período 2007-2011.
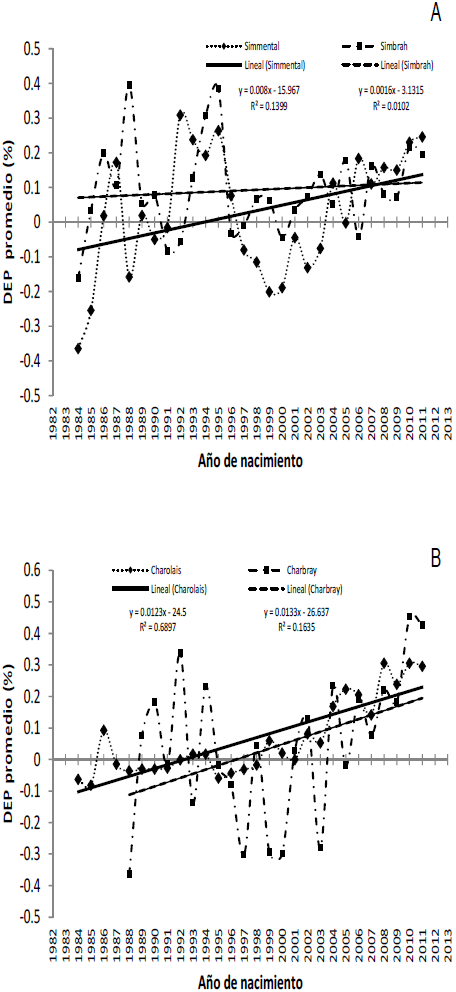
Figura 4 Tendencia genética de la fertilidad de vaquillas para la evaluación Simmental-Simbrah (A) y Charolais-Charbray (B)
En general, se observó un ligero cambio genético en las poblaciones en estudio, aun cuando se ha aplicado mayor presión de selección sobre las características de crecimiento, como el peso al destete, donde las DEP se han incrementado en el tiempo33. Resultados similares se encontraron en un estudio de selección para peso al año en Nelore, donde las medias anuales de los valores genéticos para tasa de parto a primer servicio se mantuvieron prácticamente sin cambio durante 15 años23; estos autores concluyeron que las ganancias significativas en peso corporal del año en adelante, como resultado de su proceso de selección, no interfirieron en el comportamiento reproductivo de las vacas, en específico de los días al parto y la tasa de parto. Al comparar los resultados obtenidos en el presente trabajo con los de un estudio de tendencias genéticas para tasa de no retorno de vaquillas Noruegas, se encontró que el cambio genético fue positivo (0.16 %) y solo fue similar a la tendencia observada en Charbray34.
Ordenamiento de sementales
En el Cuadro 4 se presentan los estadísticos de los grupos de sementales ordenados en rangos por su mayor nivel de DEP para EvSim y EvChar. El intervalo de fluctuación de las DEP fue similar para Simmental, Simbrah y Charolais, donde el valor máximo de las DEP estuvo alrededor de los 8 puntos porcentuales; en cambio el intervalo en Charbray fue menor. En la Figura 3 se observa que las confiabilidades más altas estuvieron por arriba de 0.40, en los grupos familiares con más de 100 individuos. Aun cuando la heredabilidad estimada de FERT para las poblaciones estudiadas fue baja, habría que tomar en cuenta la amplitud existente entre las DEP de los sementales. Para ponerlo en contexto, consideremos que un semental que tiene una DEP de 8 % tiene la posibilidad de producir hijas con un 7 % más de probabilidad de parir a los 42 meses de edad que las hijas de otro semental con una DEP de 1 %.
En un estudio con Angus35, las DEP para porcentaje de preñez fluctuaron entre -0.02 y 0.01, intervalo que fue menor al observado en el presente trabajo. Estos autores concluyeron que, aunque la heredabilidad fue baja (0.12), con la variabilidad de los valores de cría obtenidos se podría lograr progreso genético mediante selección. Por otro lado, intervalos de DEP más amplios (de -11.3 a 12.2 %) se estimaron en una población multirracial Bos taurus, para porcentaje de parto15. Estos autores también determinaron que el intervalo de las DEP no fue afectado por el tipo de modelo estadístico que se utilizó, lineal, umbral o logístico. Con vaquillas Nelore y un modelo umbral, otros investigadores encontraron que el intervalo de las DEP y los valores de éstas fueron mayores que los que se obtuvieron en el presente trabajo (-24.5 a 24.55); atribuyendo estos resultados tanto al método de estimación utilizado, como a la mayor variabilidad genética de la fertilidad de las vaquillas en la población estudiada36.
CONCLUSIONES E IMPLICACIONES
Las heredabilidades estimadas indican que puede haber respuesta genética a la selección de FERT en las poblaciones evaluadas. Las tendencias genéticas estimadas muestran un cambio genético favorable de la fertilidad de vaquillas Simmental, Simbrah, Charolais y Charbray durante el período 2007-2011. Los rangos de los valores de las DEP permiten identificar sementales sobresalientes para la fertilidad de sus hijas en las cuatro razas estudiadas

