











 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroducción
En regiones cálidas como en el noroeste de México, la combinación de humedad y temperatura de verano fácilmente exceden los requerimientos térmicos del ganado Holstein1,2. Al salir de la zona termoneutral, la vaca activa un complejo sistema de ajustes endócrinos y metabólicos regulados por genes para mejorar las funciones celulares afectadas por el estrés calórico3, que finalmente comprometen la productividad del ganado lechero debido a que dichos ajustes implican un gasto energético asociado al incremento del ritmo respiratorio4.
La temperatura corporal del bovino oscila entre los 37.5 y los 38.9 ºC en animales sanos bajo condiciones termoneutrales, pero en climas cálidos su incremento es el resultado de la carga de calor autogenerado y el proveniente del medio ambiente5. Esta hipertermia se ha correlacionado negativamente con el crecimiento, lactancia y fertilidad en ganado Holstein6,7. El tracto respiratorio es una ruta evaporativa de eliminación de calor y el animal incrementa su frecuencia respiratoria como un mecanismo regulador de la temperatura corporal en climas cálidos8,9.
Además de estar implicadas con el metabolismo energético, crecimiento, fertilidad, lactogénesis y desarrollo mamario, la prolactina e IGF13,10,11, son hormonas relacionadas en el proceso de termorregulación en los rumiantes y el incremento de sus niveles en sangre son una respuesta sensible al ambiente cálido12,13,14.
El enfriamiento artificial es una alternativa para aminorar los efectos negativos de las altas temperaturas sobre la fisiología y productividad de la vaca lechera2,15, sin embargo, bajo condiciones de moderada a alta humedad la eficiencia de un sistema de enfriamiento de alta presión (nebulizadores) es limitado8. Por esta razón, mojar el pelaje y piel del animal combinado con ventilación forzada, favorece las pérdidas de calor por las vías conductivas y convectivas16,17,18.
Conforme aumenta la intensidad de la humedad y temperatura ambiental al inicio de verano en el valle del Yaqui, Sonora, la eficiencia productiva del ganado lactante disminuye debido a un clima gradualmente más adverso para esta zona lechera19. Incrementar la frecuencia de baños durante el día puede aminorar algunos indicadores fisiológicos de estrés calórico y evitar el descenso abrupto en la producción del ganado Holstein conforme empeoran las condiciones climáticas de verano. Por lo tanto, el objetivo del estudio fue evaluar la respuesta fisiológica, endócrina y productiva del ganado Holstein al someterlo a varias series de enfriamiento directo al inicio de verano.
Material y metodos
Localización experimental y animales en estudio
El estudio se realizó en la Unidad académica de investigación en producción lechera del Instituto Tecnológico de Sonora, ubicada en el Block 910 (N: 27°21’ O: 109°54’) del Valle del Yaqui, Sonora, México, durante junio y julio de 2011. Se seleccionaron 28 vacas Holstein de entre 3 y 5 partos, con 150 ± 25.9 días en leche y condición corporal de 3.5 ± 0.35, las cuales se alimentaron dos veces al día (0800 a 0900 h y 1700 a 1800 h). Todas las vacas recibieron la misma ración durante la fase experimental (aproximadamente 48.8 % de silo de maíz, 16.2 % de heno de alfalfa y 35 % de concentrado energético comercial, adicionado con vitaminas y minerales) y con libre acceso al agua. Como manejo cotidiano del establo, a partir de la semana nueve postparto y hasta 15 días antes del secado, las vacas recibían (vía subcutánea) un programa comercial de somatotropina bovina (500 mg de STb) a intervalos de 14 días durante la lactancia.
Tratamientos
Aleatoriamente se asignaron 14 animales a cada tratamiento: enfriamiento (E) antes de la ordeña (dos veces, a las 0700 y 1700 h) y enfriamiento adicional (EA) cada dos horas (seis veces por día, de 0700 a 1700 h). De esta forma, los animales de los tratamientos E y EA fueron llevados diariamente a la sala de espera a la ordeña donde recibieron dos y seis periodos de enfriamiento, equivalentes a 36 y 108 min/día de enfriamiento acumulado por tratamiento, respectivamente. En ambos grupos, cada periodo de enfriamiento era una secuencia de tres ciclos de baño alternado con tres de ventilación.
Equipo de enfriamiento
Por ciclo de enfriamiento se aplicaban 3 min de aspersión de agua (gasto de 15 L por vaca/ ciclo) con 16 regaderas (instaladas a 2.8 m del piso) hasta mojar por completo el lomo y costado del animal, alternados con 3 min de ventilación forzada (11,000 CFU) con tres abanicos eléctricos de ½ HP cada uno situados a 2.73 m del piso. Todas las vacas tuvieron 8.5 m2 de sombra en los corrales de alojamiento.
Datos climáticos
La información obtenida de temperatura ambiental (TA) y humedad relativa (HR) se colectó por una estación meteorológica automática ubicada a ~500 m del sitio experimental. El Índice de temperatura-humedad (ITH) se calculó para estimar el nivel de estrés calórico al que estuvieron expuestos los animales utilizando la fórmula7: ITH= (0.8*TA)+[(HR/ 100)*(TA”14.4)]+46.4. Para obtener valores representativos del clima durante el periodo de muestreo, la información climática se registró por hora de 0900 a 1700 h y promediando la información respecto a los dos últimos días previos a la fecha de muestreo4.
Indicadores fisiológicos de estrés calórico
En ambos grupos durante el estudio, la frecuencia respiratoria (FR; resp/min) y la temperatura rectal (TR; °C) se registraron a las 1200 h dos veces por semana. Para medir la FR se contaron visualmente los movimientos costales1, mientras que la TR se registró utilizando un termómetro digital (TES-1310®) con sonda (tipo K) de contacto (largo de 9 cm) que era introducida para hacer contacto con la mucosa del recto del animal20.
Dinámica de niveles hormonales en sangre
Entre las 1000 y 1100 h los animales de ambos grupos se sangraron una vez por semana mediante punción de la vena de la cola utilizando equipo vacutainer® para extraer 8 a 10 ml de sangre en tubos sin anticoagulante. En cada muestreo, los tubos se identificaron y transportaron (5 °C) al Laboratorio de Biotecnología Reproductiva del Departamento de Ciencias Agronómicas y Veterinarias del ITSON (Cd. Obregón, Sonora, México) para centrifugarse a 2,500 rpm por 30 min y extraer el suero. Las muestras séricas se transportaron al Laboratorio de Endocrinología de New Mexico State University, USA, para medir las concentraciones (ng/ml) de prolactina (PRL) y del factor de crecimiento parecido a la insulina tipo 1 (IGF1), por radioinmunoanálisis (Packard Instrument Company Inc., Meriden, CT, USA) de doble anticuerpo11,12 bajo un coeficiente de variación de 9.4 % y 13.4 % para PRL e IGF1, respectivamente.
Producción de leche
La producción (kg) individual de leche (PL) correspondiente a las dos ordeñas del día (0700 y 1700 h), se registró diariamente utilizando un sistema electrónico de ordeño (Metraton 21™, GEA. Westfalia-Surge Farm Technologies, Siemensstraße, Bönen Germany).
Análisis estadístico
Los procedimientos estadísticos PROC UNIVARIATE y PROC GLM (prueba de Levene) se utilizaron para analizar la distribución normal e igualdad de varianzas, respectivamente, de todas las variables analizadas. Mediante el procedimiento MIXED del paquete estadístico SAS V9.421, los comandos Repeated y Random se utilizaron para analizar las mediciones repetidas por semana de FR, TR, PRL, IGF1 y PL. El modelo mixto incluyó los efectos fijos de tratamiento, tiempo, la interacción tratamiento por tiempo, y el número de lactancias, así como el efecto aleatorio de la vaca. La variable días en leche se consideró como covariable dentro del modelo, la cual resultó ser una fuente significativa de variación (P<0.05). De acuerdo a los criterios Bayesiano y Akaike, se seleccionó la estructura de covarianza auto-regresiva para las mediciones repetidas del modelo mixto, ya que ésta resultó ser la estructura de covarianza de mejor ajuste. Si el efecto tiempo resultó significativo (P<0.05), se ejecutaron comparaciones pareadas de medias de cuadrados mínimos utilizando la opción PDIFF en SAS.
Resultados y discusión
Datos climáticos
Durante el estudio, la temperatura ambiental mostró una variación de 30.4 hasta 36.6 °C (Cuadro 1). El umbral térmico máximo para la raza Holstein es de 25 °C 7, y con fluctuaciones térmicas entre 21 y 32 °C se han observado pérdidas en leche22. El nivel de HR durante el día se mantuvo entre 18.1 y 32.8 % a lo largo del experimento. El efecto de la humedad es insignificante a 24 °C, pero cuando la temperatura del día es >34 °C su impacto negativo es considerable23, y a partir de una HR >45 % las funciones evaporativas (jadeo y sudoración) de regulación térmica son limitadas24. Aunque la HR no alcanzó ese límite, sí fue evidente una tendencia en su intensificación al inicio del verano, lo que representaría la entrada a las condiciones regulares de estrés por calor en esta región. El promedio de ITH durante el día fue de 79.1 unidades, considerándose un estrés calórico moderado15, experimentándose condiciones más adversas después de la semana cuatro. Collier et al25 demostraron que a partir de 68 unidades de ITH, la producción en vacas altas productoras comienza a declinar.

Indicadores de estrés calórico
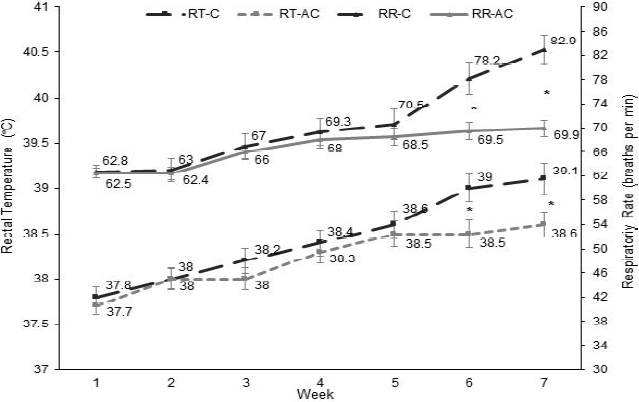
El Cuadro 2 muestra que la interacción tratamiento por tiempo fue significativa (P<0.001) para FR y TR, considerados los indicadores de estrés calórico. En cierto modo era de esperar que al incrementar el número de baños el beneficio pudiera ser mayor; sin embargo, el efecto del tratamiento sobre las variables indicativas de estrés calórico sólo se observó al final del estudio. Ello coincidió con el inicio del verano (semanas 6 y 7), por lo que el efecto favorable de incrementar la frecuencia de baños dependió de la variable tiempo/ semana, y por lo tanto, de las variables climáticas. Berman20 probó diferentes niveles de enfriamiento a 38 °C y 47 % de humedad, donde fueron necesarios hasta siete ciclos de baño y ventilación para disminuir la temperatura rectal de vacas Holstein, sin observar mejoría al aumentar los ciclos a ocho o diez. Spiers et al9 reportaron bajo condiciones termoneutrales (TA de 19 °C y HR de ~55 %), un grupo de vacas mostraron FR de 59.6 respiraciones/ minuto y TR de 39 °C, pero al ser sometidas a un periodo de estrés calórico (TA de 29 °C y HR de ~50 %), su FR y TR se incrementaron en 32.8 % y 2.5 %, respectivamente. En este estudio la FR fue similar entre tratamientos (P>0.05) en las primeras cinco semanas (Figura 1), pero fue menor en el grupo EA en comparación con el grupo E (P<0.05) a partir de la semana seis (78.5 vs 82.9 respiraciones/ minuto) y siete (69.5 vs 69.9 respiraciones/ minuto), lo que coincidió con el momento en que las condiciones climáticas fueron más severas. Del mismo modo, la TR en EA fue menor que en E (P<0.05) sólo en las semanas 6 (38.5 vs 39.0 °C) y 7 (38.6 vs 39.1 °C). A lo largo del experimento, las vacas en el grupo E incrementaron (P<0.001) en 32.0 y 3.4 % su FR y TR, respectivamente, mientras que en EA las vacas sólo mostraron un aumento de 11.8 y 2.3 % en su FR y TR, respectivamente. Es importante mencionar que, en general, la diferencia no es tan amplia debido a que ambos grupos recibieron enfriamiento. Cuando la vaca rebasa los 39.2 °C de TR y las 70 respiraciones/ minuto de FR, entra a una condición de estrés por calor, por lo que bajo estas condiciones el enfriamiento artificial tiene como objetivo mantener estas variables lo más cercano a un umbral fisiológico. Al concluir este estudio, ambos grupos lograron mantener la TR <39.2 °C aunque en el E la FR fue >70 respiraciones/ minuto.
Cuadro 2 Medias de cuadrados mínimos y probabilidad para los efectos de tratamiento, tiempo y su interacción en vacas Holstein expuestas a diferentes frecuencias de enfriamiento

RR= Respiratory rate (breaths per minute); RT= Rectal temperature (°C); PRL= Prolactin levels (ng/ml); IGF1= Insulin like growth factor-1(ng/ml); MY= Milk yield (kilograms per day).
Dinámica de niveles hormonales en sangre
Los niveles de PRL difirieron por tratamiento (P= 0.004) y por efecto del tiempo de muestreo (P<0.001; Cuadro 2). Durante el experimento, los valores promedio de PRL fueron más elevados para el tratamiento EA (35.1 ± 1.3) en comparación con E (29.4 ± 1.1 ng/ml). En contraste, Igono et al22 reportaron una reducción en los niveles de PRL en vacas que recibieron enfriamiento más sombra, en comparación con aquéllas que sólo recibieron sombra, por lo cual concluyeron que la PRL sérica se redujo al disminuir el estrés calórico y mejorar el confort ambiental de la vaca. Sin embargo, en la presente investigación es probable que los niveles superiores de PRL encontrados en las vacas con enfriamiento adicional estén asociados al proceso de lactación, ya que el muestreo sanguíneo se realizó durante las mañanas inmediatamente después del ordeño. Por lo tanto, el horario de toma de la muestra pudo haber enmascarado el efecto del estrés calórico sobre los niveles de PRL en vacas sin enfriamiento adicional.
Respecto al tiempo de muestreo, la Figura 2 muestra una reducción gradual de PRL (P<0.05) en las semanas 4 y 5, estabilizándose sus niveles en las semanas 6 y 7. Contrario a estos resultados, Scharf et al14 han reportado que la concentración de PRL incrementa en climas cálidos, ya que es una hormona homeotérmica ante eventos de estrés por calor26,27. Del mismo modo, la PRL estimula la expresión de las proteínas de choque de calor (HSP)28, las cuales evitan la desnaturalización de proteínas en células ante la amenaza de estrés calórico en el bovino 29 . Es probable que la respuesta no esperada en los niveles de PRL a mediados del mes de julio se debió a la corta duración del experimento, ya que el impacto del clima de esta zona es más severo para el ganado después de julio19 , por la acumulación de la carga calórica en la vaca, siendo posible que la activación de mecanismos que involucran la PRL sería más evidente a finales del periodo de verano. Además, la covariable días en leche resultó significativa (P<0.05), por lo que dicha variable pudo haber influido en la disminución en los niveles de PRL al final del estudio.
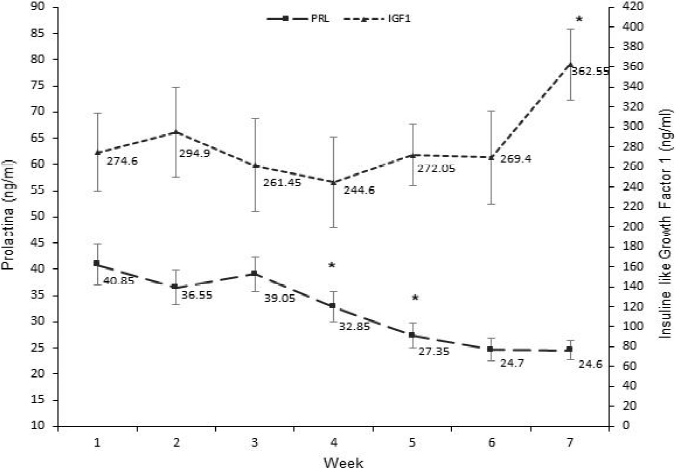
As effect of time [*] indicate statistical difference (P<0.05) in relation to the previous week on average level per week (i.e., including both treatments; ng/ml) of PRL and IGF1 during the study.
Figura 2 Dinámica de los niveles (ng/ml) de prolactina (PRL) y del factor de crecimiento similar a la insulina tipo-1 (IGF1) durante el estudio
La concentración de IGF1 solo se afectó por efecto de tiempo (P=0.0143; Cuadro 2), observándose un incremento (P<0.05) al final del ensayo (Figura 2). Rhoads et al10 han reportado una ligera reducción en los niveles de IGF1 del ganado expuesto a condiciones cálidas. La hormona del crecimiento (GH) y el IGF1 tienen una relación muy estrecha en procesos metabólicos que promueven la producción de leche11 , por lo que la medición de GH apoyaría en la explicación del efecto ambiental sobre la IGF1 y su relación con la producción de leche. Vale la pena señalar que las vacas de este estudio recibían la aplicación de un producto comercial como manejo habitual a base de somatotropina bovina (STB). La STB es una hormona sintética cuya función es similar a la GH en el bovino y su administración exógena incrementa potencialmente la carga de calor en ganado lactante bajo estrés calórico13,30,31, así como también la concentración de IGF1 en sangre32,33,34,35. Por lo tanto, la administración de STB en este trabajo pudo inducir un efecto directo sobre los niveles de IGF1 y consecuentemente en los de PRL. Aún y cuando los niveles de IGF1 se mantuvieron constantes pero sin una tendencia clara a lo largo de casi todo el experimento, durante la semana 7 sí se observó un incremento significativo de IGF1 (P<0.05) alcanzando su máxima concentración, lo que coincide con el periodo más caliente del muestreo.
Existen rutas alternativas que puede tomar la GH para movilizar energía de otros tejidos en el bovino ante la simultánea demanda energética de las funciones de mantenimiento (ej. termorregulación) y lactogénesis en el ganado lechero especializado10. La regulación metabólica de PRL e IGF1 permite priorizar algunas funciones emergentes como la del intento de adaptarse a un clima cálido en transición y mantener una lactancia (por demanda ambiental) en ganado especializado, aún en bajos niveles. La PRL tiene acción en la síntesis de leche en rumiantes, sin embargo, no es esencial para mantener la lactancia36, mientras que la ruta del IGF1 estimula la galactopoyesis e interactúa en forma independiente de la prolactina y STB37. Lo anterior, puede explicar la caída en los niveles de PRL y el incremento del IGF1 en el periodo más cálido de este estudio.
Producción de leche
La interacción de tratamiento por tiempo fue significativa (P=0.002) para PL (Cuadro 2), observándose mayor producción en EA en comparación con E en las semanas 6 (17.9 vs 21.0 kg) y 7 (18.0 vs 21.1 kg), cuando el nivel de estrés (representado por ITH) fue mayor (Figura 3). El enfriamiento directo (baños) y ventilación forzada en ambientes cálidos y húmedos, ha demostrado que puede reducir los efectos negativos de la temporada de calor sobre la fisiología de la vaca16,18, contribuyendo en el estado hormonal12 y corrigiendo el metabolismo energético13, que en conjunto mejoran su desempeño productivo1. Se ha reportado una mejoría de 9.3 % en la producción de leche al incrementar de cinco a ocho las sesiones de enfriamiento en la sala de espera17. Con temperaturas entre 27 y 46 °C, el enfriamiento de baja presión en la sala de espera a la ordeña puede evitar pérdidas en leche de 0.76 kg/d por vaca15.

MY= Milk yield. [*] In means of columns indicate the effect of treatment by week (P<0.05).THI= Temperature-Humidity Index.The treatments C= Cooling and AC= Additional cooling.
Figura 3 Efecto del enfriamiento adicional sobre la producción de leche (PL) en vacas Holstein durante el estudio
En este experimento, la reducción en PL inició en la tercera semana cuando las vacas experimentaron un nivel de ITH de 76.6 unidades (Figura 3). Collier et al25 demostraron que las vacas con una producción >35 kg/día requieren enfriamiento artificial a partir de un ITH de 68 unidades, ya que al alcanzar este umbral, la pérdida negativa en leche se ha estimado en 2.2 kg/día. Trabajos realizados en Sonora por Leyva et al19 identificaron que a partir de un umbral de 71.2 unidades de THI las pérdidas alcanzaban 0.65 kg/día por vaca19.
Mientras que el efecto negativo de verano se acercaba, condiciones de estrés calórico más altas se mitigaron con el aumento de las sesiones de enfriamiento en EA durante el día. Esto representa una ventaja teniendo en cuenta que todos los cambios fisiológicos y metabólicos que la vaca puede experimentar en un intento de regular su temperatura corporal por sí sola, podrían disminuirse con el enfriamiento suplementario. Es notable como los niveles de PRL se mueven en la misma dirección que la PL en ambos grupos (Figura 2 y 3), por lo que la PRL se relaciona con la lactancia en vez de termorregulación. Sin embargo, es probable que el aumento en los niveles de IGF1 y el enfriamiento suplementario permitieran la movilización de energía para mantener la lactancia ante una demanda energética cada vez más alta para mantenimiento (termorregulación), lo cual fue experimentado por las vacas en las últimas dos semanas.
Conclusiones e implicaciones
Incrementar la frecuencia de baños en vacas Holstein mitigó la magnitud de los indicadores fisiológicos de estrés por calor al inicio del verano, lo cual se reflejó en el mantenimiento de la producción de leche en una época del año caracterizada por el incremento gradual en el nivel del ITH. El comportamiento de los niveles de prolactina e IGF1 no fue el esperado debido posiblemente al efecto de la corta duración del experimento y el horario matutino de muestreo hormonal, en el caso de la prolactina, y debido a la aplicación de STB comercial, en el caso de prolactina e IGF1. Un incremento significativo en producción de leche de 3.1 L por vaca se encontró en las semanas 6 y 7, que representa un ingreso adicional de $23.25 por vaca, el cual es probable que no compense el costo por la elevación en el consumo de agua que implica le tecnología de enfriamiento adicional, a menos que la empresa cuente con un sistema de reciclaje de agua que permita mayor eficiencia en este proceso. En base a lo anterior, se sugiere un estudio de impacto económico antes de iniciar con un sistema de enfriamiento adicional. Por otra parte, la identificación de ganado con un umbral de tolerancia al estrés calórico más alto sería una alternativa de impacto económico y ambiental para seguir produciendo leche en verano con poca o nula cantidad de agua para su enfriamiento artificial.

