










Servicios Personalizados
Revista
Articulo
 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de ciencias pecuarias
versión On-line ISSN 2448-6698versión impresa ISSN 2007-1124
Rev. mex. de cienc. pecuarias vol.6 no.4 Mérida oct./dic. 2015
Technical notes
Yield and nutritive value of forage legumes in two seasons and four regrowth stages
a Campus Tabasco. Colegio de Postgraduados (CP). Periférico Carlos A. Molina s/n 86500 H. Cárdenas, Tabasco. México.
b Campo Experimental Huimanguillo, Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP). Km 1 Carretera Huimanguillo - Cárdenas. Huimanguillo, Tabasco. México. Tel. (917) 375-0764,
c Área de Ciencia Animal. CP. Cárdenas, Tabasco. México.
d Área de Ciencia Vegetal. CP. Cárdenas, Tabasco. México.
The effect of age of regrowth (21, 42, 63 and 84 d) on the dry matter yield (DMY), leaf proportion (LP) and nutritive value of cacahuatillo (Arachis pintoi), stylo (Stylosanthes guianensis), clitoria (Clitoria ternatea) and kudzu (Pueraria phaseoloides) was evaluated during the dry season (March-May) and rainy (June-August) of 2013. The DMY and neutral detergent fiber (NDF) increased with age, while the LP, protein (CP) and degradability decreased. Stylo and cacahuatillo recorded the highest DMY in both seasons; on average 5.5 and 7.5 t ha-1 at 84 d in dry and rainy season respectively. In both seasons, the highest and lowest LP were for cacahuatillo and stylo, respectively. High CP was recorded by clitoria and cacahuatillo in both seasons; greater than 14 g kg-1 DM even 84 d old. No significant variation was observed in CP from 42 d in the species. At 21 d, stylo (704.7 g kg-1 DM) in the dry season and kudzu (658.4 g kg-1 DM) in the rainy season had the largest NDF. There were no differences between species from 42 d. Cacahuatillo had the highest degradability (>700 g kg-1 DM) during its growth in both seasons. In clitoria, stylo and kudzu was just above 700 g kg-1 DM at 21 d, but from 42 d of age, degradability decreased. In conclusion, the age of regrowth that integrates DMY and better nutritional value is 42 d old in the legumes evaluated, regardless of the season of year.
Key words: Arachis pintoi; Stylosanthes guianensis; Clitoria ternatea; Pueraria phaseoloides; Protein; Degradability; Neutral detergent fiber
El efecto de edad de rebrote (21, 42, 63 y 84 días) sobre el rendimiento de materia seca (RMS), proporción de hoja (PH) y valor nutritivo de cacahuatillo (Arachis pintoi), stylo (Stylosanthes guianensis), clitoria (Clitoria ternatea) y kudzú (Pueraria phaseoloides), se evaluó durante la época seca (marzo-mayo) y lluviosa (junio-agosto) de 2013. El RMS y la fibra detergente neutro (FDN) se incrementaron con la edad, mientras que la PH, proteína (CP) y degradabilidad disminuyeron. Cacahuatillo y stylo registraron los RMS más altos en las dos épocas; en promedio 5.5 y 7.5 t ha-1 a los 84 días en época seca y lluviosa, respectivamente. En ambas épocas, la mayor y menor PH fueron para cacahuatillo y stylo, respectivamente. En CP, clitoria y cacahuatillo presentaron las mayores en ambas épocas; superiores a 140 g kg-1 MS aún a 84 días de edad. No se observó variación significativa en CP a partir de los 42 días en las especies. A los 21 días, stylo (704.7 g kg-1 MS) en la época seca y kudzú (658.4 g kg-1 MS) en la época lluviosa presentaron la mayor FDN. No hubo diferencias entre especies a partir de los 42 días. Cacahuatillo tuvo la mayor degradabilidad (>700 g kg-1 MS) durante su crecimiento en ambas épocas. En clitoria, stylo y kudzú sólo fue superior a 700 g kg-1 MS a los 21 días, pero desde los 42 días de edad, la degradabilidad disminuyó. En conclusión, la edad de rebrote que integra mayor RMS y valor nutritivo es la de 42 días en las leguminosas evaluadas, independientemente de la época del año.
Palabras clave: Arachis pintoi; Stylosanthes guianensis; Clitoria ternatea; Pueraria phaseoloides; Proteína; Degradabilidad; Fibra detergente neutro
Grasslands in the tropical region of Mexico (dry and humid tropics) occupy an area of 12 million hectares, producing 81 million tons of dry matter and with one third of the country’s cattle herd under the grazing system also combined with stabling1 but to a lesser extent. In the humid tropics, animal production (meat, milk, calf) in the state of Tabasco is obtained mainly from permanent grassland grazing. For this reason, the dominance of pasture management and use of pastures with a higher production of dry matter and quality are important for productivity and sustainability of livestock production systems in this region. However, tropical grasses are commonly of low nutritional value, particularly protein. Depending on the season, regrowth age and species, the protein content in pasture varies from 6 to 14 %, and in some cases contain less than 6 %2. This low concentration in protein affects animal productivity, by requiring the rumen to be at least 7 % of this nutrient for optimal functioning3, which together with the seasonal forage production limits the sustainability of animal productivity through the year4.
An alternative to improve the diet of grazing animals and maintain sustainability of the system is introduced forage legumes in the pasture or in association or in banks of protein. Legumes fix nitrogen in the soil, which can benefit the associated grass and provide high quality and economic food. The protein content and minerals in legumes is much higher than grasses5, although it has been observed that the former also have variations in content when compared between humid and dry regions6. There is sufficient information on dry matter yield of tropical forage legumes at the end of a period and at different cutting intervals7,8. However, little information is available about the optimal harvest time considering the greater quantity and quality of forage, since the first increases and second declines with age9. By understanding this behavior under humid tropical conditions in Tabasco, will allow for designing management schemes for tropical forage legumes to ensure their quantity and quality for animal feeding.
Given the above, the objective of this study was to determine the effect of regrowth age of the plant on yield and nutritive value of four forage legumes into two contrasting periods of the year in terms of moisture availability.
This study was conducted under field conditions during the dry (March, April and May) and rainy (June, July and August) seasons in 2013 in the garden of forage genetic resources of the Huimanguillo Experimental Station (17°50’ N, 93°23’ W), of the National Institute of Livestock Agricultural and Forestry Research (INIFAP) in Tabasco. The soil was of a loamy texture with 41.1 % sand, 24.5 % clay and 34.4 % loam, pH 7.0 and 21.5 millequivalents 100 g-1 of ion exchange capacity. No fertilizer was applied, nor was irrigation available in the dry period of the year. Climatological data is presented in Figure 1. The biological material consisted of four species of herbaceous forage legumes: cacahuatillo (Arachis pintoi Krapovickas & Gregory), stylo (Stylosanthes guianensis (Aubl.) Sw.), or Tehuana clitoria (Clitoria ternatea L.) and kudzú (Pueraria phaseoloides Roxburgh Bentham). These species were sown on August 8, 2012 in plots of 2 x 4 m, at a seeding density of 5 kg seed ha-1 distributed in five furrows drawn along the length of each plot and spaced at 50 cm, except cacahuatillo. This latter was seeded with vegetative material (root stems) at a distance of 25 cm from stem to stem. The plots were distributed at random in the field, spacing between them was 1.5 m and with three replicates per species.

Each plot was divided into four subplots of 2 x 1 m, one of four regrowth ages: 21, 42, 63 and 84 d were assigned at random. The regrowth age in days were counted after cutting uniformly, which occurred for all plots on February 26 for the dry season, and May 20 for the rainy season. Thus, harvests in the dry season were carried out on March 18, April 8 and 29 and for the rainy season on May 20, on June 10, July 1 and 22 and August 12. In each subplot (regrowth age), all biomass or green matter (GM) with cuts of 10 cm from ground level for stylo, and 5 cm for the other legumes harvested. The collected material was weighed in a portable electronic scale with capacity of 10 kg ± 1 g. To determine the yield of dry matter (DMY t ha-1), subsamples of 300 g of GM were separated, which were dried in forced air oven at 65 °C for 48 h. The DMY per species within each regrowth age was calculated based on the concentration of DM obtained from 300 g of GM and GM yield total harvest of each subplot. The leaf-total aboveground biomass ratio was determined by taking leaves and stems of an additional subsample of 100 g of GM. These leaves and stems were separated and dried separately in stoves with forced air at 65° C for 48 h. After drying the leaf-total aboveground biomass ratio was obtained by dividing the dry weight of leaves by total dry weight of leaves + stems.
The protein concentrations (g kg-1 DM) and neutral detergent fiber (NDF, g kg-1 DM) were quantified in the dry samples of GM 300 g, which were previously ground to a particle size of 1 mm. Protein concentration was determined by the Kjeldahl method, this involves multiplying the percentage of nitrogen x 6.2510. NDF was determined using sodium lauryl sulfate at a neutral pH by the methodology of Van Soest11.
The in situ DM (g kg-1 DM) degradability was determined at 48 h of incubation according to the technique of nylon bags12 with the recommendations of Villalobos13 and Ayala14, using cross bred bulls Bos Taurus x Bos indicus with ruminal fistula. Four bulls were used and for each bull the sample was introduced in duplicate; i.e. 24 bags per bull. The formula for the calculation of in situ degradability = (g initial DM - g residual DM) / g initial DM.
The variance analysis applied to all the variables of response was based on the SAS GLM program15. The effect of species, age of regrowth and interaction species x age of regrowth was analyzed separately by time of year, using a completely randomized design in split plot arrangement. The main plot was the species and the secondary plot regrowth age. These two factors plus their interactions were included in the model. Comparison of means was carried out by the method of Tukey (P<0.05) and correlations between variables with the CORR program, calculated from the averages of the field data for each particular species x age of regrowth.
Environmental conditions
The temperature and precipitation for the two periods evaluated for the year are presented in Figure 1. In the dry season (March, April and May) the highest temperatures and lowest average rainfall was recorded compared to the rainy season. In March the lowest temperature was recorded, 3.7 °C lower than the average of April and May. These last two months had 1 °C higher average temperature than the rainy season. There was a marked difference in rainfall between the two periods, 190 mm of rain was recorded in the dry season than in the rainy season. In May there were 98 mm more rainfall than the average for the months of March and April.
Total dry matter yield (DMY) and leaf-total aboveground biomass ratio
There was interaction species x regrowth age in both seasons for the DMY (Table 1). In the dry season, stylo was the species that mostly contributed to this interaction by increasing its DMY 3.7 t ha-1, from 63 to 84 d of regrowth (Figure 2a). This significant increase changed the production status stylo with respect to other species, and coincided with the significant increase in rainfall in May. This result shows the high potential of stylo in response to favorable moisture conditions. Kudzu was the species had the lowest yields throughout the dry season, but at 21 and 84 d showed similar yields to that of clitoria, with average values of 0.39 and 3.56 t ha-1, respectively. In the rainy season the interaction species x age of regrowth was caused by four species, which changed its production status by age of regrowth (Figure 2c) and stylo again recorded a high DMY at 84 d regrowth, which coincided with the wettest month recorded (August) within the rainy season. At this time, again clitoria and kudzu are the species with smallest DMY throughout their growth period, with similar values as stylo at 21 d (0.865 t ha-1, the average of the three species), and cacahuatillo at 84 d (5.61 t ha-1 on average). In both seasons, the DMY had a progressive increase with age of regrowth, with an average variation of the four species, from 0.749 to 4.37 t ha-1 during the 21 to 84 d of regrowth in the dry season. In the rainy season this increase was 1.17 to 6.52 t ha-1 in the same time period of growth.
Table 1 Least square means for total dry matter yield (DMY), leaf-total above ground biomass ratio (LTAB), protein, degradability and neutral detergent fiber (NDF) for four legumes harvested at four ages of regrowth in two seasons.

**, *** Significance at a probability level of 0.01 and 0.001, respectively. DF= degrees of freedom.

Figure 2 Yield of dry matter and Leaf total aboveground biomass ratio of four forage legumes into four stages of regrowth during the dry (a, b) and rainy (c, d) season.
The interaction of species x age of regrowth was present only in the rainy season for the leaf-to-stem ratio (LSR) (Table 1). In the dry season, the four species maintained their values in terms of LSR at different ages of regrowth (Figure 2b), where cacahuatillo and stylo species had the highest and lowest values of LSR at the different ages of regrowth, respectively. In the rainy season interaction was induced by clitoria, it changed from being a species with high LSR at 21 d of regrowth, to become a species with lower LSR (along with stylo) at 42 d, to later increase its LSR with advancing age of regrowth (Figure 2d). Cacahuatillo and stylo species also had higher and lower LSR, respectively, at different ages of regrowth at this time (except in rainy season at 21 d). In both seasons, a decreasing pattern in the values of leaf-total aboveground biomass ratio with advancing age of regrowth was recorded, being more accentuated in all species during the rainy season (Table 2). However, between regrowth ages, after 42 d no differences of regrowth was found for this parameter in all four legumes evaluated in both seasons. In the dry season, the average value of leaf-total aboveground biomass ratio species decreased from 0.676 to 0.480 at 21 to 84 d of regrowth ages, and 0.786 to 0.450 during the rainy season in the same period of growth.

Protein
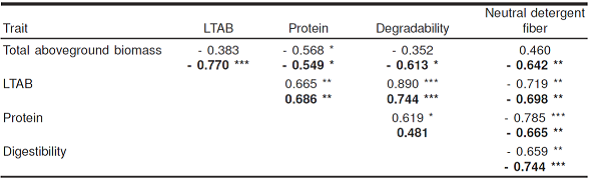
In both seasons there was interaction between species x age of regrowth (Table 1), induced by the four species in the dry season, in terms of protein in each regrowth age (Figure 3a). In the rainy season, the interaction was given by cacahuatillo to become the species with the greatest concentration of protein at 21 d, the species with lowest concentration was stylo and kudzu (124.3 g kg-1 DM average for the three species) at 84 d (Figure 3d). During both seasons the protein concentration, the average of the four species, decreased with age of regrowth and ranged from 186 to 133.8 g kg-1 DM at 21-84 d in the dry season, and 174.64 to 134.44 g kg-1 DM in the rainy season during the same period. However, the protein concentration showed its greatest decline after an age of 42 d, in most species tested. In both seasons of the year clitoria and cacahuatillo maintain the highest concentration of protein with regrowth age, also evident was that cacahuatillo and kudzu did not alter their protein concentration after 42 d of regrowth in the dry season (Table 2). The latter two species also maintained stability in terms of leaf-total aboveground biomass ratio in the dry season, which could influence the close relationship that exists between protein concentration and leaf-total aboveground biomass ratio (Table 3). Stylo, however, was the species that recorded the lowest concentrations of protein (which coincides with the lowest values of leaf-total aboveground biomass ratio) at different ages of regrowth in both seasons (except at 21 d in the dry season). In this legume, protein concentrations were observed in the rainy season greater than levels observed in the dry season from 42 d of age.

Degradability
The interaction species x regrowth age in both seasons was produced by the change of regrowth age in the stylo, clitoria and kudzu species (Figures 3b and c). In both seasons, the degradability of DM maintained a decreasing pattern with age. In the dry season decreasing from 757.48 to 624.99 g kg-1 DM, during the 21 to 84 d of regrowth, respectively, and in the rainy season, from 731.72 to 587.29 g, in the same period. In both seasons, cacahuatillo recorded the highest degradability values at different ages of regrowth (with values above 700 g at different ages), and showed greater stability by decreasing its degradability to just 57 g at 21 to 84 d of regrowth in the dry season, and 59 g in the rainy season over the same regrowth period. The previous result (Table 2) is related to the higher leaf-total aboveground biomass ratio present in cacahuatillo during both seasons, since it has lower fiber concentration, and consequently the reason for its greater degradability, variations in degradability were highly related to variations in the values of leaf-total aboveground biomass ratio (Table 3). The remaining species (stylo, clitoria and kudzu) degradability values were below 650 g kg-1 DM after regrowth age of 42 d in both seasons.
Neutral detergent fiber
There was interaction of species x age of regrowth for NDF given by the change of status in the dry season cacahuatillo, clitoria and kudzu, with advancing age of regrowth, and when the four species grew in the rainy season (Figures 3c and f). In both seasons a slight increase in the NDF with plant age, except stylo in the dry season (Table 2) was recorded. In this period, stylo maintained the highest values in each of the regrowth ages, with an average concentration of 724 g kg-1 DM. The average of the four species showed that NDF increased from 590 to 687.51 g from 21 to 84 d of regrowth in the dry season, and 586.87 to 683.38 g in the rainy season in the same period. Cacahuatillo was the species that recorded the lowest values in all regrowth ages except at 21 d in the dry season. In general, the concentration of NDF was inversely related to the leaf-total aboveground biomass ratio, the protein concentration and the degradability of dry matter (Table 3).
The interaction species x age of regrowth was present for all traits evaluated, which show that tested species have a differential response to the environment for growth16, mainly soil moisture, and possibly photoperiod17, which makes it more difficult to maintain a stable response to different environmental conditions that arise during the growth period. Also the intensity and quality of light and water stress can affect the chemical composition of the plant18,19 and the response can vary between legumes20. This is mainly reflected in the differences in DMY of species between in the seasons evaluated, which has implications for the availability of forage between species per season for livestock. Furthermore, the maturity of the plant influences the availability and quality of feed19. The DMY and NDF increased with age of regrowth in all species and the LSR, protein and digestibility decreased, and more acutely after 42 d. Cacahuatillo and stylo always showed divergent responses in all traits evaluated, possibly a product of their contrasting growth habits: prostrate and erect, respectively.
The highest DMY in species were obtained at 84 d, which is consistent with other studies in northern Veracruz21, being stylo and cacahuatillo the species that excelled. In the dry season, cacahuatillo had yields above 3 t ha-1 after 42 d of regrowth, although the literature presented it as a species with little tolerance to drought22. In fact this response was expected of stylo based on reports of its tolerance to drought23. This result is possibly due to the fact that during the months of the dry period there was light rain reaching values of 50 mm per month, with the month of May being exceptional with about 150 mm, which favored the growth of cacahuatillo since water availability influences cell division, elongation and differentiation of leaf organs20. By contrast, stylo recorded a high potential response to favorable moisture conditions, this species having the largest DMY during the rainy season in the different regrowth ages and surpassed cacachuatillo in the wettest month of the dry season (May), with 1.6 t ha-1. Information is available in the literature8 that supports this high responsiveness of stylo to favorable moisture conditions compared with other legumes.
The variations in the LSR in total aboveground biomass, was a key factor in the variations of nutritive value, particularly protein, since a large proportion of N in the above ground vegetative material is derived from the reserves of N that are mobilized at the base of the stems or roots to the stems and leaves in development24,25. In addition, moisture promotes soil N mineralization and availability26. Therefore, cacahuatillo species with the highest values of leaf-to-stem ratio at different ages of regrowth, it also had the highest concentration of protein degradability and the lowest NDF, in most ages of regrowth of the two seasons studied. These significant concentrations of protein and degradability of cacahuatillo are similar to those found in the same species in studies in South America27. By contrast, stylo is a plant that is erect or semi-erect with a semi-woody main stem, which serves to as support, and has short branches, presented the lowest LSR and protein concentration in both seasons, and the highest concentration of NDF in the last two cutting ages during the rainy season. This lower protein concentration in stylo has been attributed to its high proportion of stems8,23, which increases the concentration of NDF.
Kudzu was the species with the lowest adaptation to the dry season, with DMY below 1 t ha-1up to 42-d old, with a better response in the month of May (84-d old), due to increased storm precipitation. Although there are reports of high adaptation of this species due to drought23, this adaptation is detrimental to their forage production. Although this species showed a leaf ratio close to that of cacahuatillo (above 600 g kg-1 DM up to 42 d, and about 500 g kg-1 DM from 63 d in both times), was overtaken by clitoria in protein concentration in the different ages of regrowth. There are reports22, which point to the low concentration of protein in kudzu, though it has many leaves. However, the degradability of kudzu was intermediate, with the rest of legumes in the dry season, and equal to those recorded for stylo and clitoria, while stylo was higher in the different ages of regrowth in the rainy season.
Clitoria was a species with intermediate response in DMY characters and LSR in both seasons. Although it was not a species with high LSR, recorded the highest concentrations of protein in the dry season, with similar values to that of cacahuatillo at 84 d regrowth, with values just below 200 g kg-1 DM after 42 d of regrowth in both seasons. Since many years ago, protein values were reported up to 314 g kg-1 DM in clitoria23, the higher concentration than in other legumes studied.
Figures 2 and 3 show that between both seasons, the difference was due to DMY (greater in the rainy season), given that the nutritional value of legumes between seasons showed no significant variations. Generally, the DMY of legumes increased and nutritional value decreased with age of regrowth in both seasons, and also after 42 d in both seasons the decreasing trend was even steeper. Therefore, 42 d of regrowth of legumes probably should be considered the maximum allowable age for exploitation for animal intake in both periods.
Literatura citada
1. Bolaños-Aguilar ED, Émile JC, Enríquez-Quiroz JF. Les fourrages au Mexique: ressources, valorisation et perspectives de recherche. Fourrage 2010;204:277-282. [ Links ]
2. Juárez-Hernández J, Bolaños-Aguilar ED, Reinoso M. Contenido de proteína por unidad de materia seca acumulada en pastos tropicales. Época de Nortes. Rev Cub Cienc Agric 2004;38:423-430. [ Links ]
3. Van Soest PJ. Nutritional ecology of the ruminant; LTTIECA and London: Comstock Publishing Associates, Cornell University Press; 1984. [ Links ]
4. Fulkerson WJ, Neal JS, Clark CF, Horadagoda A, Nandra KS, Barchia I. Nutritive value of forage species grown in the warm temperate climate of Australia for dairy cows: Grasses and legumes. Livest Sci 2007;107:253-264. [ Links ]
5. Onyeonagu CC, Eze SM. Proximate compositions of some forage grasses and legumes as influenced by season of harvest. Afric J Agric Res 2013;8:4033-4037. [ Links ]
6. Adjolohoun S, Bindelle J., Adandedjan C, Toleba SS, Houinato M, Sinsin B. Variety and environmental effects on crude protein concentration and mineral composition of Arachis pintoi (Kaprovickas & Gregory) in Benin (West Africa) 2013;1:24-28. [ Links ]
7. Hare MD, Tatsapong P, Phengphet S, Lunpha A. Stylosanthes species in north-east Thailand: dry matter yields and seed production. Trop Grassland 2007;41:253-259. [ Links ]
8. Phengsavanh P, Frankow-Lindberg BE. Effect of harvesting interval on biomass yield and nutritive value of five tropical forage legumes (Aeschynomene histrix ‘BRA 9690’, Canavalia brasiliensis ‘CIAT 17009’, Stylosanthes guianensis ‘CIAT 184’ and ‘Composite’ and Vigna unguiculata ‘CIAT 1088-4’) in Lao PDR. Grassland Sci 2013;59:80-86. [ Links ]
9. Hiep NV, Wiktorsson H, Man NV. The effect of cutting interval on foliage yield and chemical composition of tropical Kudzu (Pueraria phaseoloides) cultivated as cover-crop in rubber plantation. Livest Res Rural Dev 2008;20 (Suppl),: Livest Res Rural Dev 2008;20 (Suppl),: http://www.lrrd.org/lrrd20/supplement/hiep1.htm Accessed July 27, 2014. [ Links ]
10. AOAC. Association of Official Analytical Chemist. Orr. of Anal. Chem. 17th ed. Washington, USA. 2000. [ Links ]
11. Van Soest PJ. Nutritional ecology of the ruminant; LTTIECA and London: Comstock Publishing Associates, Cornell University Press; 1994. [ Links ]
12. Orskov ER. Protein nutrition in ruminants 2nd ed. London, England: Academic Press; 1992. [ Links ]
13. Villalobos GC, González VE, Ortega SJA. Técnicas para estimar la degradación de la proteína y materia orgánica en el rumen y su importancia en rumiantes en pastoreo. Téc Pecu Méx 2000;38:119-123. [ Links ]
14. Ayala BAJ, Rosado RCM, Capetillo LCM, Sandoval CCA. Evaluación del método de lavado de bolsas (manual vs lavadora) en la técnica de degradación ruminal in situ. Téc Pecu Méx 2003;41:337-342. [ Links ]
15. SAS Institute. User‘s Guide: Statistics, version 9.3. SAS Inst. Inc., Cary, North Caroline, USA. 2010. [ Links ]
16. Kretschmer AE. Consideraciones sobre factores que afectan la persistencia de leguminosas forrajeras tropicales. CIAT. Pasturas Tropicales 1988;10:28-33. [ Links ]
17. Pitman WD. Environmental constraints to tropical forage plant adaptation and productivity. In: Sotomayor-Ríos A, Pitman WD editors. Tropical forage plants. Development and use. Boca Raton Florida. USA: CRC Press, 2001:17-23. [ Links ]
18. Perbandt D, Fricke T, Wachendorf M. Effects of changing simulated sky cover on hyperspectral reflectance measurements for dry matter yield and forage quality prediction. Comp Electr Agric 2010;73:230-239. [ Links ]
19. Evers GW. Forage legumes: Forage quality, fixed nitrogen, or both. Crop Sci 2011;51:403-409. [ Links ]
20. Fenning JO, Quansah C, Sarfo-Kantanka A. Response of three forage legumes to soil moisture stress. J Sci Technol 2009;29:24-30. [ Links ]
21. Valles B, Castillo E, Hernández T. Producción estacional de leguminosas en Veracruz, México. CIAT. Pasturas Tropicales 1992;14:32-36. [ Links ]
22. Meléndez NF. Principales forrajes para el trópico. 1rst ed. Tabasco, México: SAGARPA-UPCH; 2012. [ Links ]
23. Bogdan AV. Pastos tropicales y plantas de forraje (Pastos y leguminosas). 1a ed en español. México DF: AGT Editor SA; 1997. [ Links ]
24. Volenec JJ, Ourry AA, Joern BC. A role for nitrogen reserves in forage regrowth and stress tolerance. Physiol Plantarum 1996;97:185-193. [ Links ]
25. Erice G, Sanz-Sáez GA, Aranjuelo I, Irigoyen JJ, Aguirreolea J, Avice JC, Sánchez-Díaz M. Photosynthesis, N2 fixation and taproot reserves during the cutting regrowth cycle of alfalfa under elevated CO2 and temperature. J Plant Physiol 2011;168:2007-2014. [ Links ]
26. Dewhurst RJ, Delaby L, Moloney A, Boland T, Lewis E. Nutritive value of forage legumes used for grazing and silage. Irish J Agric Food Res 2009;48:167-187. [ Links ]
27. Godoy EV, Barrera AA, Vivas MR, Quintana ZJ, Peña GM, Villota GL, et al. Evaluación fenológica y digestibilidad in vivo de la leguminosa forrajera (Arachis pintoi) en diferentes edades de corte. Ciencia y Tecnología 2012;5:7-16. [ Links ]
Received: September 22, 2014; Accepted: November 24, 2014
 Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
