










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista mexicana de ciencias pecuarias
versão On-line ISSN 2448-6698versão impressa ISSN 2007-1124
Rev. mex. de cienc. pecuarias vol.4 no.2 Mérida Abr./Jun. 2013
Notas de investigación
Crecimiento en longitud foliar y dinámica de población de tallos de cinco asociaciones de gramíneas y leguminosa bajo pastoreo
Leaf length growth and tiller population dynamics in five different grasses and legume associations under grazing conditions
Rigoberto Castro Riveraa, Alfonso Hernández Garayb, Omar Ramírez Reynosoc, Gisela Aguilar Benitezb, Javier Francisco Enríquez Quirozd, Sergio Iban Mendoza Pedrozab
a Centro Interdisciplinario de Investigación para el Desarrollo Integral Regional, Instituto Politécnico Nacional, Unidad Oaxaca. Hornos 1003, Sta. Cruz Xoxocotlán, Oaxaca, México. Tel. 01 951 51 706 10 Cxt. 82754. rcastror@ipn.mx, rigocastro4@hotmail.com. Correspondencia al primer autor.
b Colegio de Postgraduados en Ciencias Agrícolas-Campus Montecillo. Texcoco, México.
c Unidad Académica de Medicina Veterinaria y Zootecnia. Universidad Autónoma de Guerrero. Cuajinicuilapa Guerrero. México.
d Campo experimental La Posta. Instituto Racional de Investigaciones Forestales, Agrícolas y Pecuarias. Paso del Toro Veracruz. México.
Recibido el 7 de octubre de 2010.
Aceptado el 3 de mayo de 2011.
Resumen
El objetivo del estudio fue determinar la elongación, crecimiento, senescencia, peso y dinámica de tallos de cinco asociaciones conformadas por dos gramíneas y una leguminosa sembradas en diferentes proporciones. Se evaluaron cinco tratamientos: 4:3:3; 4:6:0; 4:0:6; 4:4:2 y 4:2:4 de trébol blanco-ovillo-ballico perenne, los cuales se distribuyeron en 20 unidades experimentales de 104 m2, en un diseño en bloques completos al azar. La mayor elongación y crecimiento neto de la hoja de las especies evaluadas fue en verano, con 7.1 y 6.53 cm tallo-1 d-1, para el pasto ovillo, y 7 y 6.7 cm tallo-1 d-1, para ballico perenne; en trébol blanco no existió diferencia en el recambio de tejido foliar y del peciolo en verano y primavera (P>0.05). El mayor peso de tallos de ballico perenne y ovillo se registró en verano (0.38 g-1 tallo-1) y las mayores densidades en la época de invierno (9,961 y 10,423 tallos m-2), respectivamente. El recambio de tejido de las especies evaluadas presentó marcada estacionalidad, siendo más dinámico en verano que en otoño. La asociación de tres especies permitió una mayor dinámica en comparación con la asociación de dos especies.
Palabras clave: Elongación, Crecimiento neto, Dinámica de tallos, Praderas asociadas.
Abstract
The purpose of the present study was to determine leaf length, net growth, senescence, tiller weight and tiller dynamics of five associations between two grasses and a legume planted in different proportions. Five treatments were evaluated: 4:3:3; 4:6:0; 4:0:6; 4:4:2 and 4:2:4 of white clover, cocksfoot, and perennial ryegrass, respectively. These treatments were distributed in 20 paddocks, 104 m-2 each, in a randomized complete block design. The greatest increase in both leaf length and leaf net growth in the species tested was found in summer, with 7.1 and 6.53 cm tiller-1 d-1 for cocksfoot and 7 and 6.7 cm tiller-1 d-1 for perennial ryegrass; while in white clover no differences in leaf length and leaf net growth and petiole elongation in summer and spring (P>0.05) were not found. The greatest tiller weight in perennial ryegrass and cocksfoot was recorded in summer (0.38 g-1 stem-1) and highest tiller population density in the winter season (9,961 and 10,423 tiller m2), respectively. Tissue turnover in the assessed species showed a noticeable seasonality, being more dynamic in summer that in autumn. Associations of three species allowed for one major tiller population dynamics compared to those of two-associated species.
Key words: Elongation, Net growth rate, Tillers dynamics, Associated swards.
La asociación de dos o más especies de gramínea-leguminosa tiene como ventaja que la producción de forraje se mantiene constante durante el año. Algunos autores(1) mencionan que las praderas asociadas mejoran la calidad nutricional, la productividad de los animales en pastoreo y la fertilidad del suelo por la fijación de nitrógeno(2,3).
En una pradera asociada las unidades de crecimiento son los tallos (gramínea) y estolones (trébol blanco), de modo que el aumento en la producción de forraje se atribuye a incrementos en la densidad y peso individual de estas unidades o a una combinación de ambos(4). Cl rendimiento está integrado por la tasa de crecimiento individual y por la tasa de aparición de nuevos tallos y estolones(5); por ello es importante conocer la dinámica de población de tal l os y estolones en praderas bajo condiciones de pastoreo.
En praderas asociadas, los tallos y estolones están continuamente emergiendo, creciendo y muriendo, a tasas que varían en función de las condiciones ambientales y de manejo(4,6). En gramíneas, la formación de tallos está relacionada con la tasa de aparición de hojas, y el número de hojas determina el potencial de aparición de tallos(7). En ballico perenne, la persistencia y el rendimiento dependen del balance entre las tasas de aparición y muerte de tallos, el cual es afectado por la frecuencia e intensidad de defoliación(8). Con cortes frecuentes se reduce la competencia por luz, de modo que la respuesta del pasto es la formación de hojas cortas y alta densidad de tallos. Por el contrario, con defoliaciones poco frecuentes se desarrollan hojas largas y se reduce la tasa de aparición de tallos(9,10).
Algunos estudios(11,12), han demostrado que el rendimiento incrementa al aumentar el número y el crecimiento individual de cada tallo. Otros estudios han informado(13,14) que en praderas asociadas, los componentes que contribuyeron al rendimiento variaron durante un periodo de estudio de dos años, en el primer año el ballico perenne tuvo mayor crecimiento, mientras que en el segundo el rendimiento dependió del trébol blanco.
En México existen pocos estudios sobre dinámica de tallos y de la tasa de recambio de tejido (crecimiento neto, elongación y senescencia de las hojas) como componentes del rendimiento(15). Sin embargo, cuantificar dichas variables y su variación durante el año, genera información importante para diseñar un manejo de praderas que favorezca simultáneamente una alta utilización y persistencia de las especies forrajeras, evitando su deterioro por una excesiva acumulación de tallos y material senescente(16).
El presente estudio tuvo como objetivo evaluar la densidad, peso, tasa de aparición y muerte de tallos, además el recambio de tejido de ballico perenne, pasto ovillo y trébol blanco asociados en diferentes proporciones.
El estudio se realizó en una pradera asociada de trébol blanco (Trifolium repens L.), pasto ovillo (Dactylis glomerata L.) y ballico perenne (Lolium perenne L.), establecida en marzo del 2006, en el campo experimental del Colegio de Posgraduados, en Montecillo, Texcoco, Estado de México. Cl suelo es de textura franco arenosa, pH 7.8, 2.4 % de materia orgánica, y se clasificó como un Typic ustipsamments(17). Clima templado subhúmedo con lluvias en verano, precipitación y temperatura media anual de 645 mm y 15 oC, respectivamente(18).
Las gramíneas se sembraron en hileras con separación de 30 cm, la leguminosa se sembró perpendicular a las gramíneas, a una distancia entre hileras de 40 cm. La relación de siembra fue 4:6 con base en la densidad de siembra recomendada para asociaciones de trébol blanco y gramíneas(19). Los tratamientos consistieron en cinco proporciones de trébol blanco, ovillo y ballico perene: 4:3:3, 4:6:0; 4:0:6; 4:2:4 y 4:4:2, las cuales se distribuyeron aleatoriamente en 20 parcelas experimentales de 104 m2, en un diseño de bloques completos al azar con cuatro repeticiones. Las unidades experimentales no se fertilizaron y en la época seca se aplicaron riegos a capacidad de campo cada dos semanas. Las parcelas se delimitaron con cerco eléctrico y fueron pastoreadas por 12 ovinos adultos, hasta uniformizarlas a una altura aproximada de 5 cm de forraje residual.
La frecuencia de pastoreo fue cada cuatro semanas en primavera y verano (marzo-septiembre 2007) y seis semanas durante otoño e invierno (septiembre 2007-marzo 2008), de acuerdo con las recomendaciones para pasto ovillo y ballico perenne en monocultivo(20,21). Las alturas promedio del forraje al inicio del pastoreo fueron de 22 y 26 cm en primavera y verano, respectivamente, mientras que en otoño e invierno fueron de 11 cm, y la altura de forraje residual fue de 5 cm durante todo el experimento.
El crecimiento neto, elongación y senescencia foliar se evaluaron a mediados de cada estación del año (abril, julio y octubre de 2007 y enero del 2008). En un transepto de 2 m de largo se seleccionaron al azar cinco tallos de cada gramínea y cinco estolones de trébol blanco, los cuales se identificaron con anillos de alambre de diferente color. En los tallos se midió: la longitud de lámina foliar, desde la base de la lígula hasta el ápice en hojas verdes o hasta la base del tejido clorótico en hojas senescentes; mientras que, en los estolones se registró la distancia desde la base del raquis primario hasta el ápice de la nervadura media del foliolo central y se determinó el área foliar con un integrador foliar (LI-COR, LI-3100C USA). Se obtuvo una ecuación de regresión lineal considerando como la variable dependiente el área foliar y la variable independiente la longitud de la nervadura (modificación de la metodología de Williams et al(22).
La tasa de elongación foliar por tallo (TCF; mm tallo-1 d-1), se calculó para hojas en crecimiento, como la diferencia entre la sumatoria de las longitudes de las láminas foliares final (LFf) e inicial (LFi), dividida entre el número de días (RD), entre mediciones sucesivas(4).

La tasa de senescencia foliar por tallo (TSF; mm tallo-1 d-1) se calculó como la disminución en longitud de la hoja entre intervalos de mediciones sucesivas, dividido entre el número de días en el intervalo(4).
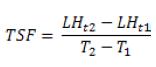
La tasa de crecimiento neto foliar por tallo (CRF; mm tallo-1 d-1), se calculó como la diferencia entre la tasa de elongación foliar (TCF) y la tasa de senescencia foliar (TSF)(4).
CRF=TCF-TSF
La densidad de tallos se midió inmediatamente después del pastoreo de uniformización, se colocaron dos aros de pvc de 10.4 cm de diámetro en cada unidad experimental; cada aro tenía dentro un macollo y cuando se tenían las tres especies asociadas, cada aro contuvo una especie de gramínea. Los tallos presentes dentro del aro se consideraron como población inicial. Posteriormente, cada mes, durante un año, los tallos emergidos se marcaron con anillos de diferente color y se registraron, y los tallos muertos se contaron y se les retiró el anillo.
Los valores de densidad, aparición y muerte de tallos se expresaron con base en un m2, y con esa información se estimó: la densidad de tallos (DT; tallos vivos m2 en cada muestreo) y sus respectivas tasas mensuales de aparición (TAT) y muerte (TMT) (%), mediante las siguientes ecuaciones:(4).

El peso por tallo se registró un día antes del pastoreo y a mitad de cada estación, se cortaron a ras de suelo 10 tallos de cada especie, se secaron en una estufa de aire forzado por 48 h a 55 oC y se registró su peso. El peso promedio por tallo se obtuvo dividiendo, el peso total entre el número de tallos cosechados.
Los registros de temperatura a la intemperie (máxima, media y mínima), y precipitación mensual , se obtuvieron de la estación climatológica del Colegio de Postgraduados, situada a 400 m del sitio experimental (Figura 1).

Los datos obtenidos se analizaron con el procedimiento estadístico PROC MIXED del software SAS(23), las medias de tratamientos fueron estimadas por LSMEANS y se compararon mediante la prueba de "t" de "Student" a una significancia (P<0.05), que se generó con la opción PDIFF. Cuando no se cumplió el supuesto de normalidad las variables fueron transformadas. Las figuras se procesaron con el software SigmaPlot de Jandel Scientific (Versión 10).
En primavera, la asociación 4:2:4 presentó la mayor elongación y senescencia foliar con 2.39 y 0.79 cm tallo-1 d-1 que superó significativamente (P<0.05), a las otras asociaciones, entre las cuales no hubo diferencias (Cuadro 1). El CNF fue similar entre asociaciones (P>0.05), aunque la combinación 4:2:4 superó en 32, 23 y 12 % a las asociaciones 4:4:2, 4:6:0 y 4:3:3, respectivamente. En verano, todas las asociaciones registraron el mayor dinamismo de recambio de tejido. La elongación y crecimiento neto foliar fueron en promedio de 6.6 y 5.9 cm tallo-1 d-1, no existiendo diferencias entre asociaciones. Sin embargo, la asociación 4:3:3 presentó la mayor S con 1.1 cm tallo-1 d-1 (P<0.05). En otoño no se registraron diferencias entre las asociaciones. En invierno, las asociaciones 4:3:3 y 4:2:4 registraron la mayor E superando en 47 % a la asociación 4:4:2. No se detectaron diferencias (P>0.05) con respecto a S y CNF. Estos resultados son similares a los obtenidos en pasto ovillo, en donde se observó el mayor índice de área foliar, crecimiento neto y rendimiento en la época de verano, en comparación con otoño, invierno y primavera (P<0.05)(20). Se informó de un comportamiento similar de pasto ovillo al evaluar su curva de crecimiento en asociación con alfalfa(1). Sin embargo, en un estudio de invernadero, el pasto ovillo presentó tasas de elongación y crecimiento neto foliar similares (P>0.05) en verano y otoño(24).
Para ballico perenne en primavera, se registraron diferencias entre asociaciones (P<0.05, Cuadro 1); la asociación 4:4:2 presentó la menor E Y CNF con 1.7 y 1.4 cm tallo-1 d-1.
La senescencia foliar fue en promedio de 0.3 cm tallo-1 d-1 (P>0.05). La mayor tasa en E y CNF se registró en verano con promedios de 7.2 y 6.7 cm tallo-1 d-1 (P>0.05), respectivamente, siendo, la asociación 4:4:2 la que presentó mayor S con 0.7 cm tallo-1 d-1, superando a la combinación 4:0:6 que tuvo el menor valor (P<0.05). En otoño no se observaron diferencias significativas en las variables E, CNF y S. Durante el invierno, la asociación 4:0:6 presentó la mayor E y CNF con 1.4 y 1.1 cm tallo-1 d-1, respectivamente, superando en 45 y 78 % a la asociación 4:4:2 (P<0.05). Se ha mencionado(25,26) que E y S en ballico perenne están correlacionadas con la temperatura ambiental y que la producción neta de forraje es negativa a bajas temperaturas, pues valores < a 6 o C afectan negativamente la E y CNF. Asimismo, se ha informado(27) que la elongación es mayor en verano porque esta época presenta las temperaturas óptimas de crecimiento (18 a 25 oC), por lo mismo, en verano se obtuvieron los mayores rendimientos de forraje.
El trébol blanco presentó las mayores E y CNF, durante primavera y verano (Cuadro 2). En primavera, no se observaron diferencias entre asociaciones en E y CNF, registrándose sólo diferencias (P<0.05) en la senescencia foliar; la asociación con 4:6:0 presentó el mayor valor con 0.05 cm2 hoja-1 d-1. En verano se registraron diferencias en E y CNF, y sólo la asociación 4:6:0 fue (P<0.05) diferente, sin que se registrasen diferencias en senescencia de hojas. En otoño, a excepción de la senescencia foliar no se observaron diferencias entre tratamientos; mientras que en invierno, la asociación 4:0:6 (0.3 cm2 hoja-1 d-1) fue inferior y diferente al resto de los tratamientos (P<0.05).
La mayor elongación y crecimiento neto del peciolo de trébol blanco se observó en primavera y verano (Cuadro 2); en invierno no se registraron diferencias. La senescencia no registró diferencias entre asociaciones en todas las estaciones del año. En primavera la asociación 4:2:4 registró el valor mayor en E y CN del peciolo (2.01 y 1.92 cm2 hoja-1 d-1) siendo superior en 60 y 58 %, respectivamente a la asociación 4:6:0. En verano la asociación 4:4:2 superó en 42 y 45 % a las asociaciones 4:3:3 y 4:6:0 en E y CN del peciolo (P<0.05), respectivamente. En otoño se observó la menor E y CN del peciolo; estos resultados son similares a los reportados en trébol blanco asociado con ballico perenne(4), en donde se observaron diferencias (P<0.05) en la tasa de elongación de peciolo y expansión foliar del trébol blanco, al variar el intervalo de pastoreo, independientemente del tratamiento asignado, en experimentos donde dejaron un forraje residual de 900-1000 kg MS ha-1 (cada 3 semanas), 1500-1700 kg MS ha-1 (cada 6 y 12 semanas).
Los resultados en E, CNF y S en las asociaciones con ovillo, coinciden con otros estudios, que informan que en praderas asociadas, el ovillo afecta la velocidad de rebrote del trébol blanco, debido a la competencia interespecífica, la cual influye en el tamaño de los foliolos y estolones y en la distribución espacial del dosel vegetal del trébol blanco(28).
La velocidad con la que un forraje se recupera después de una defoliación, depende de los carbohidratos de reserva en raíces y el numero de yemas vegetativas iniciadas, lo que favorece un crecimiento foliar acelerado, y por consecuencia una mayor intersección de luz, lo que incrementa la capacidad fotosintética de las hojas; y una mayor elongación y crecimiento neto foliar(29).
Las condiciones ambientales determinan la tasa de crecimiento foliar del trébol blanco, la cual es particularmente alta en primavera-verano(30,31). Pero mucho menor en otoño-invierno debido al estrés ocasionado por bajas temperaturas y menor fotoperiodo, que provoca que se presente más del 70 % del total de la senescencia, principalmente de la parte apical(32). La E y CN del trébol blanco es más baja que la de los pastos templados al inicio de primavera, pero en verano el trébol tiene una tasa de sobrevivencia y crecimiento mayor debido a que se encuentra en sus temperaturas optimas de crecimiento (< 24 oC) (Figura 1), lo que restablece el equilibrio de la especie(32).
Independientemente de la asociación, la mayor E y CNF del trébol blanco, ovillo y ballico perenne se presentó en primavera y verano y las menores en otoño e invierno, y se atribuyen a que, en los meses de abril a septiembre se registraron los rangos de temperatura óptimos para el crecimiento, mientras que de octubre a febrero las temperaturas más bajas lo que retrasa el crecimiento de las especies evaluadas (Figura 1)(20,21,33,34).
La asociación 4:3:3 registró (P<0.05) el mayor peso por tallo de ballico perenne en primavera, verano e invierno (Cuadro 3). El promedio anual de esta asociación superó en 8, 18 y 116 % (P<0.05) a las asociaciones 4:4:2, 4.2:4 y 4:0:6, respectivamente. En verano y otoño se registró el mayor y el menor peso promedio por tallo; 0.24 y 0.12 g tallo-1 respectivamente, y se registraron diferencias en las otras épocas del año (P<0.05). La asociación 4:6:0 registró el menor peso por tallo en las otras épocas del año (Cuadro 3). Estos resultados coinciden con los de otros autores(4,15), quienes observaron el mayor peso de los tallos de ballico perenne en verano, independientemente de la frecuencia de corte; pero difieren a otros obtenidos en praderas puras con diferentes frecuencias de pastoreo(27), donde reportaron que en la época de invierno no existieron diferencias entre tratamientos.
La asociación con el 4:2:4 presentó el mejor peso promedio anual de pasto ovillo (0.25 g tallo-1), y fue mayor (P<0.05) al resto de las asociaciones en primavera y verano (0.31 y 0.38 g tallo-1), superando en 280 y 225 % a la asociación 4:6:0 (Cuadro 3). Durante el otoño no se observaron diferencias entre tratamientos. En invierno, la asociación 4:6:0 fue menor y diferente al resto de las asociaciones (P<0.05). Los resultados del experimento confirman lo informado por otros autores(4,20), quienes evaluaron curvas estacionales de crecimiento de esta especie, y observaron que en primavera y verano se presenta la mayor cantidad de biomasa producida, lo cual está estrechamente correlacionado con en número y peso por tallo(4,20).
Para la densidad de tallos los resultados no muestran diferencias entre tratamientos durante los diferentes meses del año; sin embargo, en ambas especies, independientemente de la asociación, a inicios de primavera (abril) y en otoño (octubre y noviembre) se presentaron las mayores densidades de tallos (Figura 2). Lo anterior coincide con otro estudio(35) que reportó las mayores densidades de tallos en la época de menor temperatura, en las mismas especies.
El ballico perenne no registró diferencias durante el experimento y mostró dos picos en la aparición de tallos (Figura 3A); el primero fue en abril y el segundo pico se presentó en noviembre, siendo la asociación 4:2:4 la que mostró el valor más alto (6.59 tallos*100 tallos d-1). Estudios en praderas de ballico perenne(15), reportaron las mayores tasas de aparición de tallos en verano e invierno, las cuales duplicaron a las de otoño y primavera, no coincidiendo con los resultados observados en este estudio, ya que las mayores tasas de aparición ocurrieron después de una helada presentada en octubre (Figura 1). Así mismo, otros estudios(8) respaldan este trabajo, al demostrar que el ballico perenne promueve una rápida formación de tallos con defoliaciones frecuentes.
La tasa de mortalidad de tallos de pasto ballico perenne varió considerablemente entre asociaciones durante el periodo de evaluación (Figura 3B) y sólo se registraron diferencias estadísticas (P<0.05) en abril. En octubre se registró la mayor mortalidad con 4.3 tallos*100 tallos d-1. Estos resultados difieren a los obtenidos por otros autores(27), quienes reportaron la mayor tasa de muerte en verano (septiembre) y especularon que esto se debe a la competencia por luz y nutrientes o a la defoliación severa de tallos reproductivos que son la fuente de nutrientes de los tallos jóvenes. En este experimento la mortalidad de tallos se atribuye principalmente a la presencia de heladas y a la defoliación severa y selectiva de los ovinos.
En pasto ovillo se registraron diferencias significativas (P<0.05) entre asociaciones durante el invierno (diciembre, enero y febrero) (Figura 4A), siendo la asociación 4:4:2 la que registró el valor promedio más alto (6.3 tallos * 100 tallos d-1). Al igual que en ballico perene, se observaron dos picos de aparición en abril y noviembre. El mayor crecimiento de tallos (P<0.05) en todas las asociaciones se presentó en enero y febrero (6.2 tallos*100 tallos d-1 en promedio).
Las especies evaluadas mostraron un comportamiento similar en respuesta a las condiciones ambientales; de esta forma se confirma que el rebrote y los componentes del rendimiento de las plantas forrajeras dependen de los factores ambientales(36). Otro factor que influye en la dinámica de tallos es la competencia intraespecífica de tallos en el mismo macollo y al crecimiento individual de cada tallo, lo que provoca sombréo a los tallos pequeños incrementando su mortalidad(37).
Se concluye que el ballico perenne, el pasto ovillo y el trébol blanco presentaron una marcada estacionalidad en la elongación foliar, crecimiento neto y senescencia, con mayor crecimiento y tasa de recambio de tejidos en primavera y verano, y la menor durante otoño-invierno. Las asociaciones con las tres especies evaluadas registraron mayores valores de elongación, crecimiento neto, senescencia y dinámica de tallos en comparación con las que sólo se asociaron dos especies, siendo las mejores proporciones 4:2:4 y 4:3:3. El ballico perenne presentó mayores tasas de aparición de tallos en relación al pasto ovillo, lo que explica las mayores tasas de recambio de tejido en las asociaciones donde el ballico estuvo en mayor proporción. Tratamientos con las tres especies presentaron mayores valores en las variables evaluadas, por lo que se puede recomendar una asociación 4:2:4 ó 4:3:3 en praderas de clima templado.
LITERATURA CITADA
1. Zaragoza EJ, Hernández GA, Pérez PJ, Herrera HJG, Osnaya GF, Martínez HPA, González MS, Quero CAR. Análisis de crecimiento estacional de una pradera asociada alfalfa-pasto ovillo. Téc Pecu Méx 2009;47(2):173-188. [ Links ]
2. Barrientos DL, Montenegro BA, Pino NI. Evaluación de la fijación simbiótica de nitrógeno de Lupinus albus y L. angustifolius en un andizol vilcun del sur de Chile. Terra Lat 2002;(20):001. [ Links ]
3. Davies A. Competition between grasses and legumes in Established Pastures. In: Competition and succession in pastures. CABI Publishing UK. 2001. [ Links ]
4. Hernández GA, Hodgson JG, Matthew C. Effect of spring grazing management on perennial ryegrass and ryegrass-white clover pastures. 1. Tissue turnover and herbage accumulation. N Z J Agric Res 1997;(40):25-35. [ Links ]
5. Hernández-Garay A, Matthew C, Hodgson, J. The influence of defoliation height on dry-matter partitioning and CO2 exchange of perennial ryegrass miniature sward. Grass Forage Sci 2000;(54):1-5. [ Links ]
6. Hodgson J. Grazing management. Science into practice. Harlow, England: Longman Scientific and Technical; 1990. [ Links ]
7. Davies A, Simons RG. Effect cutting regime on developmental morphology and spring growth of perennial ryegrass. J Agric Sci 1979;92:457-459. [ Links ]
8. Hernández GA, Matthew C, Hodgson J. Tiller size/density compensation in perennial ryegrass miniature swards subject to differing defoliation heights and a proposed productivity index. N Z J Agri Res 1999;(54):347:356. [ Links ]
9. Mazzanti A, Lemaire G. Effect of nitrogen fertilization on the herbage production of tall fescue swards grazed continuously with sheep. I. Consumption and efficiency of herbage utilization. Grass Forage Sci 1994;49:352-359. [ Links ]
10. Lemaire G. Ecophysiology of grasslands Aspects of forage plant populations in grazed swards. Proc XIX Int Grassland Congr. Brazilian Soc Anim Husb Sociedade Brasileira de Zootecnia. Sao Pedro, San Paulo. Brasil. 2001. [ Links ]
11. Matthew C, Quilter SJ, Korte CJ, Chu ACP, Macckay AD. Stolon formation and significance for tiller dynamics in perennial ryegrass. Proc N Z Grassland Assoc 1989;(50):255-259. [ Links ]
12. Matthew C, Xia JX, Hodgson J, Chu ACP. Effect of late spring grazing management on tiller age profiles and summer-autumn pasture growth rates in a perennial ryegrass (Lolium perenne L.) sward. Proc XVI Int Grassland Cong. 1989. [ Links ]
13. Da Silva SC, Matthew C, Matthews PNP, Hodgson J. Influence of spring grazing management on summer and autumn production of dairy pastures. Proc XVII Int Grassland Cong. 1993. [ Links ]
14. Da Silva SC, Hodgson J, Matthews PNP, Matthew CW. Effect of contrasting spring grazing management on summer-autumn pasture and milk production of mixed ryegrass-clover dairy swards. Proc N Z Soc Anim Prod 1994:54:79-82. [ Links ]
15. Velasco ZME, Hernández GA, González HVA. Cambios en componentes del rendimiento de una pradera de ballico perenne, en respuesta a la frecuencia de corte. Rev Fitotec Mex 2007;30(1):79-87. [ Links ]
16. Hodgson J, Da Silva SC. Options in tropical pasture management. Proc Ann Meet Brazilian Soc Anim Sci. Recife, Brazil. 2002:180-202. [ Links ]
17. Ortiz SC. Colección de monolitos. Montecillo, Texcoco, Edo. México. México: Depto. Génesis de suelos. Edafología, IRENAT. Colegio de Postgraduados. 1997. [ Links ]
18. García E. Modificaciones al sistema de clasificación climática de Köppen. 4a ed. Universidad Nacional Autónoma de México. México: Ed. Limusa; 1988. [ Links ]
19. White J, Hodgson J. Pasture establishment. In: Pasture and crop science. New Zealand: Oxford University Press; 1999:104-106. [ Links ]
20. Velasco ZME, Hernández GA, González HVA, Pérez PJ, Vaquera HH, Galvis SA. Curva de crecimiento y acumulación estacional del pasto ovillo (Dactylis glomerata L.). Téc Pecu Méx 2001;39(1):1-14. [ Links ]
21. Velasco ZME, Hernández GA, González HVA. Rendimiento y valor nutritivo de Ballico perenne (Lolium perenne L.) en respuesta a la frecuencia de corte. Téc Pecu Méx 2005;43(2):274:258. [ Links ]
22. Williams RF, Evans LT, Ludwig LJ. Estimation of leaf area for clover and Lucerne. Aust J Agric Res 1994;15:231-233. [ Links ]
23. SAS. User's Guide: Statistics (version 9.0 ed.). Cary NC, USA: SAS Inst. Inc. 2002. [ Links ]
24. Wilson GCY, Zavaleta MHA, López DH, Hernández GA. La citoquinina BAP retrasa la senescencia, aumenta antioxidantes, proteína y crecimiento en el pasto ovillo (Dactylis glomerata L.). Agrociencia 2008:42:799-806. [ Links ]
25. Brereton JA, Carton OT, O'Keffe WF. Tissue turnover in Perennial Ryegrass (Lolium perenne L.) during winter. J Agric Res 1985;24:49-62. [ Links ]
26. Hennessy D, O'Donovan M, French P, Laidlaw AS. Factors influencing tissue turnover during winter in perennial ryegrass-dominated swards. Grass Forage Sci 2008;63(2):202-211. [ Links ]
27. Garduño VS, Pérez PJ, Hernández GA, Herrera HJG, Martínez HPA, Joaquin TBM. Rendimiento y dinámica de crecimiento estacional de ballico perenne, pastoreado con ovinos a diferentes frecuencias e intensidades. Téc Pecu Méx 2009;47(2):189-202. [ Links ]
28. Sanderson MA, Elwinger GF. Plant density and environment effects on Orchardgrass - White clover mixtures. Crop Sci 2002;42:2055-2063. [ Links ]
29. Lemaire G, Agnusdei M. Leaf tissue turnover and efficiency of herbage utilization. Grassland Ecophysiol Grazing Ecol. CABI Publishing UK. 2000. [ Links ]
30. Hay RJM. Seasonal variation in the distribution of white clover stolons among three horizontal strata in two grazed swards. N Z J Agric Res 1983;26:29-34. [ Links ]
31. Hay RJM, Ryan DL. A review of 10 years' research with red clovers under grazing in Southland. Proc N Z Grassland Assoc 1989;50:181-187. [ Links ]
32. Brock JL, Tilbrook JC. Effect of cultivar of white clover on plant morphology during the establishment of mixed pastures under sheep grazing. N Z J Agric Res 2000;43:335-343. [ Links ]
33. Clark H, Newton PCD, Bell CC, Glasgow EM. The influence of elevated CO2 and simulated seasonal changes in temperature on tissue turnover in pasture turfs dominated by ryegrass (Lolium perenne) y white clover (Trifolium repens). J App Ecol 1995;32:128-136. [ Links ]
34. Brock JL, Caradus JR, Hay MJM. Fifty years of white clover research in New Zealand. Proc N Z Grassland Assoc 1989;50:25-39. [ Links ]
35. Ganderats FS, Hepp KC. Mecanismos de crecimiento de Lolium perenne, Festuca arundinacea y Dactylis glomerata en la zona intermedia de Aysén. Agric Téc 2003;63(2):259-265. [ Links ]
36. Durand JL, Schaufele R, Gastal F. Grass leaf elongation rate as a function of developmental stage and temperature: Morphological analysis and modeling. Ann Bot 1999;83:577-588. [ Links ]
37. Matthew C, Hernández-Garay A, Hodgson J. Making sense of the link between tiller density and pasture production. N Z Grassland Assoc 1996;57:83-87. [ Links ]

