










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista mexicana de ciencias pecuarias
On-line version ISSN 2448-6698Print version ISSN 2007-1124
Rev. mex. de cienc. pecuarias vol.4 n.2 Mérida Apr./Jun. 2013
Artículos
Reconocimiento mutuo madre-cría en ovinos Columbia con restricción nutricional durante la gestación
Mother-young mutual recognition in Columbia sheep with nutritional restriction during pregancy
Alan Olazábal Fenochioa, Héctor Raymundo Vera Ávilab, Norma Serafín Lópezc, José Alfredo Medrano Hernándeza, Hitandewy Sánchez Saucedo, Angélica María Terrazas Garcíaa,c
a Departamento de Ciencias Pecuarias, Facultad de Estudios Superiores Cuautitlán, Universidad Nacional Autónoma de México. México. garciate@unam.mx. Correspondencia al último autor.
b Centro Nacional de Investigación en Fisiología y Mejoramiento Animal, Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP).
c Departamento de Neurobiología Conductual y Cognitiva, Instituto de Neurobiología, Universidad Nacional Autónoma de México (UNAM).
Recibido el 22 de octubre de 2010.
Aceptado el 14 de abril de 2011.
Resumen
Se evaluó en ovejas Columbia multíparas la capacidad de reconocer a sus crías (8 y 12 h postparto) y en éstas la capacidad de reconocer a sus madres (12 y 24 h post nacimiento) y las características espectrales de sus balidos (24 y 48 h post nacimiento). Se establecieron dos grupos experimentales, uno con restricción nutricional durante la segunda mitad de la gestación (grupo restringido, 70 % de requerimientos de energía y proteína a partir del día 70 post concepción, n=24) y el otro sin restricción nutricional (grupo control, 100 % de requerimientos durante toda la gestación, n=11). Las ovejas del grupo restringido no mostraron capacidad de discriminar a su cordero de uno ajeno a 8 h postparto (P>0.05), pero sí a las 12 h. En contraste las ovejas del grupo control discriminaron a su cría de una ajena desde las 8 h postparto pues permanecieron más tiempo cerca de ésta y la miraron por más tiempo (P<0.05). En los corderos, los del grupo restringido no fueron capaces de discriminar a su madre de una ajena a 12 post nacimiento (P>0.05), pero sí a las 24 h. En contraste, los corderos del grupo testigo fueron capaces de discriminar a su madre de una ajena desde las 12 h post nacimiento (permanecieron más tiempo cerca de su madre y la miraron por más tiempo comparado con una madre ajena, P<0.05). Las características espectrales de las vocalizaciones en los corderos no fueron influenciadas por el tratamiento nutricional de sus madres durante la gestación (P>0.05). Se concluye que la restricción nutricional durante la gestación retrasa el reconocimiento madre-cría sin alterar aparentemente las características de las vocalizaciones en las crías.
Palabras clave: Desnutrición, Motivación materna, Afiliación, Ovinos, Vocalizaciones.
Abstract
It was evaluated in multiparous Columbia ewes their ability to recognize their offspring (8 or 12 h postpartum), and in lambs their ability to recognize their mothers (12 or 24 h after birth), plus the spectral characteristics of the bleating of the litter (24 and 48 h of age). There were two experimental groups, one with nutritional restriction during the second half of gestation (Restricted, 70 % of their requirements in protein and energy from d 70 post conception, n= 24) and another without nutritional restriction (Control, 100 % of their requirements during pregnancy, n= 11). Nutritional restricted mothers to 8 h, showed no ability to discriminate the own and alien lamb (P>0.05), but at 12 h, they could do so. While control females were able to discriminate its own lamb from an alien one since 8 h, they spent more time near their offspring than the others and looked them for longer period (P<0.05). While lambs from restricted group at 12 h were not able to discriminate against an alien mother (P<0.05), but they could do so at 24 h. While lambs of control group were able to discriminate against an alien mother since 12 h after birth (spend longer time near their mother and looked for longer time compared to the alien dam, P<0.05). The spectral characteristics of the vocalizations of the lambs were not influenced by the nutritional treatment of their mothers during pregnancy P<0.05). It was concluded that nutritional restriction during gestation delays the recognition mother-lamb, without alters apparently the spectrum of vocalizations in lambs.
Key words: Undernutrition, Maternal motivation, Affiliation, Lambs, Ewes, Vocalizations.
INTRODUCCIÓN
En ovinos como en los demás mamíferos, el despliegue adecuado de la conducta materna es fundamental para la sobrevivencia de la cría(1). En la oveja se establece un vínculo maternal selectivo al parto, por lo cual entre 30 a 120 min post parto, la gran mayoría rechaza a cualquier cría con la cual no ha tenido interacciones y amamanta exclusivamente a su propia cría(1-3).
Se ha establecido que el reconocimiento olfatorio es el factor crítico para que la madre permita el acceso de la cría a la ubre, sin embargo, éste no es el único mecanismo por el cual la madre identifica a su progenie. En efecto, se ha demostrado que las ovejas son capaces de reconocer a su cría a distancia, es decir sin la ayuda de señales olfatorias(4), y se piensa que lo hacen con otras señales como son las visuales y auditivas(4,5,6).
Actualmente, se sabe que tanto en la oveja como en la cabra las madres pueden discriminar entre su cría y una ajena sin la ayuda del olfato, esto desde las primeras 12 h después del parto, e incluso antes en algunos casos(7-10).
La capacidad en los corderos para discriminar entre su madre y una ajena también ha sido investigada. Se ha demostrado que desde las 12 h de edad, los corderos pueden mostrar una clara preferencia por su madre cuando se les da a elegir entre ésta y una ajena(11-13). En otros estudios, se ha observado que a las 24 h de vida los corderos son capaces de discriminar a su madre de una ajena, a través de reconocer y preferir un comportamiento de aceptación que muestra la madre propia y evitar uno de rechazo que muestra la madre ajena(14).
Además, en ovejas se demostró que las madres fueron capaces de reconocer a su cordero a las 24 h post parto sólo percibiendo sus vocalizaciones, mientras que los corderos son capaces de reconocer a su madre de esta forma desde las 48 h post nacimiento(15). De igual manera, se ha demostrado que tanto las ovejas como los corderos poseen una firma acústica que permite reconocerse entre ellos desde la primera semana post parto(16,17).
En estudios recientes en cabras, se encontró que la desnutrición en la gestación deteriora el reconocimiento de la cría a distancia por las madres a las 8 h post parto. Ese mismo efecto se observó en los cabritos nacidos de hembras nutricionalmente restringidas, ya que tuvieron problemas para discriminar entre su madre y una ajena(18,19). Poco se ha estudiado sobre el efecto de la desnutrición en ovejas sobre la vinculación madre-cría(20). Por ejemplo, ovejas pastoreando praderas de baja calidad fueron más susceptibles a abandonar a una de sus crías en el caso de partos dobles, comparado con ovejas bien alimentadas(21,22). Asimismo, ovejas restringidas nutricionalmente durante la gestación, estuvieron menos vinculadas con su cordero en los primeros días post parto que hembras bien alimentadas(20).
El objetivo del presente trabajo fue determinar si la condición nutricional de las ovejas durante la gestación influye en la capacidad de reconocimiento mutuo entre madres y corderos poco después del parto. La hipótesis experimental fue que la restricción nutricional durante la segunda mitad de la gestación afecta el evento de reconocimiento distal madre-cría y el componente que representan las vocalizaciones de la cría como parte de éste.
MATERIALES Y MÉTODOS
El presente trabajo fue aprobado por el Subcomité Institucional para el Cuidado y Uso de Animales Experimentales del Posgrado en Ciencias de la Salud y la Producción Animal-UNAM.
El estudio se realizó en el Centro de Enseñanza Agropecuaria de la Facultad de Estudios Superiores Cuautitlán, UNAM, localizado a 19° 41' 15" N, 99° 11' 45" O y 2,252 msnm. Se utilizaron inicialmente 50 ovejas de la raza Columbia, multíparas, de entre 3 a 5 años de edad, las cuales fueron apareadas a un estro sincronizado(18), obteniéndose en total 35 animales gestantes.
A la octava semana post concepción, los animales gestantes fueron asignados a un grupo testigo (n=11), o a uno de restricción nutricional (n=24), procurando una distribución uniforme en los grupos de acuerdo al peso corporal.
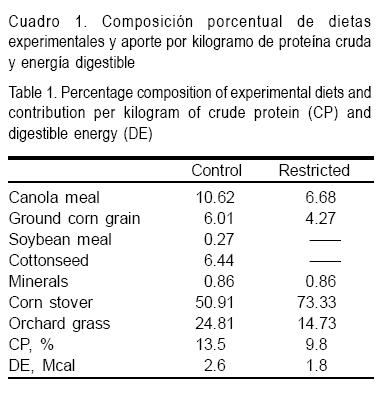
En el grupo testigo, las hembras fueron alimentadas durante toda la gestación con una dieta formulada para aportar el 100 % de sus requerimientos nutricionales teóricos(23). El manejo de la alimentación en el grupo restringido fue similar al del grupo testigo hasta el día 70 ± 5 de gestación; a partir de ese momento y hasta el parto, se redujo a 70 % el aporte de los requerimientos teóricos de energía y proteína, de acuerdo al consumo voluntario esperado. Una vez que se realizó el cambio de dieta en el grupo restringido, durante las dos primeras semanas se llevó a cabo una estimación del consumo voluntario promedio en cada grupo [(alimento ofrecido - alimento rechazado)/número de ovejas en el grupo]. El consumo voluntario promedio observado fue de 1.59 y 1.57 kg/oveja en los grupos testigo y restringido, respectivamente, manteniéndose de acuerdo a esto el aporte de nutrientes mayores establecidos para cada grupo según el diseño experimental. La cantidad de alimento ofrecido fue siempre suficiente para asegurar un consumo ad libitum; la composición de la dieta para cada grupo experimental y su aporte de energía digestible y proteína cruda, se presentan en el Cuadro 1.
En las ovejas se registraron los pesos corporales los días 70, 117 y 138 de gestación y en ovejas y corderos inmediatamente después del parto/ nacimiento. Adicionalmente, en las ovejas se colectaron muestras de sangre durante la gestación (70, 117 y 138 días) y en éstas se determinó por radioinmunoanálisis en un solo ensayo la concentración sérica de insulina (Coat-A-Count Insulin, DPC, Los Angeles, CA). La sensibilidad del ensayo fue de 0.064 ng/ml y los coeficientes de variación para los controles alto y bajo, 10.2 y 5.2 % respectivamente.
La totalidad de partos ocurrió en un lapso de 9 días (31 de marzo a 8 de abril), presentándose 6 partos sencillos y 5 dobles en el grupo testigo y 3 sencillos y 21 dobles en el grupo restringido nutricionalmente. La distribución de crías de acuerdo a su sexo fue de 53.3 % hembras y 46.6 % machos en el grupo testigo y 52.9 % hembras y 47.1 % machos en el grupo restringido. Después del parto cada madre y su(s) cría(s) se colocaron durante las primeras 4 h postparto en un corral de 2 x 2 m, delimitado con paneles, donde a la madre se le proveyó de alimento suficiente y agua a libre acceso. Posteriormente, madres y crías fueron transferidas a un corral con otras hembras paridas en espera de realizar evaluaciones conductuales de reconocimiento mutuo. En cuanto a la capacidad de reconocimiento mutuo oveja-cría, se acordó evaluarla a partir de las 8 h post parto en las ovejas(7) y de las 12 h post nacimiento en las crías(14), según lo observado en estudios previos.
Pruebas de reconocimiento de las crías por sus madres a las 8 o 12 h post parto
En las madres se realizó una prueba de reconocimiento a distancia de la cría propia a 8 ó 12 h post parto, similar a la descrita por Terrazas et al(7) y Ramírez et al(24), en donde se utilizó un corral de prueba de las mismas características y se registraron las variables conductuales descritas en esos estudios. Adicionalmente, se registró el número de balidos emitidos por el animal durante su prueba. Se probaron en total 11 ovejas testigo (5 animales a 8 h y 6 animales a 12 h), y 24 restringidas (10 animales a 8 h y 14 animales a 12 h).
Pruebas de reconocimiento de la madre por las crías a las 12 o 24 h post nacimiento
A las 12 h esta prueba se realizó en 9 corderos nacidos de ovejas testigo (4 de parto sencillo y 5 de parto doble), así como 17 nacidos de ovejas restringidas (2 de parto sencillo y 15 de parto doble). A las 24 h la prueba se realizó en 6 corderos de ovejas testigo (uno de parto sencillo y 5 de parto doble) y 17 corderos de ovejas restringidas (uno de parto sencillo y 16 de parto doble). Los corderos se sometieron a una prueba de elección doble para evaluar la preferencia por su madre(14,24), el corral utilizado fue de las mismas características y se registraron las mismas variables conductuales, más el número de balidos emitidos por el animal durante su prueba.
Análisis de vocalizaciones de corderos
A las 24 y 48 h post nacimiento se grabaron vocalizaciones de corderos sometidos a las pruebas conductuales de reconocimiento. Se registraron y anal izaron 10 balidos altos (vocalizaciones que emitía el animal con el hocico abierto), de 10 corderos nacidos de ovejas testigo (3 de parto sencillo y 7 de parto doble) y de 15 corderos nacidos de ovejas restringidas (1 de parto sencillo y 14 de parto doble). La limpieza de las grabaciones y el subsecuente análisis espectral se realizó en el laboratorio, y el proceso fue similar al descrito por Terrazas et al(25).
El procedimiento para efectuar las grabaciones fue el siguiente: con el fin de estimular las vocalizaciones, los corderos se separaron de sus madres en un corral individual de 1 x 1 m, construido con paneles abiertos y a una distancia de 10 m del corral de las madres. Las crías se grabaron durante una sesión de 10 min con la ayuda de un micrófon o omnidireccional de ambiente (Marca Shure, SM89, Presonus Audio Electronics, USA), el cual se colocó a una distancia aproximada de 50 cm del suelo y del cordero. Una vez terminada la grabación el cordero se llevó con su madre.
Posteriormente, la información se analizó en el laboratorio con la ayuda del programa Sound Forge 4.5 (Sonic Foundry, Inc., Madison, WI, USA). Cada balido o vocalización se aisló individualmente y limpiado de cualquier ruido o contaminante, y con el programa mencionado se obtuvo un sonograma en el que se estimó el poder del espectro, mediante el cual la vocalización se muestra en un eje de frecuencia y amplitud. Los parámetros medidos en el poder del espectro fueron los siguientes.
Duración del balido (seg): este parámetro se midió directamente sobre el sonograma y corresponde a la duración de la frecuencia fundamental y armónicas. Número de armónicas: corresponde al número de resonancias (armónicas) de la frecuencia fundamental, que se registraron en el sonograma. Frecuencia máxima (kHz): corresponde a la frecuencia de la armónica con la mayor cantidad de energía. Frecuencia de la última armónica (kHz): corresponde a la frecuencia pico de la última armónica. Para cada uno de los parámetros anteriores, se obtuvo el promedio de los 10 balidos registrados en cada cordero y éste fue el valor utilizado como variable de respuesta.
Análisis estadístico
Debido a que los datos conductuales y de características de las vocalizaciones no se ajustaron a una distribución normal (Prueba de Shapiro-Wilk), se procedió a utilizar estadística no paramétrica para su análisis. Las comparaciones entre grupos experimentales dentro de tipo de cría o madre se realizaron con la Prueba U de Mann-Whitney; las comparaciones entre tipos de cría o madre dentro de grupo experimental con la Prueba de Wilcoxon para rangos. En el caso de las variables conductuales se probó a su vez el efecto de tipo de parto, sencillo vs doble. Los pesos corporal es de ovejas y corderos y las concentraciones séricas de insulina en ovejas se analizaron por ANOVA considerando como factores al grupo de tratamiento (testigo, restringido), tipo de parto (sencillo, doble) y su interacción; en su caso la separación de medias se realizó con el procedimiento de Tukey.
Todas las pruebas estadísticas se realizaron con el paquete estadístico Systat, versión 13.0 (Evanston, IL, USA). Los resultados conductuales y de los sonogramas se presentan como medianas y rangos inter cuartiles y los de peso corporal y concentración sérica de insulina como medias de cuadrados mínimos y error estándar de la media. El valor de probabilidad considerado como indicativo de significancia fue de P<0.05.
RESULTADOS
Peso corporal de ovejas y corderos y concentraciones séricas de insulina en ovejas
El peso corporal de las ovejas (Cuadro 2), fue similar en los grupos testigo y restringido a los 70 y 117 días de gestación (P>0.05), sin embargo, fue significativamente mayor (P<0.05) en el grupo testigo comparado con el restringido a los 138 días de gestación (+11.3 %, P=0.003) y al parto (+8.1 %, P=0.04). Por otra parte, ni el tipo de parto (sencillo vs doble) posterior al tratamiento nutricional, ni la interacción entre grupo experimental por tipo de parto influenciaron los pesos corporales de las ovejas durante la gestación o al parto (P>0.05). En las ovejas, las concentraciones séricas de insulina durante la gestación presentaron un comportamiento similar a sus pesos corporales; diferencia entre grupos experimentales sólo a los 138 días de gestación (0.47 ± 0.04 vs 0.27 ± 0.04 ng/ml en grupo testigo vs restringido, respectivamente; P=0.001) y ausencia de efecto de la interacción grupo experimental por tipo de parto. Por su parte, el tipo de parto como efecto principal sí tuvo influencia (P<0.05) sobre las concentraciones séricas de insulina a los 117 y 138 días de gestación, con concentraciones mayores en las ovejas que tuvieron parto sencillo derivado de esa gestación (0.50 ± 0.06 vs 0.29 ± 0.04 y 0.45 ± 0.05 vs 0.29 ± 0.03 en partos sencillos vs dobles a los 117 y 138 días de gestación, respectivamente).
En los corderos, el peso al nacimiento fue influenciado por el grupo experimental y por la interacción grupo experimental por tipo de parto (P<0.01), mas no por el tipo de parto como efecto principal (P>0.05). Los corderos del grupo testigo de parto sencillo, presentaron un peso al nacimiento mayor que los corderos de parto doble (5.6 ± 0.22 vs 4.1 ± 0.21, P<0.05). También, los corderos del grupo testigo pero de parto sencillo tuvieron pesos al nacimiento mayores que los de parto sencillo y doble del grupo restringido (5.6 ± 0.22a, 3.4 ± 0.42b y 4.2 ± 0.11b, P<0.05). Mientras que los corderos testigos de parto doble y los restringidos sencillo y dobles no difirieron en el peso al nacimiento.
Prueba de reconocimiento de las crías por las ovejas a 8 ó 12 h postparto
Ocho horas posparto. No se encontraron diferencias significativas entre grupos experimentales en la cantidad de balidos altos emitidos por las ovejas durante la prueba (64 [58-78] vs 86 [64.5-94.7] para grupo testigo vs restringido, respectivamente P>0.05); sin embargo, el tipo de parto sí influenció significativamente a esta variable (63.5 [55.570] vs 85 [65-94] para parto sencillo vs doble, respectivamente; P<0.05).
En las comparaciones realizadas dentro de tipo de cría (Cuadro 3), no se encontraron diferencias entre grupos experimentales ni entre tipos de parto en las conductas emitidas por la oveja hacia los corderos. Al comparar dentro de grupo experimental las conductas emitidas por la oveja hacia la cría propia vs la ajena, se observó que en el grupo testigo las ovejas pasaban más tiempo cerca de su cría y la miraban por más tiempo comparado con la cría ajena (P<0.05), diferencia que no se observó en las ovejas del grupo restringido. Por otra parte, en ambos grupos no se encontraron diferencias (P>0.05) en el número de visitas hechas por las ovejas a la cría propia vs la ajena. Dentro de tipo de parto, solamente se observaron diferencias en el tiempo de mirar a la cría propia vs la ajena, el cual fue mayor (P<0.05) para la cría propia tanto en las ovejas de parto sencillo como en las de parto doble. El tiempo de permanencia en la cercanía y el número de visitas a la cría propia vs ajena fue similar en ambos tipos de parto (P>0.05).
Doce horas posparto. No se encontraron diferencias (P>0.05) entre el número de balidos altos emitidos por las ovejas testigo vs restringidas (79.5 [39.2-120.2] vs 92 [83.2108.7]) ni entre las ovejas de parto sencillo vs doble (89 [76.5-131.5] vs 89 [73-108]). Asimismo, dentro de tipo de cría (Cuadro 3), no se observaron diferencias (P>0.05) entre grupos experimentales, ni entre tipos de parto, en las conductas emitidas por las ovejas hacia los corderos.
En las comparaciones dentro de grupo experimental o tipo de parto (Cuadro 3), se observó que las ovejas pasaban significativamente (P<0.05) mayor tiempo en la cercanía, miraban por más tiempo y visitaban más veces a su cría propia vs la ajena, excepto dentro del grupo control para el número de visitas por cría en donde esta diferencia fue sólo cercana a ser significativa (P=0.09).
Prueba de reconocimiento de las madres por los corderos a 12 ó 24 h post nacimiento
Doce horas posparto. No se encontraron diferencias (P>0.05) entre los corderos del grupo testigo vs restringido en el número de balidos altos emitidos durante la prueba (43 [12.5-55] vs 24 [10-48] ni entre los corderos provenientes de parto sencillo vs doble (26 [12.2-52] vs 18.5 [10.7-43.2].
Asimismo, en las comparaciones realizadas dentro de tipo de madre (Cuadro 4), no se observaron diferencias (P>0.05) entre grupos experimentales en las conductas emitidas por los corderos hacia las ovejas, situación similar a la que se encontró al comparar por tipo de parto del cual provenían los corderos.
Al contrastar dentro de grupo experimental las conductas emitidas por los corderos hacia la madre propia vs la ajena (Cuadro 4), se encontró que en el grupo testigo los corderos pasaban más tiempo cerca de su madre y la miraban por más tiempo que a la madre ajena (P<0.05), diferencia que no se observó en los corderos del grupo restringido. Por otra parte, en ambos grupos no se encontraron diferencias (P>0.05) en el número de visitas hechas por los corderos a la madre propia vs la ajena. Dentro de tipo de parto, sólo los corderos provenientes de parto doble permanecieron más tiempo cerca de su madre que de la ajena (P<0.05); sin embargo, en ambos tipos de parto miraban a su vez más tiempo a su madre que a la ajena (P<0.05). El número de visitas a la madre propia vs ajena no fue diferente entre tipos de parto (P>0.05).
Veinticuatro horas posparto. Los corderos del grupo testigo emitieron menos balidos altos que los del grupo restringido (9.5 [5.7-23.5] vs 25 [15-40.5]P<0.05). Dentro de tipo de madre (Cuadro 4), no se encontraron diferencias (P>0.05) entre grupos experimentales en las conductas emitidas por los corderos hacia las ovejas. Una situación similar se observó al comparar por el tipo de parto del cual provenían los corderos, excepto en el tiempo de permanencia cerca de la madre propia, que fue mayor en corderos provenientes de parto sencillo vs de parto doble (P<0.05).
En el análisis dentro de grupo experimental, se encontró que tanto en el grupo testigo como en el restringido los corderos pasaron más tiempo en la cercanía de la madre propia vs la ajena y la miraron también por más tiempo (P<0.05). En las comparaciones por tipo de parto, sólo los corderos provenientes de parto doble pasaron más tiempo y miraron más a su madre vs la ajena (P<0.05). El número de visitas a la madre propia vs ajena no fue diferente (P>0.05) dentro de grupo experimental o tipo de parto.
Análisis sonográfico de los balidos
Veinticuatro horas de edad. El análisis espectral de las vocalizaciones de los corderos no arrojó diferencias significativas entre los grupos tratados (P>0.05), los cuales presentaron valores similares para la duración del balido (0.8 [0.7-0.9] vs 0.9 [0.8-9.95] seg), frecuencia máxima (1.4 [1.3-1.5] vs 1.4 [1.3-1.5] kHz), número de armónicas (12.1 [9.3-13.3] vs 12 [11.75-12.6]) y frecuencia pico de la última armónica (18.75 [15.8-20.1] vs 20 [19.9-20.3], kHz), para los grupos testigo y restringido respectivamente.
Cuarenta y ocho horas de edad. A esta edad, el análisis espectral de los balidos altos de los corderos tampoco arrojó diferencias entre grupos experimentales (P>0.05): duración del balido (0.8 [0.75-0.8] vs 0.8 [0.8-9.90], seg), frecuencia máxima (1.4 [1.25-1.45] vs restringido 1.5 [1.3-1.5] kHz), número de armónicas (13.3[11.6-15.1] vs 12.6 [11.1-13.5]) y frecuencia pico de la última armónica (19.7 [17.05-20.3] vs 19 [18.65-20.2], kHz) para los grupos testigo y restringidos respectivamente.
DISCUSIÓN
Los resultados del presente estudio sugieren en primera instancia que una restricción nutricional a partir de la segunda mitad de la gestación y hasta el parto, induce un atraso en la capacidad de las madres para discriminar a su cordero de uno ajeno sin la ayuda de señales olfatorias. Las hembras del grupo restringido nutricionalmente no mostraron dicha capacidad a las 8 h, pero sí lo hicieron a las 12 h postparto (a las 8 h postparto no hubo diferencia en la cantidad de tiempo que las ovejas estuvieron cerca de los corderos propios o ajenos y en el tiempo que los miraron). En cambio, las madres del grupo control fueron capaces de discriminar a su cordero de uno ajeno ya desde las 8 h postparto.
Es la primera vez que se muestra que ovejas Columbia mantenidas en estabulación y restringidas nutricionalmente durante la segunda mitad de la gestación, no son capaces de realizar un reconocimiento distal de su cría a las 8 h postparto, mientras que esta capacidad parece adquirirse hasta las 12 h. Estos resultados concuerdan con los encontrados en cabras en estabulación, las cuales habían sido restringidas nutricionalmente bajo el mismo protocolo que el presente trabajo. En ese estudio se encontró que a 8 h postparto las madres del grupo restringido no fueron capaces de mostrar una preferencia por su cabrito en una prueba de elección doble sin la ayuda de señales olfatorias(18).
En las ovejas, previamente sólo se había comprobado que la experiencia materna podía ser un factor que alterara la capacidad de reconocimiento a distancia. Así ovejas primíparas no fueron capaces de reconocer a su cría de una ajena ni a las 8 ni a las 12 h postparto, y sólo lo pudieron hacer hasta las 24 h postparto(9). Esta comparación nos permite asumir que la experiencia materna, especialmente en las ovejas primíparas, tiene un efecto importante sobre sus habilidades cognitivas. Por su parte, la restricción nutricional durante la gestación parece ejercer efectos negativos sobre estas habilidades aún después de que ya se han establecido por efecto de la experiencia maternal previa (que fue el caso de las ovejas Columbia del presente estudio). En otro trabajo en donde se evaluó el efecto de una restricción nutricional moderada, durante el primer tercio de la gestación, en ovejas que parían por primera vez, se encontró que al medir las relaciones espaciales entre la madre y su cordero durante los primeros tres días de nacidos, las ovejas del grupo restringido nutricionalmente se relacionaron menos con sus crías que las bien alimentadas(20).
Se asume que el proceso fisiológico responsable del establecimiento de una capacidad de reconocimiento sin la ayuda de señales olfatorias (registro de señales específicas del cordero durante un periodo sensible), es similar al que determina la formación del vínculo selectivo (conducta de cuidar sólo a su progenie y rechazar activamente a crías ajenas)(1,8). Este proceso fisiológico involucra la apropiada liberación de hormonas esteroideas al final de la gestación, asociada con estímulos mecánicos a nivel del canal de parto provocados por la expulsión de la cría(1). Es posible que la restricción nutricional durante la gestación altere la generación o expresión temporal de estas señales, sin embargo, es necesario realizar estudios adicionales para comprobarlo.
El tipo de parto presentado por la oveja (sencillo vs doble), no mostró una influencia sobre el desarrollo temporal de la capacidad de reconocimiento de la cría propia. Lo anterior, descarta un posible efecto confundido entre tratamiento nutricional y tipo de parto, derivado de la mayor proporción de partos dobles que se presentaron en las ovejas del grupo restringido (87.5 vs 45.4 %). No obstante, no se puede descartar por completo un posible efecto de interacción entre los factores anteriores sobre los componentes conductuales asociados al reconocimiento materno de la cría propia, efecto que no fue posible valorar en el presente trabajo.
La similitud de pesos corporales de las ovejas entre grupos experimentales al inicio del experimento (día 70 de gestación) y su disminución en el grupo restringido al final de la gestación y después del parto (pesos 8 % menores en promedio en el grupo restringido al día 138 de gestación y al parto), indican que sí se logró generar el paradigma experimental de un diferente nivel nutricional entre grupos de tratamiento. Asimismo, las concentraciones séricas de insulina, que fueron menores en el grupo restringido al final de la gestación (-42 % a 138 días de gestación), resultan indicadoras de una condición de estrés metabólico en este grupo(26). En ambos casos, el efecto de la restricción nutricional fue independiente del número de productos gestados (interacción grupo experimental x tipo de parto, no significativa), el cual como ya se mencionó no se distribuyó de manera uniforme entre grupos experimentales.
Por otra parte, los resultados obtenidos con corderos en el presente estudio, demuestran que las crías nacidas de hembras bien alimentadas tienen una preferencia clara por su madre en una prueba de reconocimiento doble a las 12 y 24 h post nacimiento, lo cual es afín a lo reportado en corderos Merino en una prueba similar(11) y en corderos Columbia probados bajo las mismas condiciones experimentales que el presente trabajo a 24 h de edad(14). Además son congruentes con los resultados en corderos Pelibuey probados a 12h(24).
Las crías son capaces de discriminar a su madre de una ajena sobre la base de su habilidad en distinguir entre la conducta de aceptación que muestra su madre durante la prueba y la de rechazo exhibida por la madre ajena(14). En contraste, nuestros resultados mostraron que la restricción nutricional materna, impide que los corderos, sean capaces de mostrar dicha habilidad a las 12 h de nacidos, mientras que sí lo pueden hacer a las 24 h de edad. Estos resultados contrastan con los reportados en cabritos probados bajo las mismas condiciones experimentales que en el presente trabajo(18). En dicho estudio se encontró que tanto a las 12 como a las 24 h de edad, los cabritos nacidos de madres restringidas nutricionalmente sí fueron capaces de mostrar una preferencia por su madre, similar a lo que se observó en los cabritos nacidos de madres testigo. Sin embargo, se localizaron ciertas diferencias con la edad, ya que los cabritos del grupo restringido nutricionalmente y probados a 12 h tendieron a estar menos activos en la prueba e hicieron menos visitas a su madre, en comparación con los cabritos probados a 24 h(18). Las diferencias anteriores entre especies, podrían estar relacionadas con diferencias de estrategia de vida postnatal, pues los cabritos son de tipo escondidizo, mientras que los corderos son de tipo seguidor(1), asimismo, podrían también reflejar diferencias entre estudios en cuanto al diseño de los corrales de prueba; ambos aspectos requerirían en su caso de una valoración experimental.
En ovinos, resultan escasos los trabajos en que se ha valorado el efecto de la condición nutricional materna durante la gestación sobre la capacidad de las crías para reconocer a su madre en forma temprana. En este sentido, existe un estudio donde se evaluó el efecto de la restricción nutricional durante los primeros 95 días de gestación (aporte de 50 % de requerimientos), sobre la reactividad emocional y la flexibilidad cognitiva a los 18 meses de edad de las crías nacidas de estas ovejas(27). Se encontró un incremento de la reactividad emocional en las crías hembra y macho y un deterioro en la flexibilidad cognitiva de las crías macho por efecto de la restricción.
Adicionalmente, la emisión de balidos por parte de las crías no fue influenciada en forma temprana por la condición nutricional de la m adre (prueba a 12 h), aunque sí posteriormente; corderos de madres restringidas emitieron más balidos a las 24 h post nacimiento. Posiblemente porque los corderos provenientes de ovejas testigo disminuyeron la emisión de balidos entre las 12 y 24 h post nacimiento. Este patrón de vocalizaciones en el grupo restringido, pudiera haberse asociado a una menor producción de leche en las ovejas restringidas nutricionalmente y reflejar entonces una necesidad de alimentación, como lo sugieren los estudios de Sébe et al(17), sin embargo, esto no se confirmó en el presente estudio.
Al igual que en las ovejas, no se encontraron evidencias de que estuvieran confundidos los efectos de tratamiento nutricional y tipo de parto sobre la capacidad de reconocimiento temprano de la madre propia por parte de la cría, esto, inducido por la mayor proporción de corderos de partos dobles probados en el grupo restringido (93.2 %) vs 66.7 % del grupo testigo. Aunque en el caso de las crías sí hubo diferencia entre grupos de tipo de parto sobre la capacidad temprana de discriminación entre la madre ajena y la propia, esta diferencia se observó sobre todo en los corderos de partos dobles, efecto que es difícil de explicar. Por otra parte, cabe resaltar que en el grupo restringido nutricionalmente hubo una mayor proporción de corderos provenientes de parto doble, y a pesar de esto, los corderos de este grupo no fueron capaces de discriminar tempranamente a la madre propia de la ajena, como ya se mencionó. Lo anterior, permite excluir la posibilidad de que estuvieran confundidos los efectos de tratamiento nutricional y tipo de parto sobre la capacidad temprana de reconocimiento de la madre propia en los corderos.
La restricción nutricional en las madres tuvo un efecto negativo sobre el peso al nacimiento de las crías, aunque sólo sobre las crías provenientes de partos sencillos y no sobre las provenientes de partos dobles (interacción grupo experimental por tipo de parto). El peso al nacimiento fue 40 % menor en corderos de parto sencillo en el grupo restringido vs el grupo testigo, mientras que los corderos provenientes de parto doble tuvieron pesos similares en ambos grupos, aunque 25 % menores que las de parto sencillo en el grupo testigo. Como se mencionó, en el grupo restringido hubo una mayor proporción de ovejas con parto doble y se probó una mayor proporción de corderos provenientes de este tipo de parto. Esto, determinó en una medida importante que los corderos del grupo restringido pesaran en promedio 1 kg menos que los del grupo testigo (4.9 vs 3.8 kg), lo cual pudiera a su vez haber determinado el retraso en el desarrollo de la capacidad de reconocimiento temprano de la madre propia observado en los corderos del grupo restringido. Sin embargo, se observó que independientemente del tratamiento nutricional, fueron sobretodo los corderos de parto doble los que pudieron reconocer tempranamente a su madre y en estos no se encontró un efecto negativo de la restricción nutricional durante la gestación sobre el peso al nacimiento. Lo anterior, sugiere que el efecto de esta restricción nutricional sobre la capacidad de discriminación entre la madre propia y la ajena en los corderos, puede estar mediado por vías que no solamente involucran al peso al nacimiento.
En cuanto a las características sonográficas de las vocalizaciones de los corderos, no se encontraron diferencias significativas entre grupos experimentales, de tal manera, que al menos bajo las condiciones descritas en el presente trabajo y con el análisis utilizado, todas las características de las vocalizaciones de los corderos que se compararon resultaron similares en ambos grupos.
Sin embargo comparativamente, se puede apreciar que las características espectrales de los animales aquí utilizados son similares a las descritas en estudios realizados en corderos Tarasconaisses x Ile de France de 3 a 16 días de edad(16). Así la duración promedio de las vocalización en el presente estudio en nuestros corderos fue de 0.82 seg, mientras el encontrado en corderos Tarasconnaises x Ile-de-France fue de 0.71 seg. Asimismo, el valor promedio de la frecuencia máxima en el presente estudio fue de 1.3 kHz, mientras que Searby y Jouventin(16) reportaron un promedio de 1.08 kHz. En otro estudio con corderos Ile-número de armónicas, han demostrado ser los parámetros más importantes en componer el timbre de la voz en ovejas y corderos, y constituyen la esencia de la firma acústica(16). A este respecto consideramos que es necesario realizar estudios ulteriores para caracterizar de manera precisa los efectos que puede tener la restricción nutricional de los corderos en la vida prenatal, sobre el desarrollo de su firma acústica.
CONCLUSIONES E IMPLICACIONES
Los resultados demuestran que en ovejas Columbia, la restricción nutricional a partir de la segunda mitad de la gestación retrasa el desarrollo en la capacidad de madres y corderos para reconocerse, en una prueba de elección doble durante el primer día postparto. Asimismo, la restricción nutricional en la vida prenatal no pareció afectar la capacidad temprana de comunicación vocal de los corderos, al menos si se consideran las variables sonográficas evaluadas en este estudio. En base a lo anterior, habría que valorar si el deterioro en el reconocimiento temprano madre-cría asociado a la restricción nutricional de la madre, está relacionado con algún otro factor como la eventual disminución de la vitalidad de las crías durante las primeras horas post nacimiento. Por otra parte, en estudios futuros será interesante dilucidar qué nutrientes son los que están mayormente relacionados con los efectos observados, si estos efectos se replican en condiciones prácticas de producción en nuestro medio, y cuáles son sus repercusiones sobre la sobrevivencia temprana de los corderos.
AGRADECIMIENTOS
Investigación realizada gracias al Programa UNAM-DGAPA-PAPIIT IN217205 Y IN207508, por el Fondo Internacional para la Ciencia FIS B/3872-1 y la cátedra FESC-GVC-15. Alan Olazabal fue becado por el CONACYT para sus estudios de maestría. Gracias al apoyo de becas UNAM-DGAPA-PASPA, para la finalización del manuscrito durante la estancia sabática de la autora responsable. Al MVZ Alejandro Hernández Barrientos por apoyar en la recopilación y análisis de los datos de las vocalizaciones en los corderos. Al MVZ Jesús Guevara Vivero y al Dr. Job Zaragoza por su asesoría en la formulación de las dietas. También agradecemos a Filiberto Hernández Vega, Rosalba Soto González, Luis Dávila Delgadillo y Sandra Viveros Mejía, por su ayuda en la recolección de datos.
LITERATURA CITADA
1. Poindron P, Levy F, Keller M. Maternal responsiveness and maternal selectivity in domestic sheep and goats: the two facets of maternal attachment. Dev Psychobiol 2007;49(1):54-70. [ Links ]
2. Lévy F, Gervais R, Kindermann U, Litterio M, Poindron P, Porter R. Effects of early post-partum separation on maintenance of maternal responsiveness and selectivity in parturient ewes. Appl Anim Behav Sci 1991;31(1-2):101-110. [ Links ]
3. Smith FV, Van-Toller C, Boyes T. The "critical period" in the attachment of lambs and ewes. Anim Behav 1966;14(1):120-125. [ Links ]
4. Lindsay DR, Fletcher IC. Sensory involvement in the recognition of lambs by their dams. Anim Behav 1968;16(4):415-417. [ Links ]
5. Alexander G, Shillito EE. The importance of odour, appearance and voice in maternal recognition of the young in Merino sheep (Ovis aries). Appl Anim Ethol 1977;3(2):127-135. [ Links ]
6. Shillito-Walser EE, Hague P, Walters E. Vocal recognition of recorded lambs voices by ewes of three breeds of sheep. Behaviour 1982;78(3-4):261-272. [ Links ]
7. Terrazas A, Ferreira G, Lévy F, Nowak R, Serafin N, Orgeur P et al. Do ewes recognize their lambs within the first day postpartum without the help of olfactory cues? Behav Processes 1999;47(1):19-29. [ Links ]
8. Ferreira G, Terrazas A, Poindron P, Nowak R, Orgeur P, Lévy F. Learning of olfactory cues is not necessary for early lamb recognition by the mother. Physiol Behav 2000;69(4-5):405-412. [ Links ]
9. Keller M, Meurisse M, Poindron P, Nowak R, Ferreira G, Shayit M et al. Maternal experience influences the establishment of visual/auditory, but not olfactory recognition of the newborn lamb by ewes at parturition. Dev Psychobiol 2003;43(3):167-176. [ Links ]
10. Poindron P, Gilling G, Hernández H, Serafín N, Terrazas A. Early recognition of newborn goat kids by their mother: I. Nonolfactory discrimination. Dev Psychobiol 2003;43(2):82-89. [ Links ]
11. Nowak R, Poindron P, Le Neindre P, Putu IG. Ability of 12-hour-old merino and crossbred lambs to recognise their mothers. Appl Anim Behav Sci 1987;17(3-4):263-271. [ Links ]
12. Nowak R. Senses involved in the discrimination of Merino ewes at close contact and from a distance by their newborn lambs. Anim Behav 1991;42(3):357-366. [ Links ]
13. Nowak R, Lindsay DR. Discrimination of merino ewes by their newborn lambs:important for survival? Appl Anim Behav Sci 1992;34(1):61-74. [ Links ]
14. Terrazas A, Nowak R, Serafin N, Ferreira G, Levy F, Poindron P. Twenty-four-hour-old lambs rely more on maternal behavior than on the learning of individual characteristics to discriminate between their own and an alien mother. Dev Psychobiol 2002;40(4):408-418. [ Links ]
15. Sebe F, Nowak R, Poindron P, Aubin T. Establishment of vocal communication and discrimination between ewes and their lamb in the first two days after parturition. Dev Psychobiol 2007;49(4):375-386. [ Links ]
16. Searby A, Jouventin P. Mother-lamb acoustic recognition in sheep: a frequency coding. Proc R Soc Lond Ser B-Biol Sci 2003;270(1526):1765-1771. [ Links ]
17. Sebe F, Aubin T, Boue A, Poindron P. Mother-young vocal communication and acoustic recognition promote preferential nursing in sheep. J Exp Biol 2008;211(Pt 22):3554-3562. [ Links ]
18. Terrazas A, Robledo V, Serafin N, Soto R, Hernandez H, Poindron P. Differential effects of undernutrition during pregnancy on the behaviour of does and their kids at parturition and on the establishment of mutual recognition. Animal 2009;3(2):294-306. [ Links ]
19. Robledo-Murua VMA. Estudio de los efectos de la desnutrición durante la gestación sobre las relaciones madre-cría en cabras. Facultad de Estudios Superiores Cuautitlán, Unidad de Posgrado. México: Universidad Nacional Autónoma de México, 2005. [ Links ]
20. Dwyer CM, Lawrence AB, Bishop SC, Lewis M. Ewe-lamb bonding behaviours at birth are affected by maternal undernutrition in pregnancy. Br J Nutr 2003;89(1):123-136. [ Links ]
21. Putu IG, Poindron P, Lindsay DR. Early disturbance of merino ewes from the birth site increases lamb separations and mortality. Proc Aust Soc Anim Prod 1988;17:298-301. [ Links ]
22. Lindsay DR, Nowak R, Putu IG, McNeill D. Behavioural interactions between the ewe and her young at parturition: a vital step for the lamb. In: Oldham CM, Martin GB, Purvis IW, editors. Reproductive physiology of merino sheep: Concepts and consequences. Perth, Australia: The University of Western Australia; 1990:191-205. [ Links ]
23. NRC. Nutrient requirements of sheep. 6 ed. Washington, DC: The National Academy Press; 1985. [ Links ]
24. Ramírez M, Soto R, Poindron P, Álvarez L, Valencia JJ, González F et al. Comportamiento maternal alrededor del parto y reconocimiento madre cría en ovinos pelibuey. Vet Méx 2011;42(1):27-46. [ Links ]
25. Terrazas A, Serafín N, Hernández H, Nowak R, Poindron P. Early recognition of newborn goat kids by their mother: II. Auditory recognition and evidence of an individual acoustic signature in the neonate. Dev Psychobiol 2003;43(4):311-320. [ Links ]
26. Caldeira RM, Belo AT, Santos CC, Vazques MI, Portugal AV. The effect of long-term feed restriction and over-nutrition on body condition score, blood metabolites and hormonal profiles in ewes. Small Ruminant Res 2007;68:242-255. [ Links ]
27. Erhard HW, Boissy A, Rae MT, Rhind SM. Effects of prenatal undernutrition on emotional reactivity and cognitive flexibility in adult sheep. Behav Brain Res 2004;151(1-2):25-35. [ Links ]

