










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de ciencias pecuarias
versión On-line ISSN 2448-6698versión impresa ISSN 2007-1124
Rev. mex. de cienc. pecuarias vol.3 no.1 Mérida ene./mar. 2012
Artículos
Parámetros y tendencias genéticas del peso al destete y a los 18 meses de edad en ganado Cebú bermejo de Cuba
Parameters and genetic trends for weight at weaning and 18 months of age in red Zebu cattle in Cuba
Ariel Guillén Trujilloª, Danilo Guerra Iglesiasb†, Narciso Ávila Serranoc, Alejandro Palacios Espinosaª, Ricardo Ortega Pérezª, José Luis Espinoza Villavicencioª
ª Departamento de Zootecnia, Universidad Autónoma de Baja California Sur, Carretera al Sur, km. 5.5, CP 23080. La Paz, B.C.S., México. jlvilla@uabcs.mx. Correspondencia al último autor.
b Departamento de Genética, Centro de Investigaciones para el Mejoramiento Animal de la Ganadería Tropical, La Habana, Cuba.
c Universidad del Mar, Oaxaca, México.
† Fallecido en Junio de 2011.
Recibido el 20 de enero de 2011.
Aceptado el 30 de junio de 2011.
Resumen
El objetivo fue estimar la heredabilidad directa y materna del peso al destete a los 7 meses (PD), y peso final a los 18 meses (PF) y sus valores y tendencias genéticas en bovinos Cebú bermejo de Cuba. Se utilizó un modelo animal bicarácter en 5,120 registros de PD y 1,134 de PF. Los animales nacieron entre 1982 y 2008 en tres hatos. El modelo para el PD incluyó los efectos del grupo de contemporáneas (hato–año–estación de nacimiento–sexo, edad al destete como covariable lineal y edad de la madre como lineal y cuadrática), efecto aleatorio del animal, aleatorio materno, del ambiente permanente y el error. El modelo para PF incluyó el grupo de contemporáneas (hato–año–estación de nacimiento), edad al peso final como covariable lineal, efecto aleatorio del animal, efecto del ambiente permanente y el error. Las tendencias genéticas se estimaron mediante regresión lineal del valor genético aditivo promedio sobre los años de nacimiento, de 1982 a 2007. La heredabilidad del PD fue 0.11±0.04 y 0.03±0.02 (efectos directos y maternos, respectivamente) con una correlación genética de –0.15±0.24 entre dichos efectos. La heredabilidad del PF fue 0.28±0.10. Las tendencias genéticas resultaron positivas y significativas para el efecto directo de PD (0.102 kg) y PF (0.244 kg). Se concluye que la heredabilidad de los efectos directos y maternos del PD fueron bajas, y el PF tiene un índice de herencia moderado que puede favorecer la respuesta a la selección en los programas de mejoramiento genético.
Palabras clave: Heredabilidad, Bovinos en trópico, Valor genético, Peso final.
Abstract
The objective was to estimate direct and maternal heritability of weaning weight at 7 mo of age (WW), and the final weight at 18 mo (FW) and breeding values and genetic trends for WW and FW in Zebu bermejo cattle of Cuba. A bi–character animal model was used in 5,120 records of WW and 1,134 of FW. The animals were born between 1982 and 2008 in three herds. The statistical model for WW included the effects of the group of contemporaries (herd–year–birth season–sex, age at weaning as linear co–variable and age of the mother as linear and quadratic covariable), random effect of the animal, random maternal effect, permanent environmental effect and the error. The model for FW included the group of contemporaries (herd–year–birth season) age at final weight as linear co–variable, random effect of the animal, permanent environmental effect and the error. Genetic tendencies were estimated by linear regression of the average additive genetic value on the birth years from 1982 to 2007. The heritability estimated for the WW were 0.11±0.04 and 0.03±0.02 (direct and maternal effects, respectively), with a genetic correlation of –0.15±0.24 between these effects. The heritability for the FW was 0.28±0.10. Genetic tendencies were positive and significant for direct effects of PD (0.102 kg) and PF (0.244 kg). It is concluded that the heritability of the direct and maternal effects of the WW were low and that the FW has a moderate heritability that can favor the response to selection in breeding programs.
Key words: Heritability, Cattle in the tropics, Breeding value, Final weight.
INTRODUCCIÓN
El ganado cebú en Cuba, predominantemente Brahman, representa la mayor proporción de bovinos productores de carne, debido a sus características de adaptabilidad y producción bajo condiciones tropicales extensivas. El país cuenta con 12,000 vacas, aproximadamente, las cuales son registradas en diferentes establecimientos(1).
Desde hace casi 40 años se estableció en Cuba el programa de mejora genética para el ganado Cebú. Durante la década de los 80s se empezaron a realizar las pruebas de comportamiento y de progenie en pastoreo, como parte fundamental del programa, aunque la selección se efectuaba por medio de un índice fenotípico. A partir del año 2000 las evaluaciones se realizan utilizando un modelo animal.
El ganado Cebú de Cuba ha desarrollado de manera independiente tres variantes en el color del pelo, (blanco, bermejo o rojo y sardo). Los trabajos encaminados a la estimación de parámetros genéticos y tendencias genéticas se han realizado en el Cebú blanco(2); sin embargo, no aparecen reportes en la literatura relacionados con el mejoramiento genético del Cebú bermejo cubano.
El peso al destete a los 7 meses (PD) y el peso final (PF) son dos características importantes en el mejoramiento genético del ganado bovino de carne en Cuba. El PD es un rasgo influido por los efectos maternos; por tanto, para lograr un progreso genético se debe tener en cuenta los componentes genéticos directos y maternos, especialmente si hay una relación genética antagónica entre ellos(3). Cuando los efectos maternos son significativos y no se consideran en los modelos para estimar parámetros genéticos, la estimación puede estar sesgada y reducir la eficiencia de la selección (4) debido a una sobreestimación de la heredabilidad directa(5). Para los rasgos de crecimiento, la mayoría de las estimaciones de heredabilidad directa, con los efectos directo y materno incluidos en el modelo, han variado entre 0.20 y 0.30(6,7). Cuando los efectos maternos son ignorados, la heredabilidad directa para la ganancia de peso antes del destete(8) y el PD(9) pueden alcanzar valores de 0.73 y 0.45, respectivamente. Para tales rasgos, Dickerson(10) presentó un modelo genético cuantitativo general en el que una característica es la suma de un efecto directo debido al individuo en que se mide el fenotipo, y un efecto materno debido a su madre. Este modelo se usa comúnmente en el mejoramiento genético de animales(11).
En Cuba el destete del ganado Cebú se hace a los 7 meses de edad y se selecciona el 25 % de los terneros machos, aproximadamente para las pruebas de comportamiento hasta los 18 meses de edad. Las pruebas se realizan en sistemas de producción extensivos basados en pastos y sales minerales. Esta selección puede disminuir la variabilidad en el peso a los 18 meses cuando los análisis que se hacen incluyen un solo rasgo y, por tanto, se supone una correlación genética de cero entre el peso a los 7 y 18 meses de edad(3). Sin embargo, también se plantea(12) que cuando los datos provienen de poblaciones donde ha habido selección de caracteres correlacionados genéticamente, los análisis univariados pueden estar sesgados, mientras que con análisis multivariados se puede reducir el sesgo. Por los motivos anteriores, el objetivo de este trabajo fue estimar los parámetros genéticos del peso al destete y del peso final en la prueba de comportamiento utilizando un modelo animal bicaracter y monitorear el trabajo de selección realizado, por medio del cálculo de las tendencias genéticas.
MATERIALES Y MÉTODOS
Para el desarrollo del estudio se utilizaron 5,120 registros del PD y 1,134 registros del PF de los animales que concluyeron la prueba de comportamiento. Los datos pertenecen a bovinos Cebú bermejo de Cuba que nacieron entre 1982 y 2008 en tres hatos ubicados en las provincias de Cienfuegos, Camagüey y Granma de la República de Cuba, situada en la entrada del Golfo de México, entre los 20° a 23° N y 74° a 85° O. Según la clasificación de Köppen (modificada), en la mayor parte de Cuba el clima predominante es del tipo cálido tropical. Hay dos estaciones claramente definidas, la seca (invierno) de noviembre a abril, y la lluviosa (verano) de mayo a octubre. La temperatura media anual es 25 °C, la media en invierno es 20 °C y en verano 26 a 27 °C. La humedad relativa promedio es 80 %. La precipitación media anual es 1,200 mm, alrededor de 30 % ocurre en el período invernal y 70 % en el verano(13).
Los becerros fueron producto del programa de inseminación artificial con observación del estro natural durante todo el año. Fueron destetados a los 7 meses de edad y los machos, seleccionados fenotípicamente, se sometieron a la prueba de comportamiento. Para esto se conformaron dos grupos, animales de 7 a 12 y de 13 a 18 meses hasta concluir la prueba. El sistema de alimentación se basó principalmente en pastoreo en zacate estrella (Cynodon nlemfuensis) y zacate Guinea (Panicum máximum). Los bovinos dispusieron de sales minerales a libre demanda. Se aplicaron vacunas contra leptospira, septicemi a hemorrágica y carbunco, así como la vacuna recombinante contra la garrapata Boophilus microplus cada tres meses, a partir del mes de edad.
En la edición de los datos se eliminaron aquellos grupos de contemporáneas (hato–año–estación de nacimiento) con menos de cinco observaciones. Se eliminaron los individuos con madre desconocida y todos aquellos a los que les faltara la información relacionada con la fecha de nacimiento, edad de la madre, hato, edad y PD. De un total de 7,666 registros iniciales, se utilizaron 5,120 registros para el PD y 1,134 registros para PF. La estructura de la base de datos utilizada se presenta en el Cuadro 1.
El modelo estadístico para PD y PF incluyó los efectos fijos de hato (tres hatos), año (1982–2008) y cuatrimestre de nacimiento (enero–abril, mayo–agosto y septiembre–diciembre), agrupados en grupos de contemporáneas.
Los rasgos analizados fueron el PD y PF. El programa utilizado fue el ASReml(14). El modelo estadístico para el análisis bivariado (PD y PF) se expresa de manera matricial de la siguiente forma:

Donde yPD ( yPF) es el vector de las observaciones para PD(PF), bPD (bPF) es el vector de soluciones para los efectos fijos de los grupos de contemporáneas, adPD (adPF) es el vector de los efectos genéticos aditivos para PD(PF), amPD es el vector de los efectos genéticos maternos de PD, acPD (acPF) es el vector de los efectos del ambiente permanente para PD(PF), y ePD(ePF) es el vector de los efectos residuales para PD(PF) y X PD, X PF, ZdPD, ZdPF, ZmPD, ZcPD, y ZcPF son las matrices de incidencia conocidas.
Se supuso que todos los efectos aleatorios en el modelo tenían media cero. La estructura de (co)varianzas asumida fue la siguiente:

Donde σ2d(PD), σ2d(PF), σ2c(PD), σ2c(PF), σ2e(PD), y σ2e(PF) son las varianzas para los efectos aditivos directos de PD y PF y maternos de PD, del ambiente permanente para PD y PF y las varianzas residuales para PD y PF, respectivamente. De la misma forma, σ2d(PD),d(PF), es la covarianza entre los efectos aditivos de PD y PF, σ2d(PD),m(PD), es la covarianza entre los efectos directos y maternos de PD, σ2d(PD),m(PF), es la covarianza entre los efectos directos y maternos de PD y PF y por último, σ2e(PD),e(PF), es la covarianza de los efectos residuales entre PD y PF.
Las tendencias genéticas se estimaron mediante regresión lineal del valor genético aditivo promedio sobre los años de nacimiento, de 1982 a 2008. Todos los hatos se encontraban interconectados entre sí por al menos un semental en común y al menos por dos sementales en común durante los años de estudio. La descendencia de todos los hatos estuvo interconectada por al menos un semental.
RESULTADOS
En el Cuadro 1 se presentan las características principales (estadísticos descriptivos) de la muestra de animales estudiada. Las medias generales de pesos aumentaron de 159.7 ± 26.7 kg (PD) a 327.7 ± 52.3 kg (PF). Las desviaciones estándar mostraron tendenci as similares a las medias, aumentando con la edad. Los coeficientes de variación de los pesos permanecieron constantes entre el destete y el final de las pruebas de comportamiento. En el Cuadro 2 se muestran los componentes de (co)varianza y los parámetros genéticos del análisis bivariado. Los estimados de la varianza genética aditiva directa, así como la vaianza fenotípica aumentaron con la edad. La h2 estimada para los efectos directos aumentó entre el PD (0.11 ± 0.04) y el PF (0.28 ± 0.10). La h2 para los efectos maternos en el PD fue de 0.03 ± 0.02. La correlación genética entre los efectos directos y maternos del PD fue baja y negativa con un error estándar superior al parámetro estimado (–0.1 5 ± 0. 24). La varianza del ambiente permanente y la proporción del efecto del ambiente permanente con respecto a la varianza fenotípica dismunuyó entre el PD y el PF.

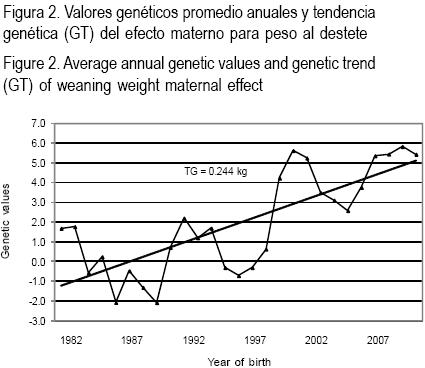
Las tendencias genéticas anuales para los efectos directos y maternos del PD y el efecto directo del PF se presentan en las Figuras 1, 2 y 3 donde se aprecian valores de 0.102, 0.013 y 0.244 kg, respectivamente.


DISCUSIÓN
Los PD registrados en el presente estudio fueron inferiores a los observados en ganado Brahman de Venezuela (163.4 kg)(15) y en Brahman de Sudáfrica (212.2 kg)(16). Resultados inferiores fueron encontrados en otro estudio con ganado Cebú de Venezuela (157.5 kg )(17) y en Cebú cubano (150.7 kg)(3).
El PF que se registró en el presente trabajo fue inferior al reportado en Brahman de Sudáfrica (360.8 kg)(16). Sin embargo, fue superior a los valores mencionados en Brahman blanco de Venezuela (297.4 kg)(15) y de Cuba (326.5 kg)(3).
La heredabilidad para el efecto directo del PD es inferior a los promedios publicados en ganado de carne en países tropicales(18). Ríos(19) en una revisión sobre el tema hace referencia a un valor promedio no ponderado de 0.27 para la h2 del efecto directo del PD para todas las razas de bovinos de carne y de 0.26 en Brahman y Nelore. En general, los valores descritos en la literatura para las razas cebuinas promedian 0.30(20). Recientemente se reportaron valores de directa para el peso de becerros Tabapúa de Brasil de 0.28 y 0.31 a los 120 y 240 días de edad, respectivamente(21). Algunas de las discrepancias con respecto al presente estudio pudieron deberse a que, en los estudios de Mercadante et al(18), y Koots et al(22), los modelos usados no incluyeron el efecto genético materno y, como ha sido señalado, cuando los rasgos son influidos por los efectos directo y materno, la inclusión en el modelo animal del efecto directo únicamente, puede dar lugar a una sobreestimación de la heredabilidad del mismo(5). Otros autores reportan valores de heredabilidad para PD similares (0.08 a 0.10) a los del presente trabajo, estimados con un modelo multirracial que incluyó el efecto genético materno(23).
El estimado de la varianza aditiva se incrementó más que la varianza fenotípica entre el PD y el PF, resultando en un aumento en la estimación de la heredabilidad directa del PF. Lo anterior podría estar relacionado con una posible correlación genética aditiva entre el PD y PF. En general, la heredabilidad directa estimada para el PF y la varianza genética aditiva directa sugieren que en la población estudiada existe suficiente variabilidad genética para el mejoramiento de dicho rasgo con base en la selección de los animales superiores. La heredabilidad directa del PF se incrementó con respecto al PD. Esta tendencia es similar, aunque con heredabilidades de menor magnitud a las descritas en bovinos Tabapua(21) y Nelore(24). La heredabilidad para el PF fue superior a los valores referidos por otros investigadores quienes encontraron estimados de 0.13(15), 0.12(25) y 0.25(3). Un valor más elevado (0.31) fue constatado por Giannotti et al(26).
La heredabilidad estimada para los efectos maternos del PD fue baja, haciendo poco importante la respuesta a la selección para dicho rasgo en esta población de ganado Cebú de Cuba. De acuerdo con Elzo et al(23) las bajas heredabilidades directa y materna del PD indican que las condiciones medioambientales predestete son restrictivas tanto para la madre (producción de leche) como para las crías (crecimiento propio). Estimados bajos de heredabilidad para el PD directo y materno fueron también reportadas por en un hato de ganado Romosinuano–Cebú en Costa Rica(27). Los autores consideran que, bajo el sistema de alimentación predestete basado en praderas con acceso solamente a sales minerales, tanto las madres como los terneros están en condiciones de alimentación insuficientes para que puedan expresar sus potenciales genéticos maternos y directos, respectivamente. Con alimentación predestete con suplementación de proteína y energía, las heredabilidades del PD directo y materno en ganado Angus–Brahman en Florida fueron de 0.25 a 0.29(28).
Albuquerque y Meyer(29) reportan que la heredabilidad de los efectos maternos se incrementó de 0.01 en el peso al nacimiento hasta 0.08 en el PD a los 210 días. La heredabilidad materna del PD en el presente estudio fue inferior a lo mencionado en ganado Nelore (0.08)(30,20,31), así como en bovinos Tabapúa (0.09)(32). Valores más altos (0.17) han sido señalados en ganado Nelore(33) y en Boran Africano(34) (0.27 y 0.31, respectivamente).
La correlación genética entre los efectos directo y materno para el PD fue negativa (–0.15) y de amplia variabilidad. Valores negativos, aunque de diferentes magnitudes (–0.07 y –0.10) fueron estimados también en Cebú blanco de Cuba(3) y en bovinos Asturianos de los Valles(35). Es evidente la existencia de un antagonismo entre los efectos directos y maternos en caracteres como peso al destete en el vacuno de carne(36), lo que supone una restricción en la mejora genética de ese tipo de rasgos. Lo anterior puede deberse a que en el caso de datos de campo, donde se pierde la conexión entre la identificación del animal cuando se desteta y cuando es madre, esta correlación tiende a ser negativa en dependencia del porcentaje de pérdida de información (37).
La literatura internacional presenta grandes diferencias en las estimaciones de la correlación genética entre los efectos directo y materno para el PD. Por ejemplo, en ganado Brangus de Brasil se reporta una correlación de 0.55(38). Las discrepancias se atribuyen a distintas causas, como el uso de muestras de datos de campo o estaciones experimentales y pobre estructura poblacional (37), entre otras. En los sistemas de producción bovina tradicionales, principalmente en los países en desarrollo, a veces es difícil identificar a los animal es y registrar de manera correcta su comportamiento y genealogía, lo que afecta la cantidad y calidad de los datos. Una de las consecuencias puede ser el uso de pedigrís incompletos, que resulta en el uso de una matriz de relaciones menos completa en el modelo animal; además, el escaso intercambio de padres entre los hatos limita el flujo de genes y causa una falta de conectividad genética parcial o total(5). La influencia de la desconectividad genética sobre los modelos animal con efectos directos se observa en la precisión de las estimaciones, principalmente(39).
El valor estimado para la varianza del ambiente permanente como proporción de la varianza fenotípica del PD fue inferior al promedio (0.14) publicado por Ríos(19), calculado sobre 171 estimadores de distintas referencias bibliográficas. Un valor similar al del presente trabajo fue estimado por Lee y Pollac(40) en ganado Simmental. La varianza del ambiente permanente se redujo entre el PD y el PF. Esta tendencia es similar a la descrita en una raza sintética de bovinos para carne de Australia(41). En contraste, otros investigadores(32) observaron un incremento de la varianza del ambiente permanente en función de la edad. De igual manera, Sousa et al(21) reportan que los estimados de varianza del ambiente permanente obtenidos mediante regresión aleatoria presentaron una tendencia de crecimiento entre el nacimiento (0.01) hasta los 160 días de edad (0.29) con una disminución pequeña a los 440 días (0.23) y, a partir de esa edad, permanecieron constantes (0.24).
De manera general los análisis bivariados para PD y PF han dado estimados de superiores para estos caracteres que cuando se realizan univariados(3,42). Schaeffer(43) plantea que en los análisis de rasgos múltiples, todos los rasgos se benefician en alguna medida, y que estos son más útiles cuando la diferencia entre la correlación genética y la residual es superior a 0.5. En el presente trabajo la correlación genética entre PD y PF fue de 0.98 ± 0,10 y la residual de 0.52 ± 0.05, con una diferencia entre ellas de 0.46 muy cerca de lo planteado por Schaeffer(43) quien señala también que pueden existir ventajas cuando hay un rasgo de baja heredabilidad y pueden ocurrir sesgos debidos a la selección, como es el caso de la presente muestra, donde solamente el 25 % de los terneros machos que nacen son llevados a las pruebas de comportamiento.
Las tendencias para los efectos directos del PD y PF representaron un 0.06 y 0.07 % de mejora anual. Esas tendencias indican que el cambio genético anual es escaso, y que se debe de dar más atención a la selección de los individuos utilizados para la reproducción. En Brasil existen al menos cuatro grupos de investigación en mejoramiento genético para la raza Nelore que publican los resúmenes de toros donde las tendencias genéticas varían entre 0.15 y 0.61 % para los pesos hasta el destete y de 0.15 a 0.77 % para los pesos posdestete(44). Por otro lado, en un hato de ganado Brahman de Venezuela sometido a selección, se encontraron tendencias del PD y peso a los 18 meses, de 0.142 y 0.263 kg, respectivamente(15). En ganado Cebú cubano refieren una tendencia de 0.359 kg, para el peso a los 18 meses de edad(45). Las tendencias genéticas bajas se pueden explicar por la influencia de los efectos ambientales o por una selección direccional baja(46). Las tendencias genéticas observadas en el presente trabajo se pueden deber a que la selección para el PD y PF se hace utilizando índices fenotípicos expresados como porcentaje de la media de los contemporáneos y que se confiere un mayor peso en la selección de características del tipo y conformación.
CONCLUSIONES E IMPLICACIONES
De acuerdo con los resultados de este trabajo se concluye que los bajos valores de heredabilidadad estimados para los efectos directo y materno del peso al destete son una limitante para el mejoramiento genético de estos rasgos. Sin embargo, el valor estimado para la heredabilidad del peso a los 18 meses de edad ofrece buenas posibilidades de respuesta a la selección.
LITERATURA CITADA
1. Espinoza JL, Palacios A, de Luna R, Ávila N, Guerra D, González–Peña D, Rodríguez F, Mellado M. Componentes de (co)varianza para caracteres de crecimiento y reproducción en ganado Cebú en Cuba. Arch Zootec 2007;56:919–927. [ Links ]
2. Menéndez A, Guerra D, Planas T, Ramos F. Factors affecting the growth of young bulls of the Zebu breeds in the performance test in grazing conditions. Cuban J Agric Sci 2006;40:371–377. [ Links ]
3. Espinoza–Villavicencio JL, Palacios–Espinosa A, Guerra–Iglesias D, González–Peña D, Ortega–Pérez R, Rodríguez–Almeida F. Comparación de dos modelos para la estimación de parámetros y valores genéticos del peso en ganado Cebú. Agrociencia 2008;42(1):29–36. [ Links ]
4. Azadi Fozi M, Van der Werf JHJ, Swan AA. The importance of accounting for maternal genetic effects in Australian fine–wool Merino breeding. Austr J Agric Res 2005;56(8):789–796. [ Links ]
5. Clément V, Bibé B, Verrier E, Elsen J, Manfredi E, Bouix J, Hanock E. Simulation analysis to test the influence of model adequacy and data structure on the estimation of genetic parameters for traits with direct and maternal effects. Genet Sel Evol 2001;33(4):369–395. [ Links ]
6. Ménissier F. Comments on optimisation of cattle breeding scheme: beef breeds for suckling herds. A review. Ann Génét Sél Anim 1976;8:71–87. [ Links ]
7. Waldron DF, Morris CA, Baker RL, Johnson DL. Maternal effects for growth traits in beef cattle. Livest Prod Sci 1993;34(1–2):57–70. [ Links ]
8. Jurado JJ, Alonso A, Alenda R. Selection response for growth in a Spanish Merino flock. J Anim Sci 1994;72(6):1433–1440. [ Links ]
9. Brash LD, Fogarty NM, Gilmour AR. Genetic parameters for Australian maternal and dual–purpose meatsheep breeds. III. Liveweight, fat depth and wool production in Coopworth sheep. Austr J Agric Res 1994;45(2):481–486. [ Links ]
10. Dickerson GE. Composition of hog carcasses as influenced by heritable differences in rate and economy of gain. Iowa Agric Exp Stat Res Bull 1947;354:489–524. [ Links ]
11. Bjjma P. Estimating maternal genetic effects in livestock. J Anim Sci 2006;84(4):800–806. [ Links ]
12. Thompson R, Meyer K. A review of theoretical aspects in the estimation of breeding values for multi–trait selection. Livest Prod Sci 1986;15(4):299–313. [ Links ]
13. Hernández I, Milera M, Simón L, Hernández D, Iglesias J, Lamela L, Toral O, Matías C, Geraldine F. Avances en las investigaciones en sistemas silvopastoriles en Cuba. En: Sánchez MD, Rosales M editores. Agroforestería para la producción animal en Latinoamérica, Roma 1999;143:89–106. [ Links ]
14. Gilmour AR, BR Cullis, SJ Welham, R Thompson. ASREML Reference Manual. Australia. NSW, Agriculture Biometric. 2000. [ Links ]
15. Plasse D, Verde O, Fossi H, Romero R, Hoogestejjn R, Bastidas P, Bastardo J. (Co)variance components, genetic parameters and annual trends for calf weights in a pedigree Brahman herd under selection for three decades. J Anim Breed Genet 2002;119(3):141–153. [ Links ]
16. Pico BA. Estimation of genetic parameters for growth traits in South African Brahman cattle [Master thesis]. South Africa: University of the Free State; 2004. [ Links ]
17. Plasse D, Verde O, Arango J, Camaripano L, Fossi H, Romero R, Rodríguez CM, Rumbos JL. (Co)variance components, genetic parameters and annual trends for calf weights in a Brahman herd kept on floodable savanna. Genet Mol Res 2002;1(4):282–297. [ Links ]
18. Mercadante MEZ, Lobo RB, de los Reyes A. Parámetros genéticos para características de crecimiento en cebuinos de carne. Arch Latinoam Prod Anim 1995;3:45–89. [ Links ]
19. Ríos A. Estimadores de parámetros genéticos para características de crecimiento predestete de bovinos. Revisión. Tec Pecu Mex 2008;46(1):37–67. [ Links ]
20. Cabrera ME, Garnero AV, Lôbo RB, Gunski RJ. Efecto de la incorporación de la covarianza genética directa–materna en el análisis de características de crecimiento en la raza Nelore. Livest Res Rural Dev 2001;13. [ Links ]
21. Sousa CS, Oliveira SMP, Albuquerque LG, Boligon AA, Filho RM. Estimação de funções de covariância para características de crescimento da raça Tabapuã utilizando modelos de regressão aleatória. R Bras Zootec 2010;39(5):1037–1045. [ Links ]
22. Koots KR, Gibson JP, Wilton JW. Analysis of published genetic parameters estimates for beef production traits. 1. Heritability. Anim Breed Abstr 1994;62:309–388. [ Links ]
23. Elzo MA, Martínez G, González F, Huertas H. Variabilidad y predicciones genéticas aditivas, no aditivas y totales para la producción del ganado de carne en el rebaño multirracial Sanmartinero–Cebú de la Libertad. Rev Corpoica 2001;3(2):51–64. [ Links ]
24. Albuquerque LG, El Faro L. Comparações entre os valores genéticos para características de crescimento de bovinos da raça Nelore preditos com modelos de dimensão finita ou infinita. Rev Bras Zootec 2008;37(2):238–246. [ Links ]
25. Martinez–Gonzalez JC, Castillo–Rodriguez SP, Lucero–Magaña FA, Ortega–Rivas E. Influencias ambientales y heredabilidad para características de crecimiento en ganado Sardo Negro en México. Zootecnia Trop 2007;25(1):1–7. [ Links ]
26. Giannotti JG, Packer IU, Mercadante MEZ. Meta–análise para estimativas de herdabilidade para características de crescimento em bovinos de corte. Rev Bras Zootec 2005;34(4):1173–1180. [ Links ]
27. Elzo MA, Manrique C, Ossa G, Acosta O. Additive and nonadditive genetic variability for growth traits in the Turipaná Romosinuano–Zebu multibreed herd. J Anim Sci 1998;76(6):1539–1549. [ Links ]
28. Elzo MA, Wakeman DL. Covariance components and predictions for additive and nonadditive preweaning growth genetic effects in an Angus–Brahman multibreed herd. J Anim Sci 1998;76(5):1290–1302. [ Links ]
29. Albuquerque LG, Meyer K. Estim ates of direct and maternal genetic effects for weights from birth to 600 days of age in Nelore cattle. J Anim Breed Genet 2001;118(2):83–92. [ Links ]
30. Mercadante MEZ, Lôbo RB, Oliveira HN. Estimativas de (co)variância entre características de reprodução e de crescimento em fêmeas de um rebanho N el ore. Rev Bras Zootec 2000;29(4):997–1004. [ Links ]
31. Nobre PRC, Misztal I, Tsuruta S, Bertrand JK, Silva LOC, Lopes PS. Analyses of grwth curves of Nellore cattle by multiple–trait and random regression models. J Anim Sci 2003;81(4):918–926. [ Links ]
32. Dias TL, Albuquerque LG, Tonhati H, Teixeira RA. Estimação de parâmetros genéticos para peso do nascimento aos 550 dias de idade para animais da raça Tabapuã utilizando–se modelos de regressão aleatória. Rev Bras Zootec 2006;35(5):1915–1925. [ Links ]
33. Bertazzo RP, Fonseca RT, de Moraes T, Garcia I, Eler J, Sterman J, Gomes A, de Andrade I. Parâmetros genéticos de longevidade e produtividade de fêmeas da raça Nelore. Rev Bras Zootec 2004;33(5):1118–1127. [ Links ]
34. Wasike CB, Ilatsia ED, Ojango JMK, Kahi AK. Genetic parameters for weaning weight of Kenyan Boran cattle accounting for direct–maternal genetic covariances. S Afr J Anim Sci 2006;36(4):275–281. [ Links ]
35. Gutiérrez JP, Fernández I, Alvarez I, Royo LJ, Goyache F. Sire x contemporary group interactions for Barth weight and preweaning growth traits in the Asturiana de los Valles beef cattle breed. Livest Sci 2006;99(1):61–68. [ Links ]
36. Robinson DL. Estimation and interpretation of direct and maternal genetic parameters for weights of Australian Angus cattle. Livest Prod Sci 1996;45(1):1–11. [ Links ]
37. Heydarpour M, Schaeffer LR, Yazdi MH. Influence of population structure on estimates of direct and maternal parameters. J Anim Breed Genet 2008;125(2):89–99. [ Links ]
38. Lopes JS, Rorato PRN, Weber T, Dornelles MA, Comin JG, de Araújo RO. Parâmetros genéticos e tendências genética e fenotípica para características de crescimento em uma população da raça Brangus. Rev Bras Zootec 2009;38(4):662–669. [ Links ]
39. Diaz C, Carabano MJ, Hernandez D. Connectedness in genetic parameters estimation and BV prediction [abstract]. Proc 46th Ann Meetting Eur Assoc Anim Prod, Prague, Czech Republic, 1995:8. [ Links ]
40. Lee C, Pollak EJ. Relationship between sire x year interactions and direct maternal genetic correlation for weaning weight of Simmental cattle. J Anim Sci 1997;75(1):68–75. [ Links ]
41. Meyer K. Estimates of direct and maternal covariance functions for growth of Australian beef calves from birth to weaning. Genet Sel Evol 2001;33(5):487–514. [ Links ]
42. Bennett GL, Gregory KE. Genetics (co)variances among birth weight, 200–day weight and postweaning gain in composites and parental breeds of beef cattle. J Anim Sci 1996;74(11):2596–2611. [ Links ]
43. Schaeffer LR. Sire and cow evaluation under multiple trait models. J Dairy Sci 1984;67(7):1567–1580. [ Links ]
44. Albuquerque LG, Mercadante MEZ, Eler JP. Recent studies on the genetic basis for selection of Bos indicus for beef production. Proc 8th World Congr Genet Appl Livest Prod, Belo Horizonte, Brazil, 2006:1–7. [ Links ]
45. Guerra D, Rodriguez M, Ramos F, Planas T, Ortiz J, Torres JA, Falcón R. Evaluación genetic de las razas de carne en Cuba [resumen]. II Congreso Internacional de Mejora Animal, La Habana, CU 2002:4. [ Links ]
46. Razook AG, Figueiredo LA Neto, LMB, Trovo JBF, Packer IU, Pacola LJ, Candido JG. Intensidades de seleção e respostas direta e correlacionadas em 10 anos de progênies de bovinos das raças Nelore e Guzerá selecionadas para peso pós–desmame. Boletim da Indústria Animal 1993;50:147–163. [ Links ]

