










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de ciencias pecuarias
versión On-line ISSN 2448-6698versión impresa ISSN 2007-1124
Rev. mex. de cienc. pecuarias vol.1 no.4 Mérida oct./dic. 2010
Artículos
Eventos reproductivos de vacas con diferente porcentaje de genes Bos taurus en el trópico mexicano
Effects of Bos taurus gene percentage on reproductive performance of crossbred cows in the humid tropics of Mexico
Rufino López Ordaza, Mario Díaz Hernándeza, José Guadalupe García Muñiza, Rafael Núñez Domíngueza, Reyes López Ordazb, Pedro Arturo Martínez Hernándeza
a Departamento de Zootecnia. Universidad Autónoma Chapingo. Km 38.5 Carretera México Texcoco. 56230. Chapingo, México. Tel: 01 595 95 21621. rlopezor@yahoo.com. Correspondencia al primer autor.
b Departamento de Genética y Estadística. FMVZ, UNAM.
Recibido el 4 de enero de 2010
Aceptado para su publicación el 2 de julio de 2010
RESUMEN
El objetivo fue determinar el porcentaje óptimo de genes Bos taurus (BT) para edad al primer parto (EPP), días al primer servicio (DPS), días abiertos (DA) e intervalo entre partos (IEP) de vacas cruzadas. La información analizada se originó de 1,704 lactancias completas de 677 vacas desde 100 % Bos indicus (BI) hasta 100 % BT de un rancho lechero típico mexicano de Tabasco. El análisis fue con un modelo mixto que incluyó los efectos fijos de número de parto de la vaca, el año de parto y la estación de parto de la misma, los efectos aleatorios de semental, vaca anidada en semental, y el efecto racial lineal de porcentaje de genes BT como covariable. Las vacas de cuarto parto tuvieron DPS más cortos que las de segundo y tercero; éstas a su vez fueron diferentes a las de primero. Paralelamente, los animales de segundo ó más partos presentaron DA e IEP más cortos (P<0.05) que las de primero. La EPP fue mínima (31 meses) con 49 % de genes BT; después de dicha proporción el mismo parámetro incrementó hasta 36 meses. Las vacas con 100 % de genes BI presentaron DPS (106 d), DA (166 d) e IEP (14.6 meses) inferiores que las demás cruzadas con BT. En conclusión, las vacas con 50 % de genes BT alcanzan la EPP más temprano y tienen DPS, DA e IEP más prolongados; por lo tanto, dichas vacas representan los genotipos óptimos para la región tropical húmeda.
Palabras clave: Cruzamiento, Ganado bovino tropical, Reproducción.
ABSTRACT
The objective of the present study was to determine the best possible percentage of Bos taurus (BT) genes on age–at–first–calving (AFC), days to first post–partum service (DFPS), days open (DO) and calving interval (CI) in crossbred cows. Data was collected in a typical commercial dairy farm of Tabasco on 1,704 full lactations (270–d) from 677 cows ranging between 100 % Bos taurus (BT) and 100 % Bos indicus (BI). Data analysis was carried out through a mixed model which included the following unchanging effects: calving number, calving year and season and the random effects of sire, cow within sire, with BT gene percentage effect as covariable. Fourth calving cows showed a lower DFPS (P<0.05) than second and third calving animals and these were different to primiparous. Correspondingly, second or more calving cows showed shorter DO and CI (P<0.05) than first calving cows. AFC was shortest in 49 % BT genes, increasing to 36 mo as this percentage increased. Cows having 100 % BI genes showed 106 d DFPS, 166 d DO and 14.6 mo CI, all of them shorter than in crossbred cows. As a conclusion, it can be stated that cows having 50 % BT genes show longer DFPS, DO and CI and shorter AFC, therefore being the best genotype for dairy herds in the humid tropics.
Key words: Crossbreeding, Tropical bovines, Reproductive performance.
INTRODUCCIÓN
La producción de leche en los ambientes tropicales se fundamenta en la capacidad reproductiva de los bovinos cruzados. Dichos animales (Bos taurus X Bos indicus) producen más leche, carne y son superiores para reproducirse comparados con sus razas parentales puras en los ambientes tropicales(1,2). Sin embargo, la existencia de distintos tipos de trópicos y la diversidad genética de los bovinos locales dificultan la aplicación de los sistemas de cruzamientos.
Los problemas más evidentes se relacionan con el mantenimiento de los bovinos F1 (BT X BI), el mantenimiento de las razas parentales puras y la sostenibilidad del equilibrio de la proporción de genes apropiada a las condiciones del ambiente especifico. Otros problemas se relacionan con la expresión de los genes; en algunos ambientes tropicales, los bovinos F1 (BT X BI) exhiben edades al primer parto más temprano, días abiertos e intervalo entre partos más reducidos y menos producción de leche que las vacas con 75 % de genes BT(3); dicho comportamiento no es similar para las vacas del mismo origen de cruzamientos y en ambientes más húmedos. Lo anterior implica la necesidad de generar información de cruzamientos que sincronicen la habilidad de la vaca para producir leche, con comportamientos reproductivos eficientes, y el ambiente apropiado para aprovechar la heterosis del apareamiento de BT X BI.
Con base en lo anterior, el objetivo del presente estudio fue determinar la proporción óptima de genes de Bos taurus (BT) en edad al primer parto (EPP), días al primer servicio posparto (DSP), días abiertos (DA) e intervalo entre partos (IEP) de vacas cruzadas con distintas proporciones de Bos indicus (BI) en un rancho lechero típico del trópico mexicano.
MATERIALES Y MÉTODOS
El estudio se realizó con información de enero de 1991 a diciembre de 2004 con 1,704 lactancias de 270 días de 677 vacas Bos taurus (BT) x Bos inidicus (BI). En dichos animales, la proporción de genes BI fue variable desde 0 hasta 100 % puros; por ejemplo, las vacas con proporciones de 1/8, 1/4, 1/2, 3/8 y 3/4 de genes BT fueron considerados como los términos relativos de 12.5, 25, 50. 62.5, y 75 % de genes BT. En general se tuvieron más de seis grupos raciales; por ejemplo, las vacas de 25, 50 y 75 % de genes BI representaron 268, 326 y 306 lactancias para cada grupo, mientras que las de 12.5, 62.5 y 87.5 de genes BI tuvieron 270, 267 y 269 lactancias completas, respectivamente.
Los genotipos 1/4BT 3/4BI provienen del apareamiento de la raza pura con 1/2BT 1/2BI, mientras que 5/8BT 3/8BI se obtuvieron del apareamiento de la raza pura con 3/4BT 1/4BI. Los genotipos 1/2BT 1/2BI se originaron del apareamiento de las razas puras BT y BI; mientras que los 5/8BT 3/8BI provienen del apareamiento entre 1/2BT 1/2BI con 3/4BT 1/4BI. Los genotipos con genes de cebú fueron Gyr (promedio, 80 %), y Brahman e Indobrasil (20 %); mientras que los genes europeos provienen de Holstein (promedio, 60 %), Simmental (20 %), y el restante Suizo Pardo y Jersey.
La información estudiada se obtuvo de un rancho comercial de Centro, Tabasco. El municipio se localiza a 18°20' N y 93° 15' O. GarcíaW clasificó el clima de la zona como Am(f)w"(i')g, que corresponde a cálido–húmedo con abundantes lluvias en verano. La temperatura media anual de 33.6 °C, y precipitación promedio de 2,237 mm anuales. El mes más cálido es mayo y el más fresco es enero.
La base de la alimentación fue el pastoreo rotacional de gramas tropicales con asignación diaria en franjas con cerco eléctrico. Las especies forrajeras incluyeron al Paspalum spp (camalote o remolino), Homolepsis spp (grama amarga) y Cynodon spp (bermuda). Las vacas en lactancia recibieron forraje y 1.0 kg de concentrado comercial (16.0% PC) durante el ordeño de la mañana y 1.0 kg durante la tarde.
El sistema de ordeño fue mecánico en una sala en paralelo con 16 plazas y pesadores individuales del tipo Waikato™. Las vacas ingresaban a la línea de ordeño dos días después del parto. El ordeño se realizó dos veces al día (0500 y 1330) sin la presencia del ternero y con la ayuda de 0.25 ml de oxitocina (Oxipar). El nivel de producción de 2.0 y 5.0 kg de leche por día fue el criterio principal del secado de las vacas.
El manejo reproductivo del hato consistió de dos empadres controlados, con el uso de inseminación artificial (57 %) y monta natural (43 %). El primer empadre se realiza entre marzo y abril, y el segundo, en junio. Como un objetivo particular del rancho, el productor desea tener animales de aproximadamente el 62.5 % de genes BT; debido a esto, con base en la proporción de genes BT de la madre, se asigna un semental que reúna dichas proporciones.
En la edición de los datos se siguieron varios criterios; se eliminaron los registros de animales con lactancias menores a 270 días, animales que no tuvieron información de una primera lactancia, o que carecían de información del número de lactancias a la que pertenecía cada registro. También se eliminaron las vacas con registros de códigos no utilizables (lactancias con abortos, muerte, venta enfermedades o traumatismos); vacas con menos de 18 meses de edad al primer parto, vacas con errores en los registros (fechas equivocadas de nacimiento, parto o inicio de la lactancia) y registros de IEP fuera del rango de 300 y 730 días.
Con base en las restricciones anteriores, la información estudiada permitió analizar 1,570, 1,704, 1,297 y 1546 registros individuales de edad al primer parto (EPP); días al primer servicio (DPS); días abiertos (DA) e intervalo entre partos (IEP), respectivamente. La información del número de lactancias se agrupó en cuatro categorías, incluyendo animales desde una hasta 10 lactancias. La categoría 1993 incluyó la información de 1991 hasta 1993, la 1997 de 1994 a 1997, la 2001 de 1998 a 2001; mientras que la 2004 incluyó la información correspondiente al 2002 hasta el 2004. La edad al primer parto también se agrupó en cuatro categorías: 1993, 1997,2001 y 2004; la primera contuvo los partos de 1991 a 1993, la segunda de 1994 a 1997, la tercera de 1998 a 2001 y la última consideró los años de 2002 a 2004. Paralelamente, la información relativa a los DA y al IEP fueron ordenadas en dos categorías correspondientes al año de parto: 1997 y 2004; la primera incluyó los partos de 1991 a 1997 y la segunda los de 1998 al 2004. Las épocas de parto y de nacimiento se agruparon en lluvias y seca. La época de lluvias fue de junio a diciembre mientras que la seca fue de enero a mayo.
En el Cuadro 1 se presentan los estadísticos descriptivos de los datos usados en el estudio para EPP, DPS, DA e IEP con sus respectivas fuentes de variación como número de parto, año y época de parto, la interacción año x época (que fue significativa en los análisis preliminares), año de nacimiento y el efecto racial lineal de la covariable porcentaje de genes BT.
La EPP, DPS, DA y el IEP se analizaron con el procedimiento Mixed de SAS(5); el modelo para EPP incluyó los efectos fijos del año de nacimiento de las vacas, el efecto fijo de la época de nacimiento, la interacción de año de nacimiento x época de nacimiento, y el efecto racial lineal de porcentaje de genes BT; mientras que el modelo para DPS, DA e IEP incluyeron los efectos fijos de año de parto de la vaca, estación de parto, número de parto, los efectos aleatorios de semental y de vaca anidada en semental, y el efecto racial lineal de porcentaje de genes como covariable.
Con el propósito de explicar las posibles diferencias debidas a genotipos; el efecto de la heterosis individual de las vacas se adicionó como covariable en los modelos de análisis. La heterosis de la vaca se calculó sumando los productos de la fracción de genes BT del padre por la fracción de genes BI de la madre. Por ejemplo, para el cruzamiento donde el padre es 5/8 BT 3/8 BI y la madre es 3/8 BI 5/8 BT, la heterosis de la vaca se estimó de la siguiente manera:
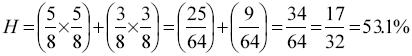
Los resultados preliminares indican que el efecto de la heterosis de la vaca fue no significativo (P>0.05) y por lo tanto no se incluyó en los modelos finales.
El modelo estadístico mixto para edad al primer parto, después de eliminar las interacciones dobles y covariables no significativas (P>0.05) en análisis preliminares, fue el siguiente:

Donde: Yijkl es una observación de la variable respuesta (EPP); μ es la media general; ANi es el efecto fijo del i–ésimo año de nacimiento de la vaca (i=1990, 1991,..., 2000); ENj es el efecto fijo de la j–ésima estación de nacimiento de la vaca (j=lluvias, seca); AN*ENij es el efecto fijo de la interacción de año de nacimiento con estación de nacimiento de la vaca; Sk es el efecto aleatorio del k–ésimo semental (k=1,..., 116); PCTGEU es el efecto racial lineal de porcentaje de genes de razas europeas como covariable; b1 es el coeficiente de regresión lineal asociado con el porcentaje de genes de razas europeas; μ PCTGEU es la media estimada de la covariable racial lineal porcentaje de genes de razas europeas; y eijkl es el error aleatorio.
Se estimaron las medias de cuadrados mínimos para los efectos principales y las interacciones de primer orden significativas (P< 0.05), y se compararon utilizando la prueba de Tukey. Para la covariable PCTGEU se obtuvieron los estimadores de los coeficientes de regresión respectivos. Para calcular el intercepto de la ecuación de regresión de las variables se utilizó la estimación de la media general de cada carácter. Para cada ecuación, se obtuvo la primera derivada con respecto al porcentaje de genes BT y se igualó a cero para obtener un máximo.
Por otro lado, el modelo estadístico mixto para días al primer servicio, días abiertos e intervalo entre partos, después de eliminar las interacciones dobles y covariables no significativas (P>0.05) en análisis preliminares fue el siguiente:

Donde: Y ijklmn es una observación de la variable respuesta; μ es la media general; APi es el efecto fijo del i–ésimo año de parto de la vaca (i=de 1994, 1995,..., 2001); EPj es el efecto fijo de la j–ésima estación de parto de la vaca (j=lluvias, seca); NPk es el efecto fijo del k–ésimo número de parto de la vaca (k=1, 2,..., 5+); Sl es el efecto aleatorio del l–ésimo semental (l=1,...,97); V(S)lm es el efecto aleatorio de la m–ésima vaca anidada en el l–ésimo semental; PCTGEU es el porcentaje de genes de razas europeas como covariable; b1 es el coeficiente de regresión lineal asociado con el porcentaje de genes de razas europeas; μ PCTGEU es la media estimada de la covariable porcentaje de genes de razas europeas; y eijklmn es el error aleatorio.
Se estimaron las medias de cuadrados mínimos para los efectos principales y las interacciones de primer orden significativas (P<0.05), y se compararon utilizando la prueba de Tukey. Para la variable PCTGEU el estimador del coeficiente de regresión respectivo se obtuvo de la opción SOLUTION del procedimiento Mixed de SAS(5) Para calcular el intercepto de la ecuación de regresión de las variables se utilizó la estimación de la media general de cada carácter. Para cada ecuación, se obtuvo la primera derivada con respecto al porcentaje de genes BT y se igualó a cero para obtener un máximo.
RESULTADOS Y DISCUSIÓN
Efectos ambientales
El nivel de significancia de los efectos principales, las interacciones de primer orden y las covariables de las características reproductivas se presentan en el Cuadro 1; mientras que las medias de cuadrados mínimos de días al primer servicio (DPS), días abiertos (DA) e intervalos entre partos (IEP) se muestran en el Cuadro 2. El número de parto influyó significativamente (P<0.05) en DPS, DA e IEP. Las vacas de cuarto parto tuvieron valores inferiores de DPS con respecto a las de segundo y tercer parto; éstas a su vez fueron diferentes a las de primer parto. De forma similar las vacas de segundo o más partos presentaron DA e IEP más cortos (P<0.05) con respecto a las de primer parto. La diferencia en la reducción de días al primer servicio se explica por el crecimiento y desarrollo corporal de las vacas adultas con respecto a las jóvenes. Estos resultados coinciden con los observados por otros autores(6,7) en animales de distintos ambientes tropicales.
El ambiente climático influye en el comportamiento reproductivo de los animales de distintas formas; una forma es alterando el metabolismo del animal y otra es mediante la estacionalidad de la producción de forrajes y alimentos. En el presente estudio, la época de parto influyó (P<0.05) los DPS, DA e IEP (Cuadro 2). Los animales que parieron en la época de lluvias tuvieron DPS en 33 días antes, y consecuentemente también presentaron DA e IEP más cortos en 50 y 30 días, respectivamente. La respuesta se relaciona básicamente con la mayor disponibilidad de forrajes y alimentos para los animales; y también por el mejoramiento del ambiente, con temperaturas más confortables y menos cambiantes que favorecen un confort metabólico apropiado para la actividad reproductiva de los animales. Los resultados del presente estudio son contradictorios a los obtenidos por Vite–Cristóbal et al(8) y López–Ordaz et al(9), quienes observaron que la estación de parto no influyó en la EPP, DPS, DA e IEP de bovinos Cebú cruzados con Holstein o Suizo en las condiciones tropicales del norte de Veracruz.
Efecto del porcentaje de genes Bos taurus
Algunos estudios(8,9,10) sugieren que el grupo racial de la vaca influye positivamente en la actividad reproductiva de los animales provenientes de los cruzamientos de BI X BT. En el presente estudio se observó que la covariable proporción de genes BT influyó (P<0.05) la EPP, los DPS, DA e IEP en los animales BI.
La Figura 1 muestra el efecto lineal (P<0.05; R2 = 0.79) de PCTGEU en la EPP, y se observa que las vacas con 49 % de genes BT tuvieron EPP más cortas (31 meses) comparado con 0, 25, 62.5 y 75 %; dicha proporción fue 13.2, 4.3, 0.3 y 1.0 % más alta para los animales con 0, 25, 62.5 y 75 % de genes BT, respectivamente. La EPP de 35 meses fue similar a la observada en las vacas con 0 % de genes europeos. La variabilidad de los resultados obtenidos para EPP se explica parcialmente por las diferencias en crecimiento, madurez sexual y de adaptación al ambiente que se presentan entre BT y BI. Por ejemplo, Martin et al(11) observaron que los animales BI maduran reproductivamente a tasas más bajas que las vaquillas con genes europeos. López et al(12) observaron que vaquillas Brahman, Angus y Brangus presentaron una tasa de madurez sexual diferente, de forma que las Angus alcanzaron la pubertad a una edad menor que las Brangus y Brahman.
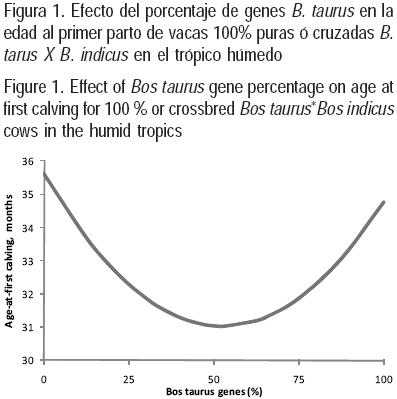
Los resultados de EPP obtenidos en el presente estudio son similares a los observados en otros (8,9,13) donde encontraron reducciones similares en EPP conforme incrementó el porcentaje de BT en los animales cruzados con razas locales; en los mismos estudios se observa que el genotipo ½BT ½BI presentó las mejores EPP.
Como se observa en las Figuras 2 y 3, los DPS y DA tuvieron comportamientos similares. Los valores más bajos (P<0.05; R2 = 0.74) de 148 y 196 días para DPS y DA, respectivamente, fueron observados en los animales 100 % BI. Posteriormente, conforme se incorporan genes BT en cruzamientos con las hembras BI, ambas variables incrementan linealmente hasta alcanzar valores máximos de 224 y 320 días para DPS y DA, respectivamente, que corresponden a los genotipos con 100 % de BT. Lo anterior sugiere que las vacas 100 % BI tienen características reproductivas mejores cuando se comparan con vacas cruzadas con cualquier proporción de genes europeos. Los resultados obtenidos en el presente estudio coinciden con los de otras investigaciones(10,14) en donde se afirmó que los DA incrementan hasta 100 % conforme la inclusión de genes europeos incrementó en las cruzas con razas cebuinas.


En el presente estudio, el mejor comportamiento reproductivo observado en Bos indicus, se explica fundamentalmente por la mayor capacidad de adaptación de dichos genotipos a los ambientes tropicales. En las condiciones tropicales del norte de Veracruz(12) observaron que los genotipos con 25 % genes BI tuvieron EPP 60 días más temprano, los DA fueron 50 días más cortos y los IEP se redujeron en 50 días comparados con los genotipos con 50 % de genes BI. Al respecto, Chenoweth(15) señaló que los bovinos BI se adaptan mejor a los ambientes tropicales, lo cual explica parcialmente las diferencias fisiológicas y endocrinas de las hembras BI X BT en ambientes tropicales.
En la Figura 4 se observa el efecto lineal (P<0.05; r2= 0.67) de la PCTGEU en IEP. El IEP incrementó conforme el porcentaje de genes europeos aumentó. El valor mínimo de 15.7 meses se obtuvo en las vacas con 0 % de genes BT. Además, se observó que conforme el porcentaje de genes BT incrementó, también el IEP aumentó en forma lineal hasta alcanzar un valor máximo de 19.4 meses en vacas con 100 % BT. Los resultados observados en el presente estudio sugieren que el comportamiento reproductivo de las vacas 100 % Bos indicus fue mejor que las vacas cruzadas con cualquier porcentaje de genes BT.

El IEP depende de tres componentes: los días abiertos, la duración de la gestación y el periodo seco. Basado en que la duración de la gestación tiende a ser constante y que el periodo seco es una función resultante del manejo de hato, el componente más influyente en el IEP es el número de días abiertos. Duarte–Ortuño et al(16) indicaron que el IEP no está asociado con factores hereditarios, en virtud de que el índice de herencia para dicha variable es cercano a cero en bovinos de doble propósito. Con base en dicha indicación, el incremento en DA en animales con mayor porcentaje de genes BT se explica parcialmente por un incremento en la producción de leche de estos genotipos(2,12).
Los resultados del presente estudio coinciden con otros(10), donde se ha mencionado que los IEP tienden a extenderse a medida que se incrementa la proporción de genes BT. Sin embargo, los resultados del presente estudio no coinciden con lo observado por Rege(17) y Rivera(18), quienes encontraron que los animales con menor proporción mencionan(19) que el IEP no presenta un comportamiento claro a medida que la proporción de genes Bos taurus incrementa.
CONCLUSIONES E IMPLICACIONES
Las vacas con 50 % de genes Bos taurus alcanzaron la edad al primer parto a los 31 meses y fue inferior a las cruzadas con cualquier otra proporción de genes europeos. Los días al primer servicio posparto, días abiertos y el intervalo entre partos fueron más prolongados en las vacas cruzadas con cualquier proporción de genes europeos con respecto a las vacas 100 % Bos indicus. El comportamiento reproductivo de vacas con 50 % de genes BI permite recomendarlos como los más apropiados para los ranchos ganaderos en los trópicos del sureste mexicano. Sin embargo, la asociación con producción de leche, manejo general del hato y mejoramiento del ambiente se relaciona más con genotipos con 50 ó 60 % de genes Bos taurus.
LITERATURA CITADA
1. Madalena FE. La utilización sostenible de las hembras F1 en la producción del Ganado lechero tropical. Estudio:FAO Producción y Sanidad Animal No. 111. Rome, Italy. 1993. [ Links ]
2. López OR, García CR, García MJG, Ramírez VR. Producción de leche de vacas con diferente porcentaje de genes Bos taurus en el trópico mexicano. Téc Pecu Méx 2009;47(4):435–448. [ Links ]
3. McDowell RE, Wilk JC, Talbott CW. Economic viability of crosses of Bos taurus and Bos indicus for dairying in warm climates. J Dairy Sci 1996;79:1292–1303. [ Links ]
4. García E. Modificaciones a la clasificación climática de Köppen. Instituto de Geografía, México, DF. Universidad Nacional Autónoma de México. 1988. [ Links ]
5. SAS®. SAS User's Guide: Statistics (version 8th ed.). Cary NC, USA. SAS Institute Inc. 2002. [ Links ]
6. Hinojosa CA, Franco A, Bolio I. Factores genéticos y ambientales que afectan el intervalo entre partos en un hato comercial en un ambiente tropical sub–húmedo. Prod Anim Trop 1980;5:181–187. [ Links ]
7. Rangel SR. Evaluación de la eficiencia reproductiva en el centro regional tropical de Puyacatengo [tesis licenciatura]. Chapingo, México: Universidad Autónoma Chapingo; 1984. [ Links ]
8. Vite–Cristóbal C, López–Ordaz R, García–Muñiz JG, Ramírez–Valverde R, Ruiz–Flores A, López–Ordaz R. Producción de leche y comportamiento reproductivo de vacas doble propósito que consumen forrajes tropicales y concentrados. Vet Méx 2007;38:63–79. [ Links ]
9. López–Ordaz R, Vite–Cristóbal C, García–Muñiz JG, Martínez–Hernández PA. Reproducción y producción de leche de vacas con distinta proporción de genes Bos taurus. Arch Zootec 2009;58(224):683–694. [ Links ]
10. Villegas–Carrasco MC, Román–Ponce H. Producción de leche durante el proceso de formación de un rancho de doble propósito en el trópico. Téc Pecu Méx 1986;51:51–61. [ Links ]
11. Martin C, Brinks S, Bourdon M, Cundiff V. Genetics effects on beef heifer puberty and subsequent reproduction. J Anim Sci 1992;70:4006–4017. [ Links ]
12. Lopez R, Thomas MG, Hallford DM, Kiesler DH, Silver GA, Obeidat BS, Garcia MD, Krehbiel CR. Case study: Metabolic hormone profiles and evaluation of associations of metabolic hormones with body fat and reproductive characteristics of Angus, Brangus, and Brahman heifers. Prof Anim Sci 2006;22:273–282. [ Links ]
13. Thorpe W, Kangethe P, Rege J EO, Mosi RO, Mwandotto BAJ, Njuguna P. Crossbreeding Ayrshire, Friesian, and Sahiwal cattle for milk yield and preweaning traits of progeny in the semiarid tropics of Kenya. J Dairy Sci 1993;76:2001–2012. [ Links ]
14. Padilla RFJ, Román PH, Castillo RH. Evaluación del comportamiento reproductivo de ganado bovino lechero cruzado con Cebú en clima tropical. Téc Pecu Méx 1984;47:170–175. [ Links ]
15. Chenoweth PJ. Aspects in reproduction in female Bos indicus cattle: a review. Aust Vet J 1994;71:422–426. [ Links ]
16. Duarte–Ortuño A, Thorpe W, Tewolde A. Reproductive performance of purebred and crossbred beef cattle in the tropics of Mexico. Anim Prod 1988;47:11–20. [ Links ]
17. Rege OJE. Utilization of exotic germplasm for milk production in the tropics. Proceedings 6th World Congress on Genetics Applied to Livestock Production. Armidale, Australia 1998:25:193–201. [ Links ]
18. Rivera VM. Las experiencias del FIRA en cruzamiento de ganado bovino de doble propósito en el trópico a través de los centros de desarrollo tecnológico. Análisis de los Recursos Genéticos, Ganadería Bovina de doble propósito. Villahermosa, Tabasco. SAGAR. 1998;78–84. [ Links ]
19. Cunningham EP, Syrstad O. Crossbreeding Bos indicus and Bos taurus for milk production in the tropics. FAO Animal Production and Health Paper No. 68. Italy (Rome): FAO Press Inc., 1989. [ Links ]
NOTAS
* Este estudio es parte de la Tesis de Maestría en Ciencias del segundo autor.

