










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de ciencias pecuarias
versión On-line ISSN 2448-6698versión impresa ISSN 2007-1124
Rev. mex. de cienc. pecuarias vol.1 no.1 Mérida ene./mar. 2010
Revisión de literatura
Apomixis y su importancia en la selección y mejoramiento de gramineas forrajeras tropicales. Revisión
Apomixis importance for tropical forage grass selection and breeding. Review
Adrián R. Quero Carrilloa, Javier F. Enríquez Quirozb, Carlos R. Morales Nietoc, Leonor Miranda Jiméneza
Recibido el 2 de agosto de 2007
Aceptado para su publicación el 18 de mayo de 2009
a Colegio de Postgraduados en Ciencias Agrícolas. Campus San Luís Potosí. Salinas, SLP. Iturbide 73, Salinas, SLP, 78660, México. Tel/Fax.01–496–9630240. e–mail: queroadrian@colpos.mx. Correspondencia al primer autor.
b Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias. Campo Experimental Cotaxtla– Sitio Experimental Papaloapan. Isla, Veracruz.
c Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias. Campo Experimental Campana–Madera, Chihuahua.
Resumen
Apomixis es un tipo reproductivo donde la producción de semilla ocurre sin fusión de gametos, resultando por generaciones, en descendencia que genéticamente, es copia fiel del progenitor materno. La apomixis se encuentra ampliamente distribuida entre las gramíneas forrajeras tropicales: Bothriochloa, Bouteloua, Brachiaria, Callipedium, Cenchrus, Dichanthium, Eragrostis, Hyparrhenia, Melinis, Panicum, Paspalum, Pennisetum, Setaria, Tripsacum y Urochloa, entre otros. Las especies apomícticas contienen individuos diploides y poliploides conformando "complejos agámicos", donde las barreras reproductivas quedan delimitadas por la apomixis y el nivel de ploidía, lo que tiene impacto en su aprovechamiento. Individuos diploides mantienen la sexualidad activa y estos son raros fuera del centro de origen de especie. Los individuos poliploides son apomícticos y los individuos sexuales poliploides pueden inducirse en el laboratorio, lo que permite obtener descendencia híbrida. La riqueza genética, originada del centro de diversidad es la base para superar problemas productivos y de estabilidad ecológica: producción durante la sequía, fijación de nitrógeno, calidad, resistencia a plagas y enfermedades; como se ha demostrado en casos exitosos en Brachiaria, Panicum y Cenchrus. Los avances en investigación sobre apomixis han elucidado su biología y diversas técnicas para su manejo. Sin embargo, el impacto de estos avances en la investigación sobre forrajes en México, promueve la recapitulación sobre el aprovechamiento de recursos genéticos y el interés en la generación de híbridos para las necesidades de producción en pastoreo extensivo en México.
Palabras clave: Recursos genéticos, Apomixis, Producción de pastos, Híbridos de pastos, Cultivares forrajeros.
Abstract
Apomixis is a reproductive system resulting in seed production without gametes fusion, and produces, through generations, offspring that is a genetic copy from the maternal progenitor. Apomixis importance is widespread among tropical grasses: Bothriochloa, Bouteloua, Brachiaria, Callipedium, Cenchrus, Dichanthium, Eragrostis, Hyparrhenia, Melinis, Panicum, Paspalum, Pennisetum, Setaria, Tripsacum, Urochloa, among the most important. Apomictic species contain diploid as well as polyploid individuals conforming "agamic complexes" and reproductive barriers limited both by apomixis as well as ploidy level which has a great impact on apomictic species utilization. Diploid individuals maintain active the sexuality and these are rare out of the center of specie's genetic origin center. Polyploid individuals in nature are apomictic, and sexual individuals may be induced to polyploidy in the laboratory, allowing hybrid descendants production. Genetic richness originated within the specie's center of diversity is the basis to overcome productive challenges and ecological stability: dry season productivity, nitrogen fixation, forage quality, pest and disease resistance, etc.; as it has been shown by successful breeding programs for Brachiaria and Panicum, between the most recognized for Latin America, and Cenchrus in USA. Apomixis research advancements has elucidated the biology for this reproductive process as well as the techniques for its manipulation; however, the impact of this advancements on forage research in México has promoted a critical review on forage genetic resources utilization, resulting in an increased interest on hybrid material production to face the requirements for extensive grazing production systems in Mexico.
Key words: Genetic resources, Apomixis, Grass production, Grass hybrids, Forage cultivars.
INTRODUCCIÓN
Apomixis es un método de clonación natural por semilla botánica y se encuentra, principalmente, en cultivos como mango, cítricos, manzanos y diversas gramíneas. Se reporta en más de 400 especies de 40 familias(1); aunque, otros autores indican que se puede encontrar en poco más de 300 especies, en 35 familias(2). La apomixis permite, de forma recurrente, utilizar la semilla producida por el agricultor para no depender de la semilla comercial. La reproducción asexual (por semilla) en pastos, predomina en varias especies en las regiones tropicales, áridas y semiáridas de México, y dado que el mejoramiento de apomícticos obligados es posible por medio de la explotación en forma natural, y también a que existen esquemas para el mejoramiento de gramíneas de reproducción apomíctica, sería relevante su incorporación en programas nacionales de investigación para la obtención de mejores cultivares forrajeros. Por lo anterior, el objetivo de la presente revisión, es presentar y discutir los avances y perspectivas que representa el fenómeno de la apomixis como herramienta para la generación de nuevos cultivares forrajeros.
Que es apomixis?
Apomixis (Apo=sin; mixis=mezcla) es la reproducción clonal que perpetúa la composición genética de una planta o grupo de individuos. Evitar la meiosis y la fertilización son características de la apomixis gametofítica, la cual puede definirse como la reproducción asexual por semilla (agamospermia), bajo un esquema en el que se mantiene la alternancia de generaciones (gametofitoesporofito–gametofito) sin la alternancia de fases nucleares (2n–n–2n) como ocurre en plantas sexuales(3), e incluye dos tipos: 1) Apomixis esporofítica, la cual incluye la formación de sacos embrionarios a partir de células no reducidas nucelares, no reportada en pastos. No se debe confundir con la reproducción con otros tejidos somáticos (esquejes o macollos) diferentes al saco embrionario y común en pastos que no producen semilla estéril o de baja viabilidad, tales como: estrella de África Cynodon plectostachyus (K. Schumann) Pilger, pangola Digitaria decumbens Stent, Para Brachiaria mutica (Forsskal) Stapf, Guatemala Tripsacum andersonni Sinónimo T. Laxum Nash, Alemán Echinochloa polystachya (H.B.K.) Hitchcock, Taiwan Pennisetum purpureum Schumacher, entre los de mayor importancia. 2) Apomixis gametofítica, donde la producción de semilla botánica ocurre sin la fusión de gametos, produciendo descendencia vía partenogénesis de células huevo ameióticas (diploides) o nucelares (somáticas diploides), cuya descendencia es copia fiel de la planta madre; esto es, la semilla no es garantía de nuevos genotipos, como ocurre en la reproducción sexual.
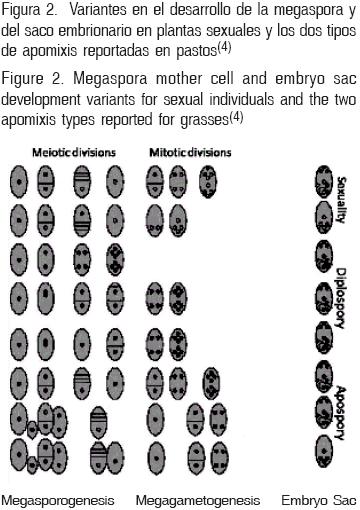
En las plantas, durante el desarrollo del gameto femenino ocurren dos fenómenos particulares: 1) Megaesporogénesis, que dará origen a la célula huevo o célula madre de la Megáspora (CMM) reducida (haploide en especies sexuales) a partir de una célula arquesporial diploide, posterior a una meiosis completa. 2) Megagametogénesis, que dará origen al saco embrionario (SE, normalmente octonucleado: con dos sinérgidas y la ovocélula, dos núcleos polares y tres antípodas), donde ocurrirá la doble fecundación que dará origen al endospermo triploide y al embrión diploide. Aunque existen variantes en el reino vegetal(3,4), éstos son los fenómenos más difundidos en las plantas de reproducción sexual (amfimícticas). Para el objeto de esta revisión se referirá como apomixis la producción de semilla clonal, esto es, la apomixis gametofítica (apogamia, agamospermia, apogametia). Existen diversos tipos de apomixis gametofitica y toda una gama de variaciones y combinaciones posibles en el desarrollo de la CMM y del gametofito(4,5,6,7) (Figura 1).
TIPOS DE APOMIXIS GAMETOFÍTICA
En semillas apomícticas, el embrión se origina de variaciones en el origen de la célula que origina la célula huevo o CMM (megagametogénesis); el cual puede seguir dos caminos en pastos 1) Diplosporia, en los géneros: Paspalum, Tripcacum, Eragrostis, Elymus, etc., con meiosis parcial, incompleta o restitutiva que resulta en una célula ameiótica (no reducida) y 2) Aposporia en Panicum, Paspalum, Brachiaria, Bouteloua, Cenchrus, Pennisetum, etc., donde la CMM puede ser eliminada o no, y es reemplazada por una célula somática (diploide) de la pared nucelar del ovario; existiendo, similarmente, variaciones en estos fenómenos. Es suficiente establecer estos dos orígenes de la CMM y del SE. Al parecer, la aposporia es dominante en Poaceae y es el mecanismo reproductivo más evolucionado entre especies apomícticas(6); (Figura 1). Existen 20 géneros de gramíneas considerados de alta importancia por su contribución a la alimentación del ganado en regiones tropicales; de éstas, 11 se reproducen por apomixis (Cuadro 1) y con el tipo apospórico como predominante(8).
En la mayoría de los óvulos apospóricos, la CMM resultante de la meiosis muere o es eliminada por competencia con células somáticas, que toman la función de CMM y esta célula somática no reducida da origen al SE tetranucleado (Brachiaria, Panicum, Cenchrus, Paspalum, Pennisetum) generalmente múltiples en un solo ovario(7,9). El gametofito o SE resultante, contiene dos sinérgidas, un núcleo polar y una célula huevo(5), lo que facilita la diferenciación de individuos sexuales y apomícticos dentro de un complejo agámico (Figura 2).
La microgametogénesis (producción de polen o gametos masculinos) en especies apomícticas es normal, produciéndose gametos meióticos sin importar el nivel de ploidía de la planta madre(10) y, debido a las altas inversiones energéticas de la planta en la producción de polen, cualquier grano de polen con aberraciones meióticas, es sustituido por otro normalmente desarrollado. En especies apomícticas ocurren tres fenómenos biológicos(6):
Funcionamiento de una célula no reducida (ameiótica) como CMM para el desarrollo del SE; los genotipos apomícticos tienen la ventaja aparente de incrementar su descendencia sobre los individuos sexuales, mediante la circunscripción de la meiosis femenina(11,12). b) Requerimiento de la polinización para el desarrollo del endospermo, en la mayoría de los pastos apomícticos (pseudogamia); en todas las especies apomícticas pseudógamas, la producción de semilla requiere del polen para la formación del endospermo viable, aunque el embrión se desarrolle partenogenéticamente(2). c) Desarrollo partenogenético del embrión, lo que resulta en descendencia que es copia fiel de la planta madre.
DIVERSIDAD Y EVOLUCION DE GRAMINEAS APOMICTICAS
La apomixis había sido considerada como un callejón sin salida de la evolución(13); sin embargo, actualmente se ha comprobado que la apomixis representa una herramienta valiosa para la selección de individuos ampliamente adaptados a condiciones ambientales dinámicas y como promotora de la perpetuación y dominancia de (los) genotipo(s) híbrido(s) (genet) de mayor aptitud a dichas condiciones. La dinámica evolutiva es muy activa en los centros de origen de especie, donde se ha demostrado un flujo genético entre la diversidad de niveles de ploidía(11,14). Durante los últimos años se han logrado avances en la interpretación de la apomixis, tanto en técnicas de laboratorio como en el manejo de la diversidad natural(4). En los centros de origen de especie, se encuentran individuos de diversos niveles de ploidía: diploides, euploides, siendo el tetraploide el de mayor abundancia, e individuos aneuploides resultado de la dinámica del flujo genético. Esta dinámica reproductiva, ocurre mediante la presencia cíclica de cruzamientos entre individuos con diferentes niveles de ploidía(14,15,16). Estas formas de recombinar de las plantas apomícticas, mediante ciclos diploide–tetraploide–diaploide, e identificados inicialmente en Bothriochloa(14) y posteriormente en Panicum maximum Jacq.(11), ha sido reportada en géneros como Brachiaria(17). En las especies apomícticas, las plantas sexuales son de baja frecuencia y normalmente poseen menor vigor que los individuos apomícticos(10). Esta asociación de poliploidía y vigor, confiere a la apomixis mayor aptitud para las condiciones ambientales limitantes para la producción.
En México, la evaluación tradicional en "jardines de introducción" multiespecíficos fue un paso importante para el conocimiento de la diversidad de especies; sin embargo, estos enfoques resultan poco eficientes para determinar el potencial de una especie valiosa para la diversidad de regiones fisiográficas que se tienen en el país. El paso siguiente, después de detectar la especie mejor adaptada a las condiciones regionales, es la evaluación de la diversidad dentro de dicha especie. Esto es, un "jardín de introducción" con muchos ecotipos originarios del centro de origen de especie, de un solo complejo agámico o una sola especie(18).
APOMIXIS EN PASTOS FORRAJEROS
Las plantas apomícticas presentan diversas ventajas adaptativas: producen descendencia fértil sin importar su estructura o balance cromosómico, asocian la fijación del vigor híbrido mediante semilla durante de generaciones. Similarmente, la apomixis facilita la producción de híbridos, toda vez que se estructuren adecuadamente los programas de mejoramiento genético, eliminando el manejo especial de líneas progenitoras, como ocurre en cultivos sexuales, reduciendo costos(2). Otras ventajas prácticas del manejo de la apomixis, incluyen aspectos que modifican la filosofía del aprovechamiento de especies forrajeras apomícticas e incluyen:
1) Revaloración de la importancia de los centros de origen de especie como fuente de riqueza genética de la especie de interés; estos se encuentran donde ocurre la mayor dinámica de flujo genético entre diversos niveles de ploidía. La importancia de recolectar la diversidad genética en los centros de origen de especie, ya había sido seńalada para cultivos básicos y apreciada ligeramente para la selección de materiales de especies forrajeras; sin embargo, es en el caso de especies apomícticas donde el impacto de la diversidad natural de los centros de origen de especie, es inmediato, como los casos de colectas de pasto Guinea por investigadores japoneses y franceses(19,20), Brachiaria(21), Paspalum en Argentina(22) y en México Bouteloua curtipendula Michx. Torr.(23,24) y Tripsacum spp.(25). La importancia de recolectas representativas de especie, ha sido constatada en cada ocasión que se evalúan las especies apomícticas con este esquema. Tres tipos de poblaciones localizadas en el centro de origen de especie han sido reportadas(26): a) monomórficas, b) polimórficas con variación discontinua y genotipos diferentes y c) polimórficas con variación continua, similar distribución fue reportada en el género Tripsacum en México(27). En las especies en que prevalece la apomixis, las plantas sexuales son raras(10). La abundancia de individuos diploides y, por tanto, con reproducción sexual ha sido localizada en los centros de origen(28) en especies como Chloris gayana Kunth(29), P. maximum(30,31), Tripsacum(25) y Brachiaria(32), mientras que, en Cenchrus ciliaris L. Link una planta sexual tetraploide espontánea ha dado origen al programa de mejoramiento genético de la Universidad de Texas A&M(10); esto es, no se han localizado individuos diploides, posiblemente porque no se ha muestreado su centro de origen genético. En condiciones naturales, estos individuos sexuales diploides exhiben un menor vigor que sus congéneres poliploides simpátricos(10).
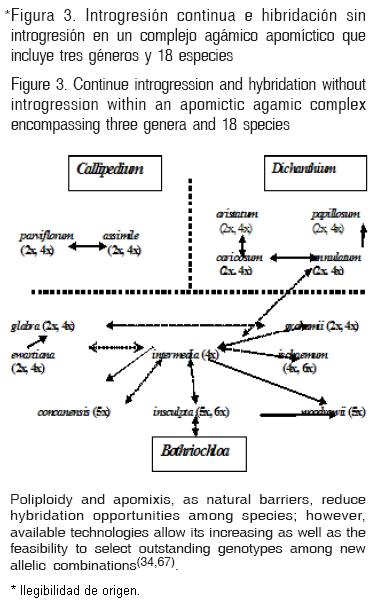
2) Conformación de Complejos Agámicos (CA). Muchas especies apomícticas consideradas como diferentes y aisladas reproductivamente, se integran en un CA, en donde la única barrera para lograr hibridación es la presencia de apomixis y poliploidía, las cuales, al superarlas mediante estrategias reconocidas, amplían la base genética para la producción de híbridos que respondan a las necesidades detectadas e indicaban la hibridación intergenérica viable entre los géneros Bothriochloa y Dichanthium, en la que diversas especies funcionan como "puente" para lograr la hibridación(33), posteriormente este CA, incluyó al género Capillipedium(34) como constituyente de esta macroespecie no aislada por otro factor más allá de la poliploidía y apomixis (Figura 3). Otros CA reconocidos en pastos tropicales incluyen a P. maximum, P. trichocladum Hack ex. Schum. y P. infestum Andersson, con formas diploides sexuales y poliploides apomícticas(19), además de amplia diversidad de morfotipos intermedios(35). En el género Paspalum, existen varios CA conformados por especies con abundancia de niveles de ploidía(36,37). En el género mexicano Tripsacum 11 especies nativas de Mesoamérica conforman el CA(27) y pueden producir híbridos, eliminadas las barreras mencionadas. En pasto Buffel, el CA está conformado por al menos dos especies: C. ciliaris y C. setigerus Vahl.(38), aunque existen indicios de otras especies que pueden ser incorporadas al CA como Pennisetum flaccidum Hochst. ex Steud. y P. mezianum Leeke(39,40). Por otra parte, la conformación del CA de la sección Curtipendula de B. curtipendula, puede contener 11 especies con riqueza de niveles de euploidía y aneuploidía, donde se siguen esquemas de selección y mejoramiento genético, desarrollado para otras especies apomícticas(41). En el género Brachiaria, tres especies conforman el CA con Brachiaria decumbens Stapf.: B. brizantha (Hochst.) Stapf. y B. ruziziensis Germ. & C. Eur., la sexualidad poliploide, ha sido la base para producir híbridos(42).
La generación de híbridos en especies apomícticas incluye los siguientes procedimientos: a) los individuos sexuales diploides, toda vez disponibles en la especie de interés, son poliploidizados en laboratorio mediante cualquier agente inhibidor de la formación del uso acromático en la mitosis, induciendo la sexualidad poliploide; b) los poliploides polinizadores siempre son apomícticos y la viabilidad del polen no es problema cuando se usan individuos al mismo nivel de ploidía que la planta hembra; c) la segregación de individuos apomícticos y sexuales es 1:1 en la descendencia obtenida, los individuos sexuales genéticamente recombinados pueden integrarse al grupo de progenitores futuros y, d) los híbridos apomícticos son evaluados para los atributos de interés, dado que el vigor híbrido queda integrado en el genoma.
3) Producción y evaluación de híbridos. La producción de híbridos no es un proceso costoso, ni difícil, por lo que se debe promover la visión de utilización de recursos genéticos adecuados dentro de cada especie de interés con el enfoque de y la producción de híbridos que respondan a las necesidades del mercado (máxima productividad y estabilidad de los ecosistemas en pastoreo) proporcionan las mejores alternativas para mejorar las condiciones de vida de la población, en un mercado globalizado que ha devastado marcadamente los ecosistemas. El aprovechamiento sistemático de recursos genéticos nativos de México, algunos apomícticos y otros de reproducción sexual incluye materiales valiosos en Tripsacum, Setaria, Lycurus, Leptochloa, Sporobolus, Distichlys, Hymenachne, Digitaria, entre los de mayor potencial e inexplorados hasta hoy.
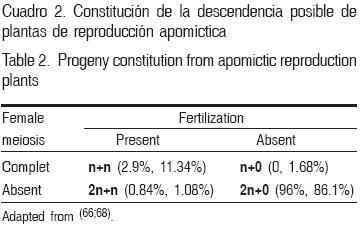
La apomixis permite a las plantas poliploides mantener la producción de semilla viable, proporcionando una estrategia de escape a la esterilidad, debido al desbalance cromosómico embrión:endospermo(6). La descendencia producida por individuos apomícticos será apomíctica en gran proporción (Cuadro 2); mientras que una proporción (50 %) de la descendencia de plantas sexuales, en cruzas dirigidas, será apomíctica(11). La fertilización tipo 2n+n ocurre frecuentemente dentro de los CA y ha recibido atención como método de mejoramiento genético cuando no se dispone de individuos sexuales. En el CA Cenchrus–Pennisetum el vigor de los híbridos 2n+n recuperados ha sido reportado como excelente(43,44); asimismo, híbridos 2n+n habían sido recuperados en Dichanthium annulatum (Forssk.) Stapf. y Bothriochloa(16,33). En Cynodon, algunos de los híbridos más productivos fueron tetraploides, posiblemente derivados de la cruza entre C. dactylon var. afghanicus Harlan & de Wet o C. dactylon var. aridus Harlan & de Wet con el tetraploide polinizador C. dactylon (L.) Pers. Var. dactylon(16,45). Es importante reconocer el valor de estos aspectos para obtener el máximo beneficio para las especies forrajeras de interés, con recursos genéticos adecuados a los requerimientos(31); en ocasiones, sin necesidad de mejoramiento genético, como en el CA de Brachiaria spp.(46), con recursos genéticos que han sentado las bases para la producción de híbridos para mejorar la producción, calidad de forraje y resistencia a mosca pinta(42); en buffel, los investigadores norteamericanos han desarrollando variedades que combinan resistencia a bajas temperaturas y sequía(33).
MEJORAMIENTO GENETICO DE ESPECIES APOMICTICAS
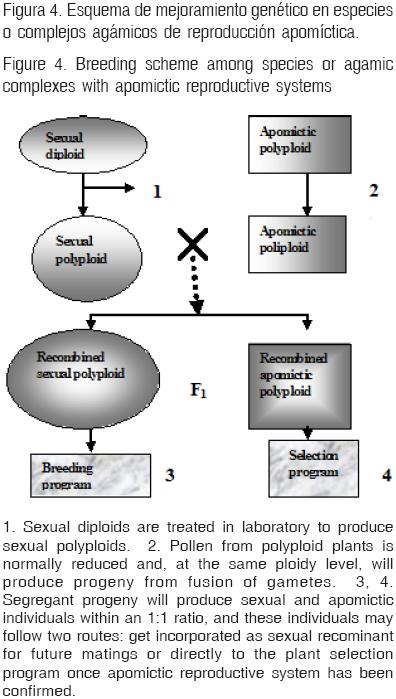
Los procesos básicos de un programa de mejoramiento genético en especies apomícticas incluyen: a) recolección y disponibilidad de recursos genéticos del centro de origen de especie; b) caracterización morfológica y citológica de las poblaciones (identificación de individuos diploides y poliploides); c) definición del tipo reproductivo de los individuos de la colección d) producción de individuos sexuales poliploides en el laboratorio, con el fin de aprovechar la variabilidad genética mediante la recombinación meiótica en la producción de la célula huevo y la recombinación genética resultante de la fusión de gametos (Figura 4); e) programa de cruzamientos con objetivos bien definidos, para lograr el mayor avance genético y f) programas de evaluación agronómica de los materiales en conjunto con la ampliación de la base genética con el aprovechamiento de la descendencia sexual.
La apomixis no ha sido reportada en individuos diploides, en especies o CA apomícticos, todas las plantas diploides son sexuales(3). La preservación del genoma y la expresión de vigor híbrido en la descendencia que resulta de la fusión de gametos, el cual queda integrado a éste por generaciones mediante la reproducción asexual por semilla, proporciona a la apomixis ventajas de mayor aptitud evolutiva(47,48,49), al proporcionar una herramienta para recombinar con cierta frecuencia, condicionada por los niveles de ploidía disponibles (diversidad genética), dada la recurrencia de la sexualidad (mediante recombinación meiótica y fusión de gametos) de las plantas apomícticas, permitiendo la promoción de dominancia de combinaciones alélicas responsivas a ambientes desafiantes.
EXPERIENCIAS EN EL MEJORAMIENTO GENETICO DE GRAMINEAS
Los programas de mejoramiento en gramíneas forrajeras se habían enfocado a la selección directa de ecotipos apomícticos, variedades de pasto Guinea seleccionadas en Australia, como Green Panic, Gatton Panic; en Cuba: Likoni; en Brasil: Mombaza, Tobiata, Tanzania, Coloniao, etc. habían sido seleccionadas de la diversidad natural. Existen reportes que indican que el cultivar de Guinea "Natzukaze" es de origen híbrído(50) y éste podría ser el primer híbrido producido en pasto Guinea. Variedades populares en el género Brachiaria incluían ecotipos seleccionados de la diversidad natural: Marandu y Basilisk. En los últimos años, se han iniciado diversos programas de producción de híbridos y la riqueza genética recolectada en África(21) ha producido nuevos cultivares: La Libertad, Toledo (MG4 y MG5 en México) y el primer hibrido "ínterespecífico" de Brachiaria denominado Mulato. Por otra parte, las líneas Laredo y Pecos son materiales híbridos en pasto buffel, al igual que el individuo sexual espontáneo descubierto en el rancho de Pat Higgins(44). Individuos sexuales diploides no han sido recolectados en África para buffel. Las variedades iniciales que se generaron en pasto buffel fueron ecotipos recolectados de la diversidad natural: Gaindah, Molopo, Nunbak, Lawes, Americano, Mbalambala, Tarewinnbar, Higgins, Frío, se han incorporado de la riqueza natural, sin hibridación. Los primeros híbridos en esta especie fueron Llano y Nueces, buscando resistencia a bajas temperaturas.
Buffel fue seńalado como nativo de África tropical, subtropical, India e Indonesia(29) y el equipo de mejoramiento genético de la Universidad de Texas A&M, ha indicado su origen en Sudáfrica(51); sin que descubrieran individuos sexuales en las áreas de recolecta. No se han llevado a cabo expediciones de recolección de buffel en el Sahel africano, seńalado como su centro de origen(52,53,54). Buffel es un recurso forrajero en espera de un programa de recolección y conservación de la diversidad genética, que promete buena rentabilidad, una vez que se hayan generado ecotipos o híbridos sobresalientes. Para el caso de México tropical, es evidente que no se requiere mayor resistencia a frío, en buffel, más allá de la resistencia natural, tal vez la de Molopo fuese suficiente aún para regiones del norte del país, donde el invierno no es tan riguroso en intensidad y duración como en regiones áridas centrales de Estados Unidos, por lo que nuestros objetivos de selección y producción de híbridos serán diferentes a las características de las variedades mejoradas producidas por el equipo de Texas A&M; por tanto, disponer de riqueza genética de la especie para un programa propio de mejoramiento acorde a las necesidades de los productores en México, es básico. En Brachiaria, el primer objetivo es la resistencia al complejo de mosca pinta(42) y la diversidad genética ha sido la mejor opción y en México, debemos estar alerta a esta plaga de gran importancia, tanto para buffel en Sonora, como para la creciente superficie de Brachiaria en regiones tropicales.
La definición del comportamiento de la heredabilidad de la apomixis es importante para su manipulación hacia cultivos básicos, de gran interés entre los centros de investigación en diversas partes del mundo(8). El control genético de la apomixis había sido definido como de herencia simple monogenética en especies vegetales no relacionadas Ranunculus y Panicum, con resultados similares, la apomixis está controlada por un gen simple dominante(5,55) y, en buffel(56), modificando los resultados de dos genes implícitos reportados para buffel(57). La ineficacia de incorporarla a cultivos básicos sexuales pone de manifiesto mayores requerimientos de investigación y la posibilidad de que se trate de genes estrechamente ligados(54,58).
OBJETIVOS PARA EL MEJORAMIENTO GENÉTICO
El fin último de la utilización de la apomixis, es responder a las presiones de utilización en pastoreo y lograr la máxima estabilidad ecológica y productividad. Existen diversos problemas y a todos ellos, la diversidad natural tiene una respuesta viable, el problema radica en saber localizar la problemática importante y disponer de estrategias adecuadas de cuestionamiento (evaluación de) a la diversidad natural. Se describen a continuación algunos aspectos urgentes para lograr estabilidad de ecosistemas y productividad sostenida.
Fertilización nitrogenada
Ningún ganadero fertiliza praderas de temporal, mucho menos pastizales de zonas áridas, donde el uso de nitrógeno derivado de energía fósil, resulta en inversiones difíciles de solventar. Por tanto, los ganaderos sufren de deficiencia de N en sus praderas. La utilización de leguminosas fijadoras de N es una opción viable e inmediata para solventar esta necesidad; sin embargo, las leguminosas requieren de cuidados específicos que la cultura ganadera de los productores no maneja en forma eficiente(18). Otra alternativa para incorporar N al sistema es la fijación de N mediante la asociación con Azospirillum spp., que puede promoverse entre los pastos. La afinidad diferencial entre los genotipos de una especie forrajera con Azospirillum, remarca la importancia de la disponibilidad de recursos genéticos para su evaluación en asociación con la bacteria y fijación de N.
Plagas y enfermedades
En Latinoamérica tropical el complejo mosca pinta (Aenolamia, Deois, Mahanarva, Zulia, entre los principales géneros) es un problema latente para años de alta humedad. Mosca pinta en buffel se presenta regularmente en condiciones de trópico seco y puede convertirse en un serio problema para las praderas del estado de Sonora, como ha ocurrido en B. decumbens en Brasil(59), donde se perdieron grandes extensiones de praderas en sólo dos semanas de alta infestación de la plaga. En México 600,000 ha han sufrido daños marcados por esta plaga, principalmente en el estado de Sonora en praderas de pasto buffel y en el estado de Veracruz en diversas especies tropicales(60,61). Igualmente, las enfermedades más serias para pasto buffel son las provocadas por diversas especies de Claviceps, que pueden destruir la semilla o resultar en bajos rendimientos y calidad de la misma. En el sur de Texas se han reportado ataques de Pyricularia(62).
Valor nutritivo del forraje
Avances en el mejoramiento de la digestibilidad del forraje han sido logrados en Bermuda "Coastcross". Después de varios ciclos de Selección Recurrente Restringida, se logró una digestibilidad mayor a 12 % de la materia seca, con respecto a Bermuda "Coastal"(63), lo que se reflejó en hasta 40 % más de aumentos de peso; al disponer de segregaciones alélicas, la digestibilidad de los forrajes es factible de mejorarse(64). Por otra parte, la digestibilidad in vitro de la materia seca en "Coastcross" no se redujo durante seis ciclos de selección fenotípica restringida recurrente hacia alto rendimiento de forraje(65), lo que es indicativo que el carácter de mayor digestibilidad del forraje, seleccionado previamente, no se modificó toda vez que se continuó la presión de selección en la población hacia otro carácter, como lo fue el alto rendimiento de forraje.
ESTACIONALIDAD EN LA PRODUCCIÓN DE FORRAJE
La diversidad natural es responsiva a las necesidades de mejoramiento genético regidas por los requerimientos antropocéntricos hacia la pradera o pastizal. En pasto Guinea(31) evaluaron la diversidad de P. maximum recolectada en su centro de origen genético por ORSTOM, actual IRD–Francia; en esta especie, encontraron variación para rendimiento de forraje (8 a 13 % del total anual) durante el periodo de sequía, lo cual resulta significativo, dado que en los trópicos es durante esta época que la disponibilidad de forraje se vuelve un factor determinante de la producción. En la diversidad genética de Brachiaria evaluada en Brasil se ha indicado una variación para rendimiento de forraje en tiempo de secas con respecto al total anual de 9 a 27 % en Brachiaria brizantha, 10 a 25 % en B. decumbens, 9 a 18 % en B. humidicola (Rendle) Schweick., 9 a 19 % en B. ruziziensis y 7 a 22 % en B. jubata (Fig. & De Not.) Stapf.(42). Estos valores son indicativos de la amplia oportunidad de mejoramiento, toda vez que se tiene acceso a recursos genéticos representativos de la diversidad natural.
CONCLUSIONES
La disponibilidad y manejo de recursos genéticos es una herramienta importante, ya que ha dado resultados valiosos como la generación de nuevos cultivares en especies de los géneros: Brachiaria, Panicum y en menor proporción en Cenchrus. La tecnología generada por los programas de incorporación de apomixis a especies de cultivos básicos puede ser aplicada en el mejoramiento de gramíneas forrajeras apomícticas. Las estrategias de manejo de especies apomícticas deben ser asimiladas en México, para lograr avances sólidos en el desarrollo de cultivares forrajeros, adecuados a las necesidades particulares de los productores y del país. Tener objetivos claros del mejoramiento, se reflejará en un rápido avance, dado que la diversidad ofrece amplias combinaciones alélicas para responder eficientemente a las demandas antropocéntricas hacia las praderas en pastoreo.
LITERATURA CITADA
1. Jessup RW. Molecular tools for marked assisted breeding of buffelgrass. [Ph. D. Dissertation]. Texas A&M University. Col. Station, TX; 2005. [ Links ]
2. Bashaw EC, Hanna WW. Apomictic reproduction. In: Chapman G editor Reproductive versatility in the grasses.1st ed. Cambridge, UK Cambridge University Press; 1990:100–130. [ Links ]
3. Nogler GA. Genetics of gametophytic apomixis: a historical sketch. Pol Bot Stud 1994;(8):5–11. [ Links ]
4. Crane CF. Classification of apomictic mechanisms. In: Savidan Y, Carman JG, Dresselhaus T editors. The flowering of apomixis: from mechanisms to genetic engineering; 1st ed. Mexico, DF: CIMMYT, IRD, European Comission DG VI (FAIR) 2001:24–43. [ Links ]
5. Nogler GA. Gametophytic apomixis. In: BM Johri editor. Embryology of angiosperms. 1st ed. Berlin, Germany: Springer Verlag 1984;475–518. [ Links ]
6. Asker SE, Jerling L. Apomixis in plants. 1st ed. Boca Ratón, Florida, USA: CRC Press; 1992. [ Links ]
7. Savidan YH. Apomixis: Genetics and breeding. Plant Breed Rev 2000;(18):13–86. [ Links ]
8. Savidan YH. Transfer of apomixis through wide crosses. In: Savidan Y, Carman JG, Dresselhaus T editors. The flowering of apomixis: from mechanisms to genetic engineering; 1st ed. Mexico, DF: CIMMYT, IRD, European Comission DG VI (FAIR) 2001;153–167. [ Links ]
9. Gustafsson A. Apomixis in angiosperms II. L. Univ. Arsskr. II; 1947;43:71–179. [ Links ]
10. Hanna WW and Bashaw EC. Apomixis, its identification and use in plant breeding. Crop Sci 1987;27(6):136–1139. [ Links ]
11. Savidan YH, Pernés J. Diploid–tetraploid–dihaploid cycles and the evolution of Panicum maximum Jacq. Evolution 1982;(36):596–600. [ Links ]
12. Silvertown J. Introduction to plant population ecology. 1st ed. Essex, England: Longman Scientific & Technical; 1987. [ Links ]
13. Darlington CD. Evolution of genetic systems. 1st ed. London, England: Oliver & Boyd; 1939. [ Links ]
14. de Wet JMJ. Diploid– tetraploid– haploid cycles and the origin of variability in Dichanthium agamospecies. Evolution 1968;(22):394–397. [ Links ]
15. de Wet JMJ, Stalker HT. Gametophytic apomixis and evolution in plants. Taxon 1974;(23):689–697. [ Links ]
16. Harlan JR, JMJ de Wet. On Ö. Winge and a prayer: The origins of Polyploidy. Bot Review 1975;41(4):361–390. [ Links ]
17. de Wet JMJ. Hybridization and polyploidy in the Poaceae. In: Soderstrom TR, Hilu KW, Campbell CS, Barkworth ME editors. Grass systematic and evolution 1st ed. Washington, DC USA; Smithsonian Institution Press; 1986:188–194. [ Links ]
18. Quero CAR, Enríquez JFQ, Miranda LJ. Evaluación de forrajeras en México. Avances o status quo. Interciencia 2007;32(8):566–571. [ Links ]
19. Combes D, Pernés J. Variations dans les nombres chromosomiques du Panicum maximum Jacq. en relation avec le mode de reproduction. CR Acad Sci Paris 1970;270:782–785. [ Links ]
20. Nakajima K, Ochi M, Mochizuki N. Characteristics and variations of guineagrass strains collected and introduced from Africa. I. Evaluation of characteristics and variations. Bull Natl Grassl Inst Japan 1978;12:38–53. [ Links ]
21. Centro Internacional de Agricultura Tropical. Germoplasma. Informe anual 1985. Pastos Tropicales. CIAT. Cali, Colombia. 1986. [ Links ]
22. Quarin CL, Norrman GA. Relaciones entre el número de cromosomas, su comportamiento en la meiosis y el sistema reproductivo en el género Paspalum. IV Cong Lat de Botánica; Bogotá, Colombia 1987:(3):25–34. [ Links ]
23. Quero CAR. Recursos genéticos forrajeros para zonas áridas. El complejo agámico de zacate banderita [Bouteloua curtipendula (Micx.) Torr.] En: Pérez JP, Hernández A, Gómez A editores. Los forrajes en México: presente y futuro. XXXII Aniversario Especialidad de Ganadería Colegio de Postgraduados en Ciencias Agrícolas. Montecillo, Texcoco, México 2001;25–56. [ Links ]
24. Morales NCR, Quero AR, Hernández A, Pérez J, González SS. Evaluación de la diversidad del pasto nativo Bouteloua curtipendula Michx. Torr. mediante marcadores de AFLP. Agrociencia 2006;40:711–720. [ Links ]
25. Quero CAR, Savidan YH, Berthaud J, Pérez JP, Espinoza JV. Estudios citogenéticos en el género Tripsacum. Agrociencia 1998;23:78–83. [ Links ]
26. Pernés J. Organisation évolutive d'un groupe agamique: la section des Maximae du genre Panicum. ORSTOM París, Tech Rep. 1975. [ Links ]
27. LeBlanc O, Peel MD, Carman JG, Savidan Y. Megasporogenesi and megagametogenesis in several Tripsacum species (Poaceae). Amer J Bot 1995;(82):57–63. [ Links ]
28. Harlan JR. Agricultural origins: Centers and non–centers. Science 1975;(174):468–474. [ Links ]
29. Bogdan AV. Rhodes Grass. Herbage Abstracts 1969;39(1):1– 13. [ Links ]
30. Savidan YH. Genetic control of facultative apomixis and application in breeding Panicum maximum. XIV Intnal Cong Genetics. Moscow, USSR. 1978. [ Links ]
31. Savidan Y, Jank L Costa JCG. Registro de 25 acessos selecionados de Panicum maximum, Brasil. EMBRAPA–CNPGC Doc. 44; 1990. [ Links ]
32. do Valle BC, Costa C. New sexual accessions in Brachiaria. In: Savidan YH, Crane CF editors. Apomixis Newsletter 1991;11–13. [ Links ]
33. Harlan JR, Brooks MH, Borgoankar D, de Wet JMJ. Nature and inheritance of apomixis in Botriochloa and Dichantium. Bot Gaz 1964;(125):41–46. [ Links ]
34. de Wet JMJ, Harlan JR. Apomixis, polyploidy, and speciation in Dichanthium. Evolution 1970;(24):270–277. [ Links ]
35. Assienan B, Noirot M. Inheritance, distribution and biology of andromonoecy in the agamic complex of the Maximae (Panicoideae). Theor Appl Genet 1999;(98):622–627. [ Links ]
36. Norrman GA, Quarin CL, Burson BL. Cytogenetics and reproductive behavior of different chromosome races in six Paspalum species. J Heredity 1989;(80):24–28. [ Links ]
37. Quarin CL. The nature of apomixs and its origin in Panicoid grasses. In: Savidan YH, Crane CF editors. Apomixis Newsletter 1992;(5):7–15. [ Links ]
38. Bashaw EC, Hussey MA, Hignight KW. Hybridization (N+N and 2N+N) of facultative apomictic species in the Pennisetum agamic complex. Int J Plant Sci 1992;153(3):466–470. [ Links ]
39. Hignight KW, Bashaw EC, Hussey MA. Cytological and morphological diversity of native apomictic buffelgrass Pennisetum ciliare (L.) Link. Bot Gaz 1991;152(2):214–218. [ Links ]
40. Quero CAR. Forage production and quality heterosis in 2n+n Pennisetum interspecific hybrids [Ph. D. Dissertation]. TX, USA: Texas A&M University; 2000. [ Links ]
41. Quero CAR, Enríquez QJF. Mejoramiento genético de gramíneas forrajeras en México. Importancia estratégica y avances. En: Flores EC et al editores. I Seminario Internacional de Manejo de Pastizales. UAA–PRODUCE; 2003. [ Links ]
42. do Valle BC, Miles JW. Breeding apomictic species. In: Savidan Y, Carman JG Dresselhaus T editors. The flowering of apomixis: from mechanisms to genetic engineering; Mexico, DF. CIMMYT, IRD, European Comission DG VI (FAIR) 2001;137– 152. [ Links ]
43. Bashaw EC, Hignight KW. Gene transfer in apomictic buffelgrass through fertilization of an unreduced egg. Crop Sci 1990;(30):571–575. [ Links ]
44. Hussey MA, Hatch SL, Bashaw EC. Evolución de las especies y transferencia de genes en el complejo asexual Cenchrus– Pennisetum. En: Clavero T editor. Producción e investigación en pastos tropicales. Univ. Venezuela 1993;133–147. [ Links ]
45. Harlan JR. Cynodon species and their value for grazing and hay. Herbage Abstracts 1970;40(3):233–238. [ Links ]
46. Miles JW, do Valle CB. Manipulation of apomixis in Brachiaria breeding. In: Miles JW, Maass BL, do Valle CB editors. Brachiaria: Biology, agronomy and improvement. Cali, Colombia. CIAT–EMBRAPA Campo Grande, Brazil: CNPGCEMBRAPA 1996;164–167. [ Links ]
47. Gustafsson A. Apomixis in higher plants. Suitzerland: Lunds University; 1946. [ Links ]
48. Bashaw EC, Hovin AW, Holt EC. Apomixis, its evolutionary signification and utilization in plant breeding. Proceed 11th Intnal Grassl Cong. Queensland, Australia. 1970;245–248. [ Links ]
49. Asker SE. History of apomixis research. In: Stelly D editor. Harnessing apomixis. A new frontier in plant science. College Station, TX, USA. 1995. [ Links ]
50. Nakagawa H, Shimizu N, Hanna WW. Cytology of "Natzukaze" guineagrass, a natural apomictic hybrid between a sexual and apomictic plant. J Jap Soc Grassl Sci 1993;39:374–380. [ Links ]
51. Hussey MA, Bashaw EC. Avances en el mejoramiento genético del zacate buffel. Memorias de la IV conferencia internacional de ganadería tropical. Universidad Autónoma de Tamaulipas, Cd. Victoria, México. 1990;12–15. [ Links ]
52. LeHouérou HN. Forage and fuel plants in the arid zone of North Africa, the Near and Midle East. In: Wickens GE, Goodin JR, Field DV editors. Plants for arid lands. London, UK. George Allen&Unwin; 1985;117–141. [ Links ]
53. Voigt PW, MacLauchlan RS. Native and other western grasses. In: ME Heath, RF Barnes and DS Metcalfe editors. Forages: The science of grassland agriculture. Ames, Iowa: Iowa State University Press; 1985;177–187. [ Links ]
54. Savidan YH. Forewords. Apomixis Newsletter 1991;3:41–47. [ Links ]
55. Savidan YH. Chromosomal and embryologycal analyses in sexual X apomictic hybrids of Panicum maximum Jacq. Theor Appl Genetics 1980;(57):153–156. [ Links ]
56. Sherwood RT, Berg CC, Young BA. Inheritance of apospory in buffel grass. Crop Sci 1994;(34):1490–1494. [ Links ]
57. Taliaferro CM, Bashaw EC. Inheritance and control of obligate apomixis in breeding buffelgrass, Pennisetum ciliare. Crop Sci 1966;(6):473–476. [ Links ]
58. Savidan Y. Foreword. Apomixis Newsletter [on line]. http://92.100.189.39/CIMMYT/Biotechnology/apomixisNewsletter/9/editoria.htm Accessed April 15, 1996. [ Links ]
59. Marcondes WF. O drama da Braquiaria. Criadores–Brasil 1978;(576):13–16. [ Links ]
60. López CA. Identificación, distribución, importancia económica y potencialidad del complejo mosca pinta en praderas de pasto buffel en Hermosillo, Sonora [tesis Licenciatura]. Monterrey, México: ITESM; 1984. [ Links ]
61. Ibarra F, Cox JR. Necesidades de tecnología para el mantenimiento y conservación de praderas de zacate buffel en México y los Estados Unidos; Fomento Ganadero; Sonora. 1990;(19):13–14. [ Links ]
62. Rodríguez, O, González–Domínguez J, Krausz JP, Odvody GN, Wilson JP, Hanna WW, Levy M. First report and epidemics of Buffelgrass blight caused by Pyricularia grisea in South Texas Plant Dis 1999;83:398. [ Links ]
63. Burton GW, Hart RH, Lowery RS. Improving forage quality in bermudagrass by breeding. Crop Sci 1967;(7):329–332. [ Links ]
64. Hanna WW, Hill RR. Forage crop breeding. In: Heath ME, Barnes RF, Metcalf DS editors. Forages the science of grassland agriculture. Ames, Iowa, USA: Iowa State University Press; 1985:80–88. [ Links ]
65. Burton GW, Monson WG. Inheritance of dry matter digestibility in bermudagrass. Cynodon dactylon (L.) Pers. Crop Sci 1972;(12):375–378. [ Links ]
66. Koltunow AM, Bicknell RA, Chaudhury AM. Apomixis: Molecular strategies for the generation of genetically identical seeds without fertilization. Plant Physiol 1995;(108):1345–1352. [ Links ]
67. Berthaud J. 2001. Apomixis and the management of genetic diversity. In: Savidan Y, Carman JG, Dresselhaus T editors. The flowering of apomixis: from mechanisms to genetic engineering. Mexico, DF. CIMMYT, IRD, European Comission DG VI (FAIR) 2001;8–23. [ Links ]
68. Combes D. Polymorphysme et modes de reproduction dans la section des Maximae du genre Panicum en Afrique. Mémoires ORSTOM 1975. [ Links ]

