










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de ciencias pecuarias
versión On-line ISSN 2448-6698versión impresa ISSN 2007-1124
Rev. mex. de cienc. pecuarias vol.1 no.1 Mérida ene./mar. 2010
Artículos
Efectos genéticos directos y maternos del peso al destete en una población de ganado Cebú de Cuba
Direct and maternal genetic effects for weaning weight in a Zebu beef cattle population in Cuba
Alejandro Palacios–Espinosaa, José Luis Espinoza–Villavicencioa, Danilo Guerra–Iglesiasb, Dianelys González–Peña Fundorab, Rafael de Luna de la Peñaa
a Departamento de Zootecnia, Universidad Autónoma de Baja California Sur, Carr. al Sur, km. 5.5, 23080. La Paz, BCS., México. Tel: (612)1238800, Ext 5010 y 5200. jlvilla@uabcs.mx Correspondencia al primer autor.
b Centro de Investigaciones para el Mejoramiento Animal de la Ganadería Tropical, La Habana, Cuba.
Recibido el 22 de abril de 2009
Aceptado para su publicación el 28 de julio de 2009
Resumen
El objetivo de este estudio fue estimar los parámetros genéticos para peso al destete en ganado Cebú. Se utilizaron datos de 20,151 terneros nacidos entre 1988 y 2006 en la provincia de Camagüey, Cuba. Se aplicaron seis modelos estadísticos que tuvieron como efecto fijo en común al grupo de contemporáneas (año–hato–época–sexo) y como covariables la edad al destete lineal y la edad de la madre al parto lineal y cuadrática. El efecto aleatorio del animal fue común a todos los modelos y se alternaron los efectos genético materno, del ambiente permanente y la covarianza genética entre los efectos directo y materno. De acuerdo con la prueba del logaritmo de la verosimilitud, el mejor modelo fue el que incluyó los efectos aleatorios del animal, maternos y del ambiente permanente, además, la covarianza entre los efectos directos y maternos. La heredabilidad de los efectos genéticos directo (0.06 a 0.34) y materno (0.06 a 0.19) varió entre los modelos. La correlación entre el efecto directo y materno varió entre –0.12 ± 0.13 y –0.19 ± 0.15. El efecto materno ambiental determinó un 13 % de la variabilidad total. Se concluyó que la heredabilidad del peso al destete en el ganado Cebú de la población estudiada puede ser sobre estimada cuando el modelo animal no incluye los efectos maternos; y no obstante que los valores de heredabilidad encontrados fueron bajos para los efectos directos y maternos, la ausencia relativa de antagonismo genético entre ellos podría permitir un mejoramiento genético moderado por selección.
Palabras clave: Ganado Cebú, Peso al destete, Parámetros genéticos, Efectos maternos.
Abstract
Genetic parameters for weaning weight were estimated in a Zebu beef cattle population in Camagüey province, Cuba. Data for 20,151 calves born between 1988 and 2006 were used in six variants of the statistical animal model. Contemporary groups (yearherd–season–sex) were used as a common fixed effect, and a weaning age linear regression and dam age linear and quadratic regressions as covariables. Animal random effect was common to all models, and models 2 to 6 had different combinations of genetic maternal effects, maternal permanent environmental effect and covariance between direct and maternal genetic effects. A log likelihood test showed the best model to be that which included animal random effects, maternal effects, covariance between direct and maternal effects and permanent environment. Heritability for direct (0.06 to 0.34) and maternal genetic effects (0.06 to 0.19) varied between models. Correlation between direct and maternal genetic effects varied from –0.12 ± 0.13 to –0.19 ± 0.15. The maternal permanent environmental effect explained 13 % of total variability. Heritability of weaning weight in the studied population can be overestimated when the animal model does not include maternal effects. The direct and maternal effects heritability values were low, but the relative absence of genetic antagonism between these effects indicates the possibility of moderate genetic improvement by selection.
Key words: Zebu cattle, Weaning weight, Genetic parameters, Maternal effects.
INTRODUCCION
El peso al destete es uno de los caracteres más estudiados en ganado de carne(1,2,3) y está altamente influido por los efectos maternales (genéticos y ambientales) que deben ser tenidos en cuenta de forma conjunta, especialmente cuando existe una relación antagónica entre ellos(4). Es ampliamente reconocido que los efectos genéticos indirectos pueden jugar un papel importante en la determinación de la respuesta fenotípica a la selección(5), en particular, la varianza genética materna, la cual se presenta cuando el fenotipo de un individuo es influenciado por el genotipo materno, independientemente de los efectos directos de los genes heredados(6).
En algunas razas cebuinas y sus cruces, los parámetros genéticos estimados para los efectos genéticos directos y maternos evidencian en la mayoría de los casos la influencia materna en los caracteres hasta el destete, así como un antagonismo genético entre efecto genético materno y el genético directo(7) y si no se tiene en cuenta el primero, se puede sobreestimar el valor genético de los animales(8,9). En presencia de efectos maternos, la respuesta a la selección está determinada por la heredabilidad total (h2T), la cual puede ser estimada como h2T = (VA + 1.5CovAM + 0.5VM)/VP(10). En esta ecuación, es evidente que cuando se incrementa la varianza genética materna y la covarianza de los efectos genéticos directo y materno, aumenta la respuesta a la selección, mientras que los valores negativos en dicha covarianza tienen el efecto contrario(5). La estimación del efecto materno sobre el peso al destete ha sido problemática, debido a que generalmente está confundido con el efecto genético directo. Con el incremento de la potencia de los equipos de cómputo, los modelos lineales empleados en la evaluación genética de los animales son cada día más complejos y el modelo animal ha sido hasta ahora el método más utilizado en la estimación de los efectos maternales en ganado de carne, basado principalmente en el modelo biométrico propuesto por Willham(11). Los modelos animal usados para analizar la influencia materna sobre el peso al destete de las crías generalmente incluyen los efectos genéticos directo y materno, así como la covarianza entre ellos y un efecto del ambiente permanente de la madre(12).
Considerando que los parámetros genéticos utilizados en los planes de mejoramiento son específicos para cada población de ganado bovino, el objetivo del presente trabajo fue estimar parámetros genéticos del peso al destete en una población de ganado Cebú cubano en la provincia de Camagüey, comparando seis variantes del modelo animal que incluían o no los efectos maternos y su relación genética con el efecto directo.
MATERIALES Y MÉTODOS
Se analizó información de ganado Cebú de la República de Cuba, situada en la entrada del Golfo de México, entre los 20° a 23° N y 74° a 85° O. Según la clasificación de Köppen (modificada), en la mayor parte de Cuba el clima predominante es del tipo cálido tropical.
Hay dos estaciones claramente definidas, la seca (invierno) de noviembre a abril, y la lluviosa (verano) de mayo a octubre. La temperatura media anual es 25° C, la media en invierno es 20 °C y en verano 26 a 27 °C. Las temperaturas mínimas varían entre 1 y 8.5 °C en el occidente del país y entre 3 y 12.5 °C en la parte oriental; las temperaturas máximas registradas están entre 36 y 38 °C. La humedad relativa promedio es 80 %. La precipitación media anual es 1,200 mm, con alrededor de 30 % en el período invernal y 70 % en el verano, y son más abundantes en el occidente que en el oriente del país(13).
El sistema de alimentación se basó principalmente en pastoreo sobre zacate estrella (Cynodon nlemfuensis), zacate Guinea (Panicum máximum) y sales minerales a libre demanda. En todos los hatos se utilizó la inseminación artificial después de la observación del estro natural durante todo el año. Los animales recibieron la vacuna contra leptospira, la vacuna recombinante contra la garrapata Boophilus microplus cada tres meses a partir del mes de nacido, septicemia hemorrágica y carbunco.
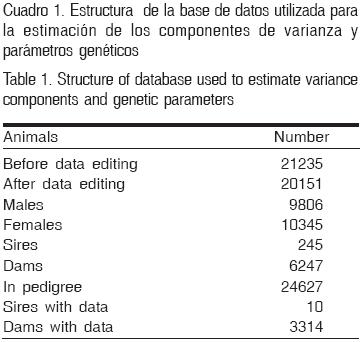
Se tomó una muestra inicial del peso al destete de 22,405 terneros que nacieron entre los años 1988 y 2006 en la Empresa Genética 'Rescate de Sanguily' de la provincia Camaguey, ubicada en la parte central de Cuba. Los datos fueron proporcionados por el Centro Nacional de Control Pecuario (CENCOP). En la edición de los datos se eliminaron aquellos grupos de contemporáneas (hato–año–trimestre de nacimiento–sexo de la cría) con menos de cinco observaciones. Se eliminaron los individuos con madre desconocida y todos aquellos a los que les faltara la información relacionada con la fecha de nacimiento, edad de la madre, hato, edad y peso al destete. La estructura de la base de datos utilizada se presenta en el Cuadro 1. Como puede observarse, la muestra quedó conformada por un total de 20,151 registros de peso al destete y 24,627 individuos en el pedigrí. Los terneros, cuya edad promedio fue de 202 días, fueron hijos de 245 padres y 6,247 madres entre 25 y 206 meses de edad que pertenecientes a 1,616 grupos de contemporáneas.
Los datos fueron analizados mediante el programa ASReml(14) aplicando seis modelos matemáticos. Como efectos fijos comunes a todos los modelos se consideraron el grupo de contemporáneos, la edad al destete del ternero como covariable lineal y la edad de la madre como covariable lineal y cuadrática.
El Modelo 1 incluyó el efecto genético directo como único efecto aleatorio.
El Modelo 2 incluyó el efecto genético directo, más el efecto del ambiente permanente atribuido a la madre y no correlacionado con los otros efectos en el modelo.
El Modelo 3 incluyó los efectos genéticos directo y materno, asumiendo una covarianza igual a cero entre ellos.
El Modelo 4 incluyó los efectos genéticos directo y materno, así como la covarianza entre ellos.
El modelo 5 incluyó el efecto genético directo, el efecto genético materno y el efecto del ambiente permanente de la madre, asumiendo una covarianza igual a cero entre los efectos directo y materno.
El modelo 6 incluyó el efecto genético directo, el efecto genético materno, el efecto del ambiente permanente de la madre y la covarianza entre los efectos directo y materno.
La notación matricial de los modelos es la siguiente:

Donde y es el vector de las observaciones del peso al destete, b es el vector de los efectos fijos que incluye el grupo de contemporáneas y la regresión del peso al destete en la edad de la cría lineal y la regresión del peso al destete en la edad de la madre lineal y cuadrática, a es el vector de los efectos aleatorios genéticos directos, m es el vector de los efectos aleatorios genéticos maternos, c es el vector de los efectos aleatorios del ambiente permanente, e es el vector de los efectos aleatorios residuales y X, Zd, Zm,y Zc son las matrices de incidencia que relacionan las observaciones con los efectos fijos, genéticos directos, genéticos maternos y del ambiente permanente, respectivamente.
El primero y segundo momento de los componentes aleatorios para el Modelo 6 que incluye todos los efectos son:

Donde A es la matriz de relaciones aditivas entre todos los individuos en el pedigrí, I es la matriz identidad, n es el número de madres, N es el número total de observaciones,  es la varianza genética aditiva de los efectos directos,
es la varianza genética aditiva de los efectos directos,  es la varianza genética aditiva de los efectos maternales, σam es la covarianza genética aditiva entre los efectos directos y los efectos maternales,
es la varianza genética aditiva de los efectos maternales, σam es la covarianza genética aditiva entre los efectos directos y los efectos maternales,  es la varianza de los efectos del ambiente permanente y
es la varianza de los efectos del ambiente permanente y  es la varianza del error y
es la varianza del error y 
Las heredabilidades para los efectos genéticos directos  , efectos genéticos maternales
, efectos genéticos maternales  y la correlación genética entre los efectos directos y maternales (ram) se estimaron con las siguientes ecuaciones:
y la correlación genética entre los efectos directos y maternales (ram) se estimaron con las siguientes ecuaciones:

Para determinar el modelo más apropiado se utilizó la prueba del logaritmo de la verosimilitud (15):

Donde log Lr y log Lc son los logaritmos de la verosimilitud para el modelo reducido y completo, respectivamente. En esta prueba, la distribución Jicuadrada tiene grados de libertad igual al número de parámetros en el modelo completo menos el número de parámetros en el modelo reducido.
RESULTADOS Y DISCUSIÓN
La edad promedio al destete de los terneros fue de 202 días y un peso de 158.5 ± 28.0 kg, (escala de 70 a 260 kg), con un coeficiente de variación de 17.6 %.
Los componentes de varianza y el logaritmo de la verosimilitud (log L) son presentados en el Cuadro 2. De acuerdo a la prueba del logaritmo de la verosimilitud los mejores modelos fueron aquéllos que incluyeron los efectos genéticos directos, los efectos genéticos maternales y los efectos del ambiente permanente (5 y 6) resultando estos similares entre sí (P>0.05). Sin embargo, el Modelo 6 que contempló además la covarianza entre los efectos directos y maternos, resultó numéricamente mejor que el Modelo 5. Los Modelos 3 y 4 resultaron similares entre ellos (P>0.05) y los componentes de varianza estimados por ambos son similares. El Modelo 1, en el que solamente se tuvo en cuenta el efecto aleatorio del animal, fue el que tuvo el ajuste más bajo y como se observa en el Cuadro 2, el componente de varianza genética aditiva para los efectos directos registró el valor más alto, superando en más de cuatro veces los valores obtenidos con los modelos que incluyeron los efectos maternales. Lo anterior coincide con otro estudio, en el cual se señala una sobreestimación de los componentes de co(varianza) para el peso cuando no se incluyen los efectos maternos en el modelo animal utilizado(16). En los modelos 2, 3, 4, 5 y 6 del presente trabajo, la suma de los componentes de la varianza sin incluir al error, presenta valores similares. La diferencia básica que existe entre los Modelos 3 y 4 con relación al 5 y 6 es que al incluir el efecto del ambiente permanente a los dos últimos, determinó una reducción en la varianza del error.
En el Cuadro 3 se presentan los parámetros genéticos estimados con los seis modelos aplicados y como se observa, la heredabilidad del efecto directo osciló entre 0.34 ± 0.02 en el modelo que solamente tuvo en cuenta el efecto aleatorio del animal (Modelo 1) y 0.06 ± 0.01 en los modelos 3, 4, 5 y 6 que incluyeron el efecto genético materno. Esta escala de valores coincide con lo reportado para ganado de carne en países tropicales(17) pero el estimado más bajo es inferior al valor reportado en ganado Hereford (0.18) cuando se utilizó un modelo animal que incluyó los efectos aleatorios del genotipo directo y materno, el ambiente permanente materno, el ambiente residual y los efectos fijos de año x sexo(1). Valores más altos aún (0.38) han sido reportados en ganado bovino para carne en Australia cuando se aplicó un modelo mixto multicarácter que incluyó el efecto materno(18). Ríos(19), utilizando 337 trabajos publicados entre los años 1,940 y 2,006 reporta para el peso al destete un valor promedio no ponderado para la heredabilidad directa de 0.27 en varias razas y de 0.26 para ganado Brahman y Nelore. La mayor heredabilidad encontrada en el Modelo 1 en el presente trabajo se debe posiblemente a una sobreestimación del componente aditivo (Cuadro 2) debido a que no incluye el efecto genético materno, por lo que debería de asumirse una heredabilidad baja para el efecto directo del peso al destete en esta población de ganado Cebú cubano. Resultados similares fueron reportados por otros autores(20).
Los valores de heredabilidad para el efecto materno (Cuadro 3) fueron menores (0.06 ± 0.01) cuando se estimaron con los Modelos 5 y 6 que cuando se aplicaron los Modelos 3 (0.18 ± 0.01) y 4 (0.19 ± 0.02). Esta diferencia probablemente se debe a que en los últimos no fue incluido el efecto del ambiente permanente de la madre, y el efecto genético materno está sobreestimado debido a su confusión con el efecto del ambiente materno. Una heredabilidad del efecto materno similar (0.08) a la obtenida con el Modelo 6 fue estimada en ganado Limousin y Gelbvieh(21) con un modelo animal que incluyó los mismos efectos. Valores inferiores aún (0.02) han sido reportados en ganado de carne de Australia(18). Sin embargo, en otros estudios que incluyeron los mismos efectos aleatorios que en el Modelo 6 del presente trabajo, la heredabilidad del efecto materno fue de 0.18(1) y de 0.20(16). En la revisión realizada por Ríos(19), se reporta un valor promedio no ponderado de 0.17 para la heredabilidad del efecto materno. Dicho valor es superior al encontrado en el presente trabajo con los modelos de mejor ajuste (5 y 6). De manera general, la heredabilidad del efecto materno resulta inferior que la estimada para el efecto directo(19,22). En el presente trabajo, los modelos de mejor ajuste estimaron un valor de heredabilidad igual (0.06±0.01) para ambos efectos genéticos (directo y materno).
Respecto al efecto del ambiente permanente, en el presente trabajo se estimó un valor de 0.13 ± 0.01 con los modelos 5 y 6. Este valor es similar a la media no ponderada (0.14) referida en varias razas de bovinos para carne(19) en ganado Brahman de Venezuela(23) pero ligeramente superior al publicado por Mercadante y Lobo(17) en un hato Nelore en Brasil (0.10). El efecto del ambiente permanente explica entre el 2 y el 29 % de la varianza del peso al destete en las diferentes poblaciones de bovinos para carne(22).
La correlación genética entre los efectos genéticos directo y materno fue de –0.12 ± 0.13 en el modelo 4 y de –0.19 ± 0.15 en el Modelo 6 (Cuadro 3). Si se observan los valores de los componentes de varianza y el logaritmo de la verosimilitud en el Cuadro 2 se puede ver que la inclusión de la covarianza entre los efectos genéticos directo y materno no determinó cambios significativos en el modelo (3 vs 4 y 5 vs 6), lo que pone en evidencia la poca importancia de dicha covarianza, al menos en el caso particular de esta población. Correlaciones genéticas con valores cercanos a los estimados con el Modelo 6 fueron reportadas en ganado Hereford (–0.22)(1) y en Limousin y Gelbvieh (–0.17 a –0.25)(21) cuando se utilizaron modelos que incluyeron los mismos efectos aleatorios.
La mayoría de los reportes en la literatura presentan correlaciones antagónicas y altas entre los efectos genéticos directo y materno para el peso al destete del ganado bovino para carne(19,24). Esta correlación genética negativa ha sido atribuida a una covarianza ambiental negativa entre madre y descendientes, que se asume igual a cero en el modelo de Willham(11). Esta covarianza ambiental entre madres e hijas ha sido llamada el síndrome de las ubres grasas y plantea que las madres con una habilidad materna superior pueden sobrealimentar a sus hijas, consecuentemente, el desarrollo de tejido mamario en ésta se inhibe de tal manera que su propia habilidad materna se expresa por debajo del promedio(25). Es probable también que la covarianza negativa de los efectos genéticos directos y maternos pudiera presentarse para genes que tienen efectos pleiotrópicos antagónicos sobre el comportamiento materno de las vacas y el comportamiento de sus terneros(26).
Los resultados obtenidos por Van Vleck et al(27) pusieron de manifiesto que el antagonismo genético entre los efectos genéticos directo y materno afecta la eficiencia de la selección a largo plazo. Sin embargo, a pesar de los bajos valores de heredabilidad encontrados en el presente estudio para los efectos genéticos directo y materno, la ausencia aparente de un antagonismo genético entre estos rasgos podría permitir una respuesta a la selección moderada en esta población.
CONCLUSIONES E IMPLICACIONES
Se concluye que la heredabilidad del peso al destete en el ganado Cebú de la población estudiada puede ser sobreestimada cuando el modelo animal no incluye los efectos maternos y aún cuando los valores de heredabilidad encontrados fueron bajos para los efectos directos y maternos, la ausencia relativa de antagonismo genético entre ellos podría permitir un mejoramiento genético moderado por selección.
LITERATURA CITADA
1. Dodenhoff J, Van Vleck LD, Kachman SD, Koch RM. Parameter estimates for direct, maternal, and grandmaternal genetic effects for birth weight and weaning weight in Hereford cattle. J Anim Sci 1998;76(10):2521–2527. [ Links ]
2. Dodenhoff J, Van Vleck LD, Wilson DE. Comparison of models to estimate genetic effects for weaning weight of Angus cattle. J Anim Sci 1999;77(12):3176–3184. [ Links ]
3. Bijma P. Estimating maternal genetic effects in livestock. J Anim Sci 2006;84(4):800–806. [ Links ]
4. Bertrand JK, Benyshek LL. Variance and covariance estimates for maternally influenced beef growth traits. J Anim Sci 1987;64(3):728–734. [ Links ]
5. Wolf JB, Brodie III ED, Cheverud JM, Moore AJ, Wade MJ. Evolutionary consequences of indirect genetic effects. Trends in Ecology & Evolution 1998;13(2):64–69. [ Links ]
6. Mousseau TA, Fox CW. Maternal effects as adaptations. Oxford: Oxford University Press; 1998. [ Links ]
7. Lee C, Pollak EJ. Genetic antagonism between body weight and milk production in beef cattle. J Anim Sci 2002;80(2):316– 321. [ Links ]
8. Ribeiro PMT, Ferraz JBS, Eler JP. Parámetros genéticos e nivel de endogamia em bovinos da raza Santa Gertrudis no Brasil. Arq Bras Med Vet Zootec 2000;52(6):641–646. [ Links ]
9. Ferraz FBP, Ramos AA, Da Silva CLO, De Sousa JC, De Alencar MM. Tendencia genética dos efeitos direto e materno sobre os pesos a desmama e pos desmama de bovinos de raca Tabapua no Brazil. Rev Braz Zootec 2002;31(2):635–640. [ Links ]
10. Willham RL. The role of maternal effects in animal breeding. III. Biometrical aspects of maternal effects in animals. J Anim Sci 1972;35(6):1288–1293. [ Links ]
11. Willham RL. The covariance between relatives for characters composed of components contributed by related individuals. Biometrics 1963;19(1):18–27. [ Links ]
12. Robinson DL. Estimation and interpretation of direct and maternal genetic parameters for weights of Australian Anguscattle. Livest Prod Sci 1996;45(1):1–11. [ Links ]
13. IMRC. Instituto Meteorológico de la República de Cuba [en línea]. http://www.met.inf.cu/asp/genesis.asp?TB0=PLANTILLAS&TB1=INICIAL. Consultado 4 de octubre de 2006. [ Links ]
14. Gilmour AR, Cullis BR, Welham SJ, Thompson R. ASREML Reference manual. Australia: NSW, Agriculture Biometric. 2000. [ Links ]
15. Meyer K. Variance components due to direct and maternal effects for growth traits of Australian beef cattle. Livest Prod Sci 1992;31(3–4):179–204. [ Links ]
16. Mourão GB, Ferraz JBS, Eler JP, Balieiro JCC, Bueno RS, Mattos EC, Figueiredo LGG. Genetic parameters for growth traits of a Brazilian Bos taurus x Bos indicus beef composite. Genet Mol Res 2007;6(4):1190–1200. [ Links ]
17. Mercadante MEZ, Lobo RB, de los Reyes A. Parámetros genéticos para características de crecimiento en cebuinos de carne. Arch Latinoam Prod Anim 1995;3(1):45–89. [ Links ]
18. Montaldo HH, Kinghorn BP. Efectos genéticos aditivos y no aditivos, directos y maternos para caracteres de crecimiento en una población multirracial de bovinos productores de carne. Arch Med Vet 2003;35(2):243–248. [ Links ]
19. Ríos A. Estimadores de parámetros genéticos para características de crecimiento predestete de bovinos. Revisión. Tec Pecu Mex 2008;46(1):37–67. [ Links ]
20. Ríos Utrera A, Martínez Velázquez G, Tsuruta S, Bertrand JK, Vega Murillo VE, Montaño Bermúdez M. Estimadores de parámetros genéticos para características de crecimiento de ganado Charolais mexicano. Tec Pecu Mex 2007;45(2):121– 130. [ Links ]
21. Iwaisaki H, Tsuruta S, Misztal I, Bertrand JK. Estimation of correlation between maternal permanent environmental effects of related dams in beef cattle. J Anim Sci 2005;83(3):537–542. [ Links ]
22. Quintanilla R, Piedrafita J. Efectos maternos en el peso al destete del ganado vacuno de carne: Una revisión. ITEA 2000;96(1):7–39. [ Links ]
23. Plasse D, Verde O, Fossi H, Romero R, Hoogesteijn R, Bastidas P, Bastardo J. (Co)variance components, genetic parameters and annual trends for calf weights in a pedigree Brahman herd under selection for three decades. J Anim Breed Genet 2002;119(3):141–153. [ Links ]
24. Mercadante MEZ, Lobo RB. Estimativas de (co)variancias e parámetros genéticos dos efeitos direto e materno de características de crescimento de femeas de un rebanho Nelore. R Bras Zootec 1997;26(6):1124–1133. [ Links ]
25. Totusek R, Stephens OF, Kropp JR, Holloway SW, Knosi JW, Whiteman JV. Milk production of range cows. Okla Agric Ext Sta. Misc Pub 85. 1971. [ Links ]
26. Wilson AJ, Réale D. Ontogeny of additive and maternal genetic effects: lessons from domestic mammals. Am Nat 2006;167(1):E23–E38. [ Links ]
27. Van Vleck LD, St. Louis D, Miller JI. Expected phenotypic response in weaning weight of beef calves from selection for direct and maternal genetic effects. J Anim Sci 1977;44(3):360–367. [ Links ]

