











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
El pak choi (Brassica rapa subsp. Chinensis) es una planta herbácea que pertenece al género brassica y comparte especie con otras plantas o cultivares que se encuentran en México. El pak choi se cultiva en Asia y Oceanía, principalmente, donde se considera uno de los vegetales de mayor importancia (Qing et al., 2000). Las brasicáceas son un conjunto de plantas que incluye a las coles, el kale, el brócoli, el nabo y la coliflor. El pak choi se clasifica de manera generalizada con otras brasicáceas en FAOSTAT con el código 0358. Existe poca información acerca del cultivo de esta planta fuera de los países donde se produce mayormente. Sin embargo, su distribución en otros países y continentes se debe principalmente a los inmigrantes asiáticos, además, el consumo de pak choi se ha incrementado con la tendencia actual de consumir alimentos con alto valor nutricional.
En México se produce para satisfacer la demanda local y de exportación a Estados Unidos de América, Canadá y Japón. Esta se lleva a cabo en Baja California, Sonora, Aguascalientes y Guanajuato, siendo este último estado el que reporta cifras oficiales. En el año 2021 hubo una producción reportada de 20.73 t en el estado de Guanajuato, con un rendimiento de 24.56 t ha-1 a un precio de $5 850.00 MXN por tonelada, según la información del Servicio de Información Agroalimentaria y Pesquera (SIAP). El cultivo de pak choi requiere un suelo franco de buen drenaje, con un pH de 5.8 a 7.5. Los requerimientos nutricionales son de 55-75 kg ha-1 N, 40-80 kg ha-1 P and 80-110 kg ha-1 K al plantar, riego 3 veces a la semana y una población de 50 000 plantas ha−1 (Maseko et al., 2017).
Cerca de 80% de las bacterias presentes en la rizosfera son capaces de producir AIA (Vega-Celedón, 2016), estos microorganismos interactúan con las plantas por medio de las raíces y pueden regular el desarrollo de estas mediante un desbalance de auxinas, afectando a la planta de forma positiva o negativa. Por otra parte, las bacterias del suelo también son capaces de solubilizar fosfatos, formar compuestos quelatados, fijar nitrógeno atmosférico, lo que significa un beneficio para las plantas. Estos microorganismos son conocidos como rizobacterias promotoras del crecimiento vegetal (RPCV).
El uso de RPCV es una alternativa para promover el crecimiento de las plantas a través de producción de reguladores de crecimiento, la solubilización de nutrientes del suelo, tolerancia al estrés biótico y abiótico, biopesticida y estimulación de las defensas de las plantas (Tsukanova et al., 2017; Leontidou et al., 2020). De esta manera, se puede mejorar el rendimiento de las áreas cultivables al hacer más eficiente el uso de fertilizantes, esto debido al elevado costo que representan en la actualidad. Además, se pueden mitigar los daños al suelo por los fertilizantes químicos (Basu et al., 2021). El objetivo de este trabajo fue evaluar el efecto de la inoculación de tres cepas de RPCV en el que los parámetros de crecimiento y contenido de N, P, K y S de plantas de pak choi en macetas y condiciones de macro-túnel.
Materiales y métodos
El experimento se estableció en el colegio de Postgraduados campus Montecillo (19° 27’ 38.975 latitud norte, 98° 54’ 14.84 longitud oeste y 2 250 m) en el mes de febrero de 2021, con una temperatura promedio de 24 °C y humedad relativa de 56%. Se sembraron semillas de pak choi (Brassica rapa subespecie Chinensis L.) variedad Cantón con un porcentaje de germinación declarado de 96% (Harley Seeds, Reino Unido) y un porcentaje de germinación experimental de 94%. Estas fueron colocadas en un semillero de 200 cavidades que previamente se llenaron con una mezcla de perlita y peatmoss (1:1). Al emerger las plántulas éstas se regaron con solución Steiner al 50%. Las plántulas de 35 días de edad se trasplantaron a bolsas que contenían 5 kg de suelo tipo franco arcillo-arenoso de textura media, libre de carbonatos, pH de 7.92. El experimento se basa en un diseño completamente al azar con 10 repeticiones para cada tratamiento y para el testigo, respectivamente.
Se utilizaron tres de cepas de rizobacterias promotoras del crecimiento vegetal el Laboratorio de Microbiología de Suelos del Colegio de Postgraduados: Arthrobacter pokkalii JLB4 (código de acceso GenBank: MW629814.1) aislada de suelo de la rizósfera de Solanum lycopersicum, Pseudomonas tolaasii P61 (código de acceso GenBank: KY933651.1) y Pseudomonas tolaasii A46 (código de acceso GenBank: KY933652.1) aisladas de la rizósfera de Solanum tuberosum. El inóculo se preparó sembrando las cepas en caldo nutritivo, incubándose a 28 °C y con agitación a 140 rpm por 72 h. Se ajustó la densidad de inóculo a 108 UFC mL-1 para cada cepa, cuantificando el número de células en la cámara de Neubauer.
Se inocularon 10 plantas de pak choi con 5 mL de inóculo, los testigos fueron inoculados únicamente con caldo nutritivo estéril, este proceso se repitió a los 32 días siguiendo el mismo procedimiento. Las plantas fueron regadas cada tercer día con agua. Todas las plantas fueron fertilizadas con una dosis de 164-53-187 de NPK, una inmediatamente al realizar el trasplante y la primera inoculación, la siguiente posterior a la segunda inoculación.
Pasados tres meses se cosecharon las plantas para evaluar el efecto de las rizobacterias. Se determinó el volumen radical por el método de volumen de agua desplazado en una probeta graduada. El material se secó en una estufa de aire forzado a 70 °C por 72 h y después se determinó el peso seco de la biomasa de la planta. Se evaluó el contenido de nitrógeno (N), potasio (K), fósforo (P) y azufre (S) en la planta, tomando muestras de 0.5 g de material seco y molido de la parte aérea y las raíces de las plantas. Para la digestión de nitrógeno, se utilizó una mezcla de H2SO4 y HClO4 en proporción 2:1, más H2O2 al 30%. Para la digestión de K, P y S, se utilizó HNO3 y HClO4 en proporción 2:1. Tanto las digestiones como las determinaciones se realizaron de acuerdo con el método de Alcántar y Sandoval (1999), con un equipo de espectrometría de emisión óptica de inducción por plasma acoplado (ICP-OES 725-ES, Agilent, Santa Clara, CA, USA).
Los datos obtenidos se analizaron con el programa estadístico SAS versión 9.0, siendo un análisis de varianza con una prueba post-hoc de comparación de medias de Tukey para los parámetros de peso seco de hojas, peso seco total y contenido de N, P, K y S. Se realizó un análisis para datos no paramétricos (comparación de Kruskall-Wallis) para volumen radical y peso seco de raíz respectivamente, con una prueba post-hoc de comparación de medias de Dunn, con un intervalo de confianza de 95%.
Resultados y discusión
En la Figura 1 se muestran parámetros agronómicos relacionados con la interacción de las plantas con RPCV. En la sección 1a se realiza una comparación del volumen radical y el peso seco de la raíz de los diferentes tratamientos con el testigo, en ella se observa que fueron las plantas inoculadas con la cepa JLB4 (A. pokkalii) las que tuvieron un mayor peso seco, siendo 8.51 g un valor entre 30 y 47% mayor que los demás tratamientos. Para el volumen radical, también fue mayor, siendo 75 ml con una diferencia entre 35 y 51% respecto a los demás tratamientos. En la sección 1b los resultados del peso seco total en las hojas de pak choi para cada tratamiento con RPCV y el testigo, sin una diferencia estadísticamente significativa (p= 0.0562).

Figura 1 Parámetros agronómicos en plantas de pak choi con tratamiento de RPCV. a) volumen radical y peso seco de raíces; y b) peso seco de hojas. Letras distintas indican diferencias significativas entre grupos.
El aumento en el volumen de la raíz en las plantas inoculadas puede estar relacionado con un mecanismo directo de promoción de crecimiento, como es el caso de la producción de ácido 3-indol -acético (Grover et al., 2020). Las auxinas son moléculas que participan activamente en todas las etapas fenológicas de las plantas, a nivel celular intervienen en la elongación, división y diferenciación celular (Ljumg, 2013). Debido a que esta hormona se distribuye de forma no uniforme en los tejidos de las plantas y esto afecta los procesos morfogenéticos se le conoce como un ‘morfógeno’ (Vanneste y Friml, 2009). Se ha reconocido la capacidad de las bacterias del género Pseudomonas y de A. pokkalii, entre otras, para producir auxinas, las cuales influyen en el desarrollo y arquitectura de la raíz (Malik y Sindhu, 2011; Krishnan et al., 2016).
Un sistema radical más abundante beneficia al anclaje y soporte de la planta al suelo, una mayor superficie de contacto permite que la planta explore el suelo, absororba agua y nutrientes, así como una mayor interacción con los microorganismos del suelo. Los resultados de las interacciones entre suelo-planta-microorganismo resultan complejas y van a diferir dependiendo de factores como el tipo de suelo, su localización geográfica, la etapa fenológica y salud de la planta (Gomes et al., 2018).
Las plantas inoculadas con la cepa P61, presentaron el mayor peso seco total de todos los tratamientos (Figura 1b), con un porcentaje de 11.5% a diferencia del testigo. A pesar de que produjo menor desarrollo de la raíz, al final permitió un mayor desarrollo de la biomasa total, el cual es un efecto deseable en vegetales cuya parte aérea es consumida, como el caso del pak choi. Para los elementos evaluados en las plantas, en el caso del nitrógeno no hay diferencias significativas entre los tratamientos y el testigo (p= 0.4265) (Figura 2a).
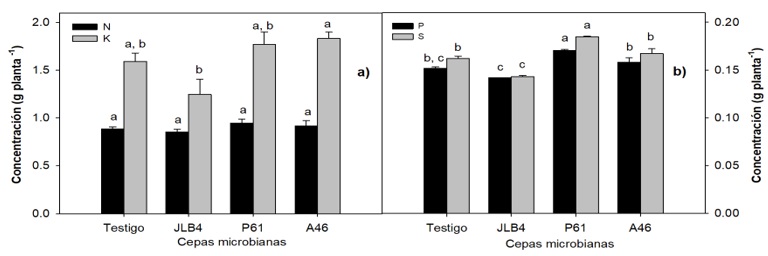
Figura 2 Contenido nutrimental en plantas de pak choi con tratamiento de RPCV, son: a) nitrógeno (N) y potasio (K), b) fósforo (P) y azufre (S). Letras distintas indican diferencias significativas entre grupos.
El nitrógeno es un elemento importante en el desarrollo de las plantas, una deficiencia de N puede provocar proteólisis en las hojas, degradación de los cloroplastos y disminución del contenido de clorofila (Almeida et al., 2020). De acuerdo con los resultados obtenidos (Figura 3a), no existen diferencias significativas en el contenido de este elemento, lo que sugiere que no existe una limitación en la disponibilidad de este elemento. El potasio es un catión con una alta movilidad en las plantas y con funciones fisiológicas relevantes, como es el movimiento del agua, nutrientes y carbohidratos dentro de la planta (Xu et al., 2020). Se puede observar en la figura 3a que, aunque la cepa A46 es la que acumuló una mayor cantidad de K (p= 0.0299), en la prueba post-hoc no se aprecian diferencias entre los grupos.
Una característica destacable de los glucosinolatos implica beneficios para la salud cuando son destinadas para el consumo humano y que son parte de todas las brasicáceas (Johnson et al., 2016). Estos metabolitos tienen la función principal en la planta como defensa contra infecciones y herbivoría (Ishida et al., 2014). Zang et al. (2015); Blažević et al. (2020) y mencionan la síntesis de glucosinolatos, la cual se lleva a cabo en las plantas a partir de aminoácidos que contienen azufre, como metionina y tirosina.
Se puede observar en la Figura 2b que el tratamiento con la cepa P61 produjo la mayor concentración de este elemento en las plantas (p< 0.0001) siendo de 0.1847 ± 0.004 g planta-1, 12.3% superior con respecto al testigo. Aunque el potasio y fósforo son abundantes en el suelo, éstos se encuentran unidos a otros minerales y esto provoca que entre 95 y 99% de estos elementos se encuentren en formas insolubles y no disponibles para las plantas (Wang et al., 2020). Las especies del género Brassica pueden tolerar suelos con baja disponibilidad de fósforo (P) con menores efectos adversos que otros cultivos con mayores requerimientos (Marschner et al., 2007). Sin embargo, la inoculación de RPCV promueve la solubilización de P y su disponibilidad para la planta (Figura 2b).
P. tolaasii P61 produjo un aumento en el contenido de fósforo, siendo de 0.1704 ± 0.001 g planta-1, el cual fue 10.8% mayor comparado con el testigo (p= 0.0003). De acuerdo con Castagno et al. (2021), P. tolaasii es una bacteria con la capacidad de promover la absorción del fósforo en las plantas, al igual que A. pokkalii (Alexander et al., 2019). Esto se lleva a cabo mediante alguno o ambos de los siguientes mecanismos para solubilizar fosfato: producción de ácidos de bajo peso molecular como el cítrico o el glucónico para el caso de fosfatos inorgánicos o mediante la síntesis de fosfatasas como fitasas o nucleasas para fosfatos orgánicos (Lobo et al., 2019).
Conclusiones
Para el caso de pak choi, el uso de las cepas P. tolaasii P61, A. pokkalii JLB4 y P. tolaasii A46 en combinación con fertilizante condujo a una mayor acumulación de biomasa, volumen radical y P, K, S en comparación con la sola aplicación de fertilizante. La mejor cepa fue P. tolaasii P61, la cual puede ser utilizada para aumentar la biomasa de hojas de pak choi y su contenido nutrimental.

