











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
La obtención de plantas doble haploides (DH) a partir de una célula germinal es una de las herramientas biotecnológicas que forma parte de programas de mejoramiento de cultivos alrededor del mundo, y establece la posibilidad de reducir los tiempos para obtener una planta homocigota. Otras ventajas de usar DH incluyen hacer la selección más eficiente, de caracteres cualitativos y cuantitativos, ya que se facilita la fijación de alelos recesivos benéficos, facilita la recuperación de recombinaciones únicas y se eliminan los efectos de la dominancia y la segregación dentro de las familias. En especies de polinización libre, los DH pueden ser utilizados como líneas parentales de una variedad sintética y son útiles para estimar los valores aditivos o la aptitud combinatoria general si son utilizados en cruzas con otros materiales de interés (Snape, 1989).
El chile (Capsicum annuum L.) es una hortaliza muy importante a nivel económico por su valor comercial a nivel internacional. En México, el chile crece en ambientes tanto tropicales como templados y su diversidad genética fundamenta la gran variabilidad de tipos de chile, así como su valor biológico, culinario e industrial. La androgénesis es el método más ampliamente utilizado para la obtención de haploides de C. annuum.
En la androgénesis un gametofito inmaduro es desviado de su desarrollo normal y es inducido a su desarrollo esporofítico (Jha et al., 2021). La obtención de plantas haploides de chile basada en el cultivo de anteras o microsporas (precursores del polen), es el método más reportado para la inducción embriogénica a partir de células gametofíticas masculinas. Las experiencias hasta ahora publicadas incluyen a las especies cultivadas de C. annuum (Comlekcioglu y Ellialtıoğlu, 2018), así como especies silvestres y cruzas interespecíficas (Munyon et al., 1989). Sin embargo, es a partir de la publicación de Dumas et al. (1981) que el protocolo para obtención de plantas doble haploide de chile ha venido desarrollándose para incrementar la eficiencia de inducción androgénica y reconversión a plantas completas de diferentes genotipos de chile.
Varios estudios han demostrado la alta frecuencia de embriones derivados de anteras; sin embargo, pocos de ellos fueron capaces de desarrollar plantas normales (Supena y Custers, 2011). Grozeva et al. (2009) establecieron que la formación de embriones oscilaba entre 0 y 9% de los cuales obtuvieron ocho plantas completas. Por su parte, Ercan y Ayar (2011) reportaron resultados similares los cuales variaron de 0 a 7.69% de formación de embriones; sin embargo, únicamente obtuvieron 12 plantas completas. El efecto de edad de la planta donante del explante (Kristiansen y Andersen, 1993), las condiciones de crecimiento (Büyükalaca et al., 2004), la temporada de crecimiento (Ata et al., 2019), los reguladores de crecimiento (Nowaczyk et al., 2016) y el genotipo de la planta donante afectan la eficiencia y éxito de la embriogénesis y ulterior desarrollo a planta completa; sin embargo, es el genotipo el factor principal que define la embriogénesis exitosa de microsporas a partir del cultivo de anteras (Niklas et al., 2012; Keles et al., 2015).
Uno de los principales objetivos de mejoramiento en chile de tipo mulato y ancho en el Programa de Mejoramiento de Chile en el Campo Experimental Bajío del INIFAP es obtener líneas con resistencia a Phytophthora capsici Leo. y a los geminivirus PepGMV y PHYVV, así como líneas ‘B’ o mantenedoras de esterilidad masculina citoplasmática. Avances importantes en la selección de materiales resistentes fueron obtenidos por lo que se hace necesario obtener líneas homocigotas de genotipos élite para la generación de híbridos comerciales.
Por lo anterior, el objetivo de este trabajo fue implementar y desarrollar un protocolo que permita regenerar plantas haploides y doble haploides en genotipos avanzados de chile mulato y ancho vía cultivo de anteras, y caracterizar la respuesta, eficiencia de inducción y obtención de plantas.
Materiales y métodos
Material vegetal
Se emplearon 27 genotipos experimentales de chile tipo mulato y ancho provenientes de dos poblaciones principales: a) población criolla heterogénea de chiles mulato y ancho con característica de esterilidad masculina citoplasmática (CMS), por sus siglas en inglés; b) población de cruzas interespecíficas (C. annuum x C. chinense) resistentes a geminivirus (PepGMV) y Phytophthora sp.; y c) variedades de chile comerciales ancho San Luis y ancho 3015, susceptibles a geminivirus (PepGMV) y Phytophthora sp. Las plantas donantes de los explantes experimentales, fueron sembradas en invernadero las cuales se les aplicó riego cada tres días por sistema de goteo y fertilizante foliar al inicio de floración y producción con Maxi-Grow Excel® 20 ml 25 L-1.
Colecta de botones florales
Los botones florales fueron colectados de las plantas de chile durante los meses de otoño e invierno boreal de 2019-2020. La etapa de colecta de los botones florales de chiles fue cuando éstos tenían aproximadamente un tamaño 6 a 6.8 mm y los pétalos aproximadamente 1 mm más largos que los sépalos, esto es cinco días a partir de la etapa fenológica de 50% de floración. Los botones florales se cortaron con una tijera desinfectada y fueron depositados en tubos Falcon® de 50 ml con agua destilada estéril.
Desinfección de botones florales
El agua donde se recolectaron los botones florales se desechó de los tubos Falcon( y se añadieron 30 ml de etanol al 70% en agitación (100 rpm) durante dos minutos. Al término de este tiempo se decantó el etanol y se añadieron 30 ml de cloro comercial al 10% (Cloralex(; 6% de cloro activo), permaneciendo en agitación durante 10 min. Finalmente, la solución de cloro se decantó y se realizaron tres lavados con agua destilada esterilizada, de 1 minuto cada uno. Después del proceso de desinfestación, los botones florales se mantuvieron en baño de hielo o en el refrigerador a aproximadamente 4 °C, hasta el momento del aislamiento de las anteras.
Cultivo de anteras en medio de inducción
Los botones florales desinfestados se colocaron sobre una caja de Petri esterilizada para extraer las anteras con la ayuda de dos pinzas quirúrgicas estériles. Los sépalos se separaron de los pétalos cuidadosamente para mantener la integridad de las anteras y éstas se disectaron con bisturí. Las anteras se cultivaron del lado convexo en contacto con el medio de inducción macroelementos: (NH4)2SO4 0.034 g L-1, NH4NO3 1.238 g L-1, CaCl2.2H2O 0.313 g L-1, KH2PO4 0.142 g L-1, Ca (NO3)2.4H2O 0.05 g L-1, Mg SO4.7H2O 0.412 g L-1, KNO3 2.15 g L-1, NaH2PO4.H2O, 0.038 g L-1, KCl 0.007 g L-1; microelementos: H3BO3 3.15 mg L-1, CoCl2.6H2O (stock 2.5 mg ml-1) 0.016 mg L-1, CuSO4.5H2O (stock 2.5 mg ml-1) 0.016 mg L-1, MnSO4.H2O 22.13 mg L-1, NaMoO4.2H2O 0.19 mg L-1, KI 0.7 mg L-1, ZnSO4.7H2O 3.625 mg L-1 (Dumas de Vaulx et al., 1981), suplementado con 0.01 mg L-1 de cinetina y 0.01 mg L-1 de ácido 2,4-diclorofenoxiacético (2,4-D), sacarosa 30 g L-1, vitaminas: tiamina 2 mg L-1, piridoxina HCl 1 mg L-1, ácido nicotínico 1 mg L-1, pantotenato de calcio 1 mg L-1, glicina 2 mg L-1, ácido fólico 1 mg L-1, biotina 1 mg L-1, mio-inositol 10 g L-1 y agar-agar 8 g L-1. El pH se ajustó a 5.9 previo a su esterilización a 121 °C y 15 libras sobre pulgada cuadrada durante 20 min, el medio fue vertido en cajas Petri esterilizadas de 90 ( 15 mm.
Las anteras cultivadas fueron seleccionadas por tener un tamaño aproximado de 2.5 a 3 mm de longitud. Las cajas Petri se sellaron con cinta Micropore( y se incubaron a 32 °C en oscuridad durante ocho días. Despues, las cajas Petri con anteras se llevaron a una cámara de crecimiento a 25 °C con un fotoperiodo de 16 h luz a una intensidad lumínica de 45-70 μmol·m-2 s-1 durante cuatro días en el mismo medio de inducción.
Cultivo de anteras en medio de germinación
Las anteras fueron transferidas a medio de germinación con los mismos macroelementos y concentración que el medio de inducción y microelementos en concentraciones como sigue: H3BO3 1.55 mg L-1, CoCl2.6H2O (stock 2.5 mg ml-1) 0.011 mg L-1, CuSO4.5H2O (stock 2.5 mg ml-1) 0.011 mg L-1, MnSO4.H2O 20.13 mg L-1, NaMoO4.2H2O 0.14 mg L-1, KI 0.33 mg L-1, ZnSO4.7H2O 3.255 mg L-1 (Dumas de Vaulx et al., 1981), adicionado con cinetina 0.1 mg L-1, sacarosa 30 g L-1, vitaminas (ver medio de inducción) y agar-agar (8 g L-1), el pH se ajustó a 5.9 previo a su esterilización. Las cajas Petri con las anteras se incubaron bajo las mismas condiciones de fotoperiodo y temperatura antes mencionadas.
Aislamiento y desarrollo de los embriones
Una vez que los embriones germinaron y mostraron sus polos apical y radicular, fueron transferidos a medio de elongación y enraizamiento libre de hormonas (Chambonnet, 1988) Macroelementos: KNO3 1.9 g L-1, NH4NO3 1.65 g L-1, MgSO4.7H2O 0.37 g L-1, CaCl2.2H2O 0.44 g L-1, KH2PO4 0.17 g L-1, microelementos: MnSO4.H2O 0.076 mg L-1, ZnSO4.7H2O 1 mg L-1, H3BO3 1 mg L-1, KI 0.01 mg L-1, CuSO4.5H2O 0.03 mg L-1, AlCl3.6H2O 0.05 mg L-1, NiCl2.6H2O 0.03 mg L-1 suplementado con sacarosa 30 g L-1, mio-inositol 10 g L-1, FeSO4.7H2O 18.65 mg L-1, Na2-EDTA 13.9 mg L-1 y agar-agar 8 g L-1. El pH se ajustó a 5.9 previo a su esterilización.
En una primera etapa de elongación, los embriones se transfirieron a cajas Petri 90 x 25 mm y conforme aumentaron su tamaño y la necesidad de espacio, se transfirieron a frascos tipo Gerber( de 170 ml Los frascos se sellaron con cinta micropore( e incubaron en la cámara de crecimiento a 25 °C con un fotoperiodo de 16 h luz, a una intensidad lumínica de 45-70 μmol m-2 s-1, hasta que las plántulas formaron cuatro pares de hojas verdaderas y raíces secundarias.
Determinación del nivel de ploidía
Ápices radiculares de dos raíces de 1 a 2 cm se disectaron y analizaron por medio de tinción de ploidía con la técnica de squash (Ahloowalia, 1965). Las raíces se colocaron en una solución de ácido acético (45%) y se incubaron a temperatura ambiente durante tres horas en agitación suave (100 rpm). Posteriormente, las raíces se transfirieron a una solución de etanol al 70% en agitación por dos horas y se refrigeraron a 4 °C hasta su análisis.
Las raíces almacenadas en alcohol fueron transferidas a un microtubo de 1.5 ml con 600 µl del inhibidor 8-hidroxiquinolina al 0.002 M por cuatro horas en oscuridad a temperatura ambiente (Valladolid et al., 2004). Transcurridas las cuatro horas de pretratamiento, se eliminó el inhibidor y se adicionaron 600 µl de la solución de Farmer (etanol absoluto + ácido acético glacial en proporción 3:1). Las muestras se incubaron durante 24 h a 4 °C. Se retiró la solución de Farmer con micropipeta y se realizó una hidrólisis ácida con HCl al 1 N por 15 min a 60 °C y posteriormente una hidrólisis enzimática con celulosa al 0.2% por 15 min a 37 °C. Se retiró la última solución de hidrólisis con la micropipeta y se realizaron dos lavados adicionando 600 µl de la solución EDTA 1 mM y se adicionaron 500 µl de colorante acetocarmín al 1% por 30 min. Se tomaron las raíces con ayuda de una pinza y se colocaron en un portaobjetos. Se adicionó una gota de ácido acético glacial al 45%, se colocó el cubreobjetos y se realizó presión suave (Squash).
Observación al microscopio
Posterior a la técnica de squash, la muestra se observó bajo el microscopio óptico, con el objetivo de aumento de 10X y 40X. Una vez localizadas las células se ajustó el objetivo 100X, para la observación de los cromosomas.
Duplicación cromosómica
Las plántulas fueron extraídas cuidadosamente del medio de cultivo y se eliminaron los restos del medio de cultivo de las raíces con agua destilada esterilizada. Cada plántula se sumergió hasta la base del tallo en un frasco con tapa que contenía una solución de colchicina (Sigma®) al 0.5%, durante 8 h a 25 °C (Arjunappa et al., 2016). Transcurrido este tiempo las plántulas fueron enjuagadas con agua esterilizada.
Aclimatación de las plántulas de chile
Posterior al tratamiento de colchicina las plántulas fueron transferidas a macetas de 250 ml con sustrato tipo Sunshine( previamente esterilizado. Las plántulas se mantuvieron en la misma cámara a 25 °C con un fotoperiodo de 16 h luz, a una intensidad lumínica de 45-70 μmol m-2 s-1. Se realizó un riego ligero al sustrato cada tercer día con una solución de macroelementos y microelementos (Chambonnet, 1988) al 50%. Una vez que la plántula alcanzó 25 a 30 cm de longitud se tomó nuevamente una muestra radicular para comprobar la duplicación cromosómica.
Diseño experimental y análisis estadístico
El diseño experimental fue un diseño completamente aleatorizado donde se analizaron 27 genotipos experimentales de chile. La unidad experimental fue una caja con 25 anteras por caja Petri y 6 repeticiones por genotipo de chile. El análisis estadístico consistió en un análisis de varianza y comparación de medias a través de la prueba de Tukey (p≤ 0.05), con el paquete estadístico Minitab 17 (Minitab Statistical Software).
Resultados y discusión
Inducción y germinación de embriones haploides
Las anteras mostraron respuesta a la inducción de desarrollo embriogénico entre 4 y 6 semanas posterior a su cultivo. La primera evidencia de inducción de androgénesis fue la aparición en los conductos polínicos de la antera, de un embrión en etapa globular de un tono verde claro a partir de la semana 4 y 5 de cultivo (Figura 1A). Luego el embrión aumentó de tamaño e inició el proceso de diferenciación, la estructura que se define primero son las yemas radiculares, esto inicia con la formación de vellosidades en uno de los extremos del embrión (Figura 1B). Posteriormente, se diferenció la yema apical que conforma los cotiledones, en el lado opuesto en donde se originó la yema radicular (Figura 1C). Tras la separación de la antera (Figura 1D), los embriones germinaron mostrando definición de raíz, hipocotilo y cotiledones (Figura 1E).
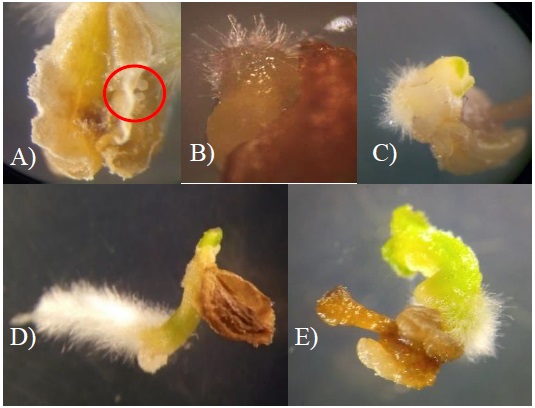
Figura 1 Germinación de embriones y desarrollo de plantas haploides durante el cultivo de anteras del genotipo 3328 de chile ancho. A) pro-embriones en el conducto polínico de la antera; B) formación de yema radicular; C) embrión con inicio de diferenciación de yema apical; D) embrión independizado de la antera; y E) etapas diferenciales de germinación de embrión y elongación de plántula.
Fue posible detectar la pérdida de la capacidad de diferenciación de cotiledones en algunos embriones, es común que los pro-embriones y embriones detengan su desarrollo en alguna parte del proceso, esto se observó en 80% de los pro-embriones y embriones obtenidos con los genotipos de chile probados. Esto se puede atribuir a que las rutas de diferenciación no están genéticamente determinadas al tratarse de tejido haploide y el proceso de inducción (medio de cultivo y shock térmico), no necesariamente culmina en una estructura morfológicamente completa.
Varios reportes coinciden en la importancia de la temperatura sobre los mecanismos celulares (Bajaj 1978; Touraev et al., 1997) así como el efecto en la expresión genética que ocasiona el cambio de microsporas a embriogénesis (Gonzalez-Melendi et al., 1996; Testillano et al., 2000; Barany et al., 2001) desviando de un desarrollo normal a la transición gametofítica a esporofítica. La estimulación por choque térmico ha sido fundamental particularmente en los genotipos con pobre respuesta; sin embargo, aun en los genotipos sin respuesta, será conveniente considerar otros factores como medio de cultivo, reguladores, suplementos, genotipo, condiciones de crecimiento de la planta donante, estadio de desarrollo de las microsporas, entre otros.
Diez de los 27 genotipos de chile empleados no mostraron respuesta a los medios de inducción y germinación. Estos genotipos mostraron anteras no viables en su mayoría y aquellas que iniciaron la germinación en embriones inducidos se detuvieron en algún punto de su diferenciación. Por su parte, los 17 genotipos que mostraron señales de inducción mostraron variabilidad en la eficiencia de respuesta y obtención de plantas haploides de 0.67 a 14.67% con respecto a las anteras viables (Cuadro 1).
Cuadro 1 Eficiencia de regeneración de plantas haploides mediante la inducción de androgénesis en chile.
| Genotipos de chile | Característica | Tipo | Núm. de anteras cultivadas viables+ | Núm. de plantas derivadas de anteras | Eficiencia (%)* |
| 3239 | Tamaño de fruto Susceptible | mulato | 123 | 1 b† | 0.81 |
| 3328 | Mantenedoras de CMS | ancho | 150 | 7 bc | 4.67 |
| 3334 | Mantenedoras de CMS | mulato | 136 | 7 bc | 5.15 |
| 3335 | Mantenedoras de CMS | mulato | 150 | 0 a | 0 |
| 3336 | Mantenedoras de CMS | mulato | 98 | 1 b | 1.02 |
| 3337 | Mantenedoras de CMS | mulato | 132 | 3 b | 2.27 |
| 3338 | Mantenedoras de CMS | mulato | 133 | 7 bc | 5.26 |
| 3339 | Mantenedoras de CMS | mulato | 148 | 0 a | 0 |
| 3340 | Mantenedoras de CMS | mulato | 80 | 5 bc | 6.25 |
| 3341 | Mantenedoras de CMS | mulato | 124 | 3 b | 2.42 |
| 3342 | Mantenedoras de CMS | ancho | 98 | 2 b | 2.04 |
| 3343 | Mantenedoras de CMS | mulato | 107 | 0 a | 0 |
| 3344 | Mantenedoras de CMS | mulato | 126 | 3 b | 2.38 |
| 3345 | Mantenedoras de CMS | mulato | 82 | 0 a | 0 |
| 3346 | Mantenedoras de CMS | mulato | 150 | 1 b | 0.67 |
| 3359 | Mantenedoras de CMS | ancho | 114 | 2 b | 1.75 |
| 3375 | Mantenedoras de CMS | ancho | 115 | 4 b | 3.48 |
| 3378 | Mantenedoras de CMS | ancho | 138 | 0 a | 0 |
| 3381 | PepGMV R, BC3 | ancho | 82 | 0 a | 0 |
| 3382 | PepGMV R, BC3 | ancho | 150 | 0 a | 0 |
| 3383 | PepGMV y P. capsici R, BC3 | ancho | 150 | 22 c | 14.67 |
| 3384 | PepGMV R, BC3 | ancho | 147 | 12 c | 8.16 |
| 3385 | PepGMV R, BC3 | ancho | 140 | 6 bc | 4.29 |
| 3386 | Oidium R, BC3 | ancho | 68 | 0 a | 0 |
| 3387 | PepGMV R, BC3 | ancho | 133 | 0 a | 0 |
| Ancho San Luis | Susceptible | ancho | 133 | 9 c | 6.77 |
| Ancho 3015 | Susceptible | ancho | 142 | 2 b | 1.41 |
Susceptible= genotipos susceptibles a Phytophthora capsici, PepGMV y Cenicilla; R= genotipos segregantes resistentes; BC3= cruza interespecífica ancho 3015 x habanero BG3821 y retrocruza con ancho 3015; += número de anteras viables proviene de la población de 150 anteras divididas en seis repeticiones de 25 anteras cada una; *= número de plantas por cada 100 anteras cultivadas; †= medias con la misma letra en la misma columna son estadísticamente iguales (Tukey, p≤ 0.05).
Lo anterior ha sido reportado de manera uniforme por diversos autores quienes establecen que uno de los principales factores para la inducción de androgénesis con fines de obtención de plantas haploides es el genotipo, encontrando que hay genotipos de chile que no responden a la inducción de androgénesis (Comlekcioglu y Ellialtıoğlu, 2018; Shimira et al., 2019).
Aun cuando en este trabajo no se empleó pretratamiento a bajas temperaturas (4 °C), no hay consenso de que proporcione alguna ventaja relevante; sin embargo, el choque térmico a 35 °C fue mencionado como fundamental para la estimulación de la división de microsporas de chile (Kim et al., 2004; Barany et al., 2005; Vivek et al., 2017; Shimira et al., 2019). De acuerdo con Barany et al. (2005), la morfogénesis de los embriones derivados de microsporas con la misma estimulación de altas temperaturas reveló cambios en la organización celular, la secuencia en el tiempo de los eventos estructurales relacionados con la actividad proliferativa y de diferenciación.
Estos cambios principalmente afectan los plastidios, el compartimento vacuolar, la pared celular y el núcleo. En este trabajo, las anteras se incubaron a 32 °C debido a que 35 °C las anteras se deshidrataban hasta el punto de su inviabilidad. De esta manera, la reducción del tratamiento de inducción a 32 °C permitió mantener la viabilidad de las microsporas, la inducción de androgénesis y posterior germinación de embriones.
Una vez que se observaron estructuras de pro-embriones, la siguiente etapa fue la germinación del embrión a partir de la elongación de la yema radicular a partir de los 28 días en medio de germinación (Figura 2A), así como el engrosamiento y separación de los bordes laterales de la antera aproximadamente a los 40 días de cultivo en medio de germinación (Figura 2B). Posteriormente, a 50-60 días de cultivo se apreció la aparición de primordios foliares (Figura 2C).

Figura 2 Anteras con evidencias de androgénesis y germinación de embrión. A) yema radicular de una antera perteneciente a la línea 3385 a los 28 días en medio de germinación; B) separación de los bordes laterales de la antera de la línea 3384 a los 42 días en medio de germinación; y C) primordios foliares de la línea 3344 a 56 días en medio de germinación.
Se ha reportado otra ruta de diferenciación con la producción de callo derivado del cultivo de anteras en medio Murashige y Skoog suplementado con zeatina 1 mg L-1, 2, 4-D 0.2 mg L-1 y AgNO3 15 mg L-1 con tratamiento a 35 °C por 8 días, de entre 54.02% a 60.92%. Sin embargo, aun cuando los callos manifestaron coloración verde, éstos no mostraron diferenciación de brotes en los híbridos F1 de chile dulce Bharat e Indra, respectivamente (Vivek et al., 2017). De acuerdo con lo obtenido en este trabajo, la formación de callo se observó en baja proporción y no constituyó una limitante para el desarrollo androgénico de las microsporas.
Determinación del nivel de ploidía
La duplicación cromosómica de plantas haploide de chile (n= 12) es necesaria para restablecer el estatus diploide (2n= 24) y con ello la fertilidad. En este trabajo no se observó doble haploides espontáneos por lo que fue necesario establecer la incubación con colchicina para todas las plantas derivadas de androgénesis y embriogénesis (Figura 3A). La concentración de colchicina empleada (0.5%) durante 8 h de exposición (Figura 3B), coincide con la mayoría de los reportes de doble haploides en chile, el método de identificación del contenido cromosómico en estadio de metafase (Figura 3C), fue eficiente y la duplicación cromosómica fue comprobada días después en etapa de aclimatación (Figura 3D). Comlekcioglu (2021) reporta que la inclusión de colchicina (0.3%), desde el medio de inducción, incrementa la tasa de duplicación cromosómica espontánea de 33% a 57.6%, lo cual se contempla dentro de las perspectivas para incorporar en futuros ensayos. Las plantas de chile doble haploides desarrollaron un porte normal en invernadero, recobraron su fertilidad y produjeron frutos (Figura 3E).

Figura 3 Duplicación y método de tinción de cromosomas por el método squash. A) plántula de chile mulato 3342 (n); B) tratamiento con colchicina (0.5 %) por 8 h; C) cromosomas en metafase antes de la duplicación cromosómica; D) cromosomas después de la duplicación cromosómica; y E) planta de chile mulato 3242 doble haploide, en invernadero.
Conclusiones
El protocolo desarrollado en esta investigación para obtención de plantas de chile poblano doble haploides establece tasas diferenciales de inducción de androgénesis y germinación de embriones en las poblaciones empleadas. Los resultados plantean que, si bien los genotipos de chile empleados tienen características genómicas y genéticas diferentes, se establecen las bases generales de inducción de embriones androgénicos, las cuales constituyen perspectivas de mejora.

