











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
La ‘pudrición escamosa’ es una de las enfermedades de mayor importancia en el nopal, se caracteriza por la aparición de escamas grisáceas de forma ondulada o semi ondulada (Souza et al., 2010). Las escamas producidas pueden llegar a cubrir completamente al cladodio, provocando una pérdida de la superficie fotosintética de la planta con pérdidas significativas en la producción (Souza et al., 2010). Los agentes causales de la ‘pudrición escamosa’ son los hongos Neoscytalidium hyalinum y Scytalidium lignicola, que además pueden causar pudrición de raíz en el nopal (Souza et al., 2017; Feijo et al., 2019).
Para controlar a estos y otros hongos fitopatógenos se suele utilizar fungicidas químicos, los cuales son efectivos, pero a menudo tienen efectos negativos en el ambiente y en la salud de los consumidores, además siempre está la posibilidad de la aparición de cepas resistentes (Spadaro y Gullino, 2004; Deising et al., 2008). Los problemas que genera el uso de fungicidas químicos han hecho que se busquen métodos alternativos de control tales como, hongos y bacterias antagonistas de las cuales se puede destacar las bacterias del género Bacillus sp. (Wilson, 1997; Santoyo et al., 2012). Entre las capacidades antagónicas que tiene Bacillus sp., se encuentran la competencia por espacio y nutrientes, la antibiosis y la resistencia sistémica inducida (Layton et al., 2017).
La resistencia sistémica inducida (RSI) es un estado en el cual se mejora la capacidad de defensa de la planta provocado por la interacción de la planta con bacterias promotores de crecimiento (Vallad y Goodman, 2004). Estas bacterias producen liposacáridos, antibióticos y sideróforos que sirven como elicitores de la RSI, a los cuales se les llama patrones moleculares asociados a microorganismos (MAMP) (Wiesel et al., 2014; Canchignia et al., 2015). Una vez la planta detecta los elicitores esta comienza a producir ácido jasmonico (JA) y etileno (ET), estos promueven la síntesis de proteínas relacionadas a la patogenicidad (PR) permitiéndole a la planta reaccionar más eficientemente ante el ataque de algún patógeno (Canchignia et al., 2015).
Se sabe que Bacillus sp., puede inducir resistencia en distintas plantas y ayudar a disminuir los daños causados por hongos como Fusarium sp., Alternaria sp., Colletotrichum sp., Pythium sp. y Phytophthora sp. (Nakkeeran et al., 2007; Chowdappa et al., 2013; Wang et al., 2014; Gond et al., 2015). En la actualidad, se desconoce si Bacillus sp. es capaz de inducir resistencia en el nopal y evitar la enfermedad de la ‘pudrición escamosa’. Por lo que, el objetivo del presente trabajo fue aislar e identificar hongos asociados a la ‘pudrición escamosa’ y evaluar la capacidad de las bacterias del género Bacillus sp., para evitar la enfermedad de la ‘pudrición escamosa’ en el nopal (Opuntia ficus-indica (L) Mill.).
Materiales y métodos
Área de estudio y toma de muestras
Las áreas de estudio fueron una huerta de nopales en el municipio de Gómez Farias, Tamaulipas con coordenadas 23° 09’ 37.64’’ latitud norte. 99° 09’ 12.23’’ longitud oeste y también los módulos hidropónicos del Instituto Tecnológico de Ciudad Victoria con coordenadas 23° 45’ 16.94’’ latitud norte. 99° 10’ 01.76’’ longitud oeste. Los cladodios de nopal pertenecientes a la especie Opuntia ficus-indica con síntomas de la ‘pudrición escamosa’ se consiguieron mediante un muestreo aleatoria en cada uno de los sitios y cada muestra individual se colocó en bolsas de polietileno etiquetada con los datos de recolecta. Por cada uno de los sitios se obtuvieron 20 muestras, las cuales fueron trasportadas al laboratorio de microbiología y biotecnología de la División de Estudios de Posgrados e Investigación del Instituto Tecnológico de Ciudad Victoria para su posterior uso.
Aislamiento de hongos y bacterias
Para el aislamiento de hongos se cortaron fragmentos (1 cm x 1 cm) de los nopales que presentaran los síntomas producidos por la ‘pudrición escamosa’. Estos fragmentos se desinfectaron con alcohol al 70% durante un minuto, después se enjuagaron con agua destilada y dejaron secar sobre papel filtro. Una vez secos, fueron colocados en cajas Petri con medio de cultivo papa dextrosa agar (PDA) y se incubaron durante dos semanas a 30 °C.
Las bacterias fueron aisladas con la técnica de dilución en placa, para esto un gramo de raíces del nopal fueron maceradas en mortero y mezcladas con 9 ml de solución salina. Después, se realizaron diluciones seriadas hasta obtener un factor de dilución de 1 x 106. De esta solución 100 µl se colocaron en cajas Petri con medio cultivo PDA para después esparcirlo con un asa de vidrio. Los hongos se utilizaron para realizar cultivos monospóricos (Valiente y Pavone, 2013) y las bacterias se resembraron a partir de una sola colonia con el asa bacteriológica y resembrándola en una caja Petri con medio cultivo PDA.
Identificación del género de hongos y bacterias
Los hongos se identificaron a genero mediante las claves taxonómicas propuestas por Watanabe (2010); Phillips et al. (2013). Para esto se prepararon laminillas con la técnica de la cinta adhesiva para posteriormente ser observadas en el microscopio óptico, utilizando el objetivo de 40x. Las bacterias se identificaron mediante pruebas bioquímicas y observación en microscopio utilizando el objetivo 100x y aquellas que fueron Gram (+), de forma bacilar y catalasa (+) se identificaron como del género Bacillus sp. (De Vos et al., 2009; Ruiz-Sánchez et al., 2014).
Pruebas de patogenicidad
Para determinar que hongos aislados generan los mismos síntomas observados en campo se realizaron pruebas de patogenicidad y para esto primero se colocaron palillos de dientes estériles sobre colonias de los hongos aislados y se dejaron en incubación a 37 °C hasta que los hongos crecieron sobre ellos y los cubrieron completamente. Posteriormente se cortaron en fragmentos de 1 cm y se usaron para inocular cladodios de nopal previamente desinfectados con alcohol al 70%. Cada cladodio fue inoculado con cinco fragmentos de palillos y se inocularon tres cladodios por género de hongo aislado, después de esto se colocaron en cámaras de humedad durante un mes.
Pasado este periodo de tiempo se observó cuáles de los nopales infectados presentaron los mismos síntomas observados en campo y de estos se re aisló el hongo de las heridas para confirmar que sea el mismo con el que se inoculo (Swart et al., 2003). Una vez identificado cuál de los géneros de los hongos aislados ocasiona la enfermedad se realizaron nuevamente pruebas de patogenicidad con todos los hongos de este género y se calculó el porcentaje de incidencia con el número de heridas causadas por el patógeno.
Pruebas de antagonismo in vitro
Para saber cuáles de las bacterias tienen potencial para controlar a los patógenos se realizaron pruebas in vitro de cultivo dual, colocando en el centro de una caja Petri con medio PDA estéril un fragmento del hongo patógeno obtenido con un sacabocados de un diámetro de 0.5 cm de una colonia del hongo de tres semanas. Para después inocular las bacterias aisladas mediante un asa bacteriológica, estas fueron colocadas en forma circular alrededor del hongo en la parte más externa de la caja, posteriormente se dejaron en incubación a 37 °C durante una semana o hasta que el testigo cubriera completamente la caja, se hicieron 15 repeticiones por cada posible combinación (Mohammadi et al., 2017). Una vez pasado el tiempo de incubación se midieron los radios del patógeno en enfrentamiento y de los testigos y se usaron para calcular el porcentaje de inhibición radial (PICR) (Reyes-Ramírez et al., 2011) mediante la siguiente fórmula:
Donde: RP= radio del patógeno en enfrentamiento; RT= radio del patógeno testigo.
Extracción de ADN
Para las extracciones primero se inocularon suspensiones de esporas de los hongos en matraces de 500 ml con 250 ml de medio de cultivo liquido (Sacarosa 20 g L-1, peptona 5 g L-1, extracto de levadura 5 g L-1 y agua destilada estéril 1 L) y se dejaron en incubación durante una semana a 32 °C. Después del tiempo de incubación se filtraron los medios de cultivo, luego el micelio recuperado se colocó en microtubos de 1.5 ml. Se empleó el método de extracción con DNAzol® para purificar el ADN, siguiendo el protocolo propuesto por Guo et al. (2005) con algunas modificaciones, que constaron en macerar la muestra con un pistilo de plástico después de agregar el DNAzol® y aumentar el tiempo de incubación a 45 min. Para las bacterias primero se inocularon tubos de ensayo con medio de cultivo LB y se incubaron a 32 °C durante dos días. Las extracciones de ADN se realizaron mediante el kit DNAzol® y se siguieron las instrucciones del fabricante.
Una vez que se realizaron las extracciones tanto de hongos como bacterias se procedió a su visualización. Esto se realizó al mezclar 2 µl de muestra y 2 µl de Buffer de corrida azul conteniendo el colorante para ADN SYBRGreen®. Luego las muestras se colocaron en un gel de agarosa al 1% en una cámara de electroforesis 14 OWL® (8 cm x 15 cm) con buffer SB x1 (ácido bórico-NaOH) a un voltaje de 100 durante 15 min, después de esto el gel se colocó en un transiluminador de luz UV y se foto documento mediante el programa Logitech webcam.
PCR-secuenciación
Se realizó la amplificación de las regiones ITS1-5.8s-ITS2 del ADN nuclear de los hongos y el 16S RNAr de las bacterias mediante PCR con los iniciadores ITS1 (5’- TCCGTAGGTGAACCTGCG G3’) e ITS4 (5’-TCCTCCGCTTATTGATATGC-3’) (White et al., 1990) para los hongos y los iniciadores Y1 (5’-CCAGCAGCCGCGGTAATACG-3’) y Y2 (5’- ATCGG(C/T) TACCTTGTT ACGACTTC-3’) para bacterias (Lu et al., 2000). Cada reacción de PCR contiene 21 µl de mix (agua 14.7, buffer 10x 2.5, DNTPs 25mM, 0.25 µl, BSA 25x 1 µl), 2 µl de ADN, 2 µl de primers ITS1-4 para los hongos y Y1-Y2 para las bacterias y 0.3 µl de TaqPol para un volumen de 25.3 µl.
La reacción de PCR se llevó a cabo en un termociclador BIO-RAD T-100TM para esto se utilizaron los siguientes ciclos de temperatura para los hongos: pre desnaturalización de 5 min a 94 °C, seguido de 30 ciclos de desnaturalización a 95 °C por 1 min, alineación a 54 °C por 30 s y extensión de 1 min a 72 °C y extensión final de 5 min a 72 °C (Moo-Koh et al., 2017), para las bacterias los ciclos usados fueron los siguientes: pre desnaturalización de 10 min a 95 °C, seguido de 30 ciclos de desnaturalización a 95 °C por 1 min, alineación a 59.9 °C por 1 m y extensión de 2 min a 72 °C y extensión final de 1 min a 72 °C (Lu et al., 2000).
Después de obtener los productos amplificados tanto de la región ITS 1-2 y del 16 S RNAr mediante PCR estos fueron enviados a la facultad de ciencias de la Universidad Nacional Autónoma de México (UNAM) para su secuenciación. Las secuencias obtenidas fueron editadas con el programa BIOEDIT y comparadas con secuencias de la base de datos del Banco de Genes del National Center for Biotechnology Information mediante Basic Local Alignment Search Tool (BLAST). Además, el programa CLC Sequence Viwer 8 fue utilizado para realizar un árbol filogenético con el método de UPGMA con el cual se compararon las secuencias obtenidas con secuencias de Banco de Genes.
Pruebas de inducción de resistencia en nopal
Una vez que se determinó cuáles son los aislamientos bacterianos más efectivos para controlar al patógeno in vitro y cuáles son los hongos aislados con mayor porcentaje de patogenicidad se realizaron las pruebas de inducción de resistencia. Para esto se colocaron tres cladodios de nopal en bolsas negras para plantas con peat moss como sustrato hasta que enraizaron, se prepararon un total de ocho bolsas. Posteriormente se inocularon con las bacterias elegidas que previamente se hicieron crecer en matraces Erlenmeyer de 500 ml con 250 ml de medio de cultivo líquido que contenía sacarosa (20 g L-1), peptona (30 g L-1), extracto de levadura (7 g L-1), KH2PO4 (1.9 g L-1) y MgSO4 (0.45 g L-1). Las raíces fueron inoculadas con 10 ml de cultivo y 72 h después se inocularon los cladodios con los hongos seleccionados (Swart et al., 2003).
Diseño experimental
Los experimentos de antagonismo in vitro e in vivo constaron de 15 repeticiones por tratamiento y fueron realizados mediante un diseño experimental completamente al azar. Todos los datos obtenidos pasaron las pruebas de normalidad y de homogeneidad de varianza y fueron comparados con un análisis de varianza de una sola vía (Anova) y una comparación de medias de Tukey, ambos con un valor de significancia de 0.05. Todos los análisis estadísticos se realizaron con Excel 2016.
Resultados y discusión
Hongos y bacterias aisladas
Se lograron aislar e identificar un total de 33 hongos para ambos sitios, 20 en el módulo hidropónico del ITCV y 14 en la huerta de Gómez Farías. Neoscytalidium sp. fue el género más común en ambos sitios pues es un hongo abundante como patógeno en lugares con climas tropicales, tal como mencionan Souza et al. (2010); Souza et al. (2017). El género Neoscytalidium sp., presentó colonias de color verde obscuro y formaron esporas en cadena, cilíndrica con un septo característico del género (Phillips et al., 2013). Otros hongos aislados de los cladodios de nopal fueron Fusarium sp. al que se le considera agente causal de la ‘pudrición de la punta’ y por causar lesiones necróticas en el nopal (Swart y Kriel, 2002; Ammar et al., 2004; Souza et al., 2010).
El género Alternaria sp., también fue aislado, causa la enfermedad conocida como ‘mal de oro’ y la ‘pudrición seca’ además de que puede provocar lesiones necróticas en la tuna (Swart y Kriel, 2002; Swart et al., 2003; Méndez et al., 2007; Faedda et al., 2016). Además, se aislaron hongos del género Curvularia sp. el cual se le asocia con lesiones circulares de color café a obscuro y de aspecto aterciopelado (Flores et al., 2013). Por último, se consiguieron hongos del género Schizophyllum sp., pero este género no está asociado con ninguna enfermedad.
Por otra parte, se aislaron 10 bacterias en ambos sitios de muestreo, pertenecientes al género Bacillus sp., ya que todas presentaron las características típicas del género como ser catalasa positiva, y Gram positivo, las colonias tuvieron bordes irregulares y al observarse al microscopio estas tuvieron forma bacilar (De Vos et al., 2009; Ruiz-Sánchez et al., 2014). Las bacterias pertenecientes al género Bacillus sp. fueron usadas en las pruebas de antagonismo in vitro.
Identificación molecular de los hongos y bacterias
Las secuencias obtenidas de los aislamientos de Neoscytalidium sp. fueron comparadas en BLAST y los aislamientos M7, M9, G81 y G112 tuvieron una similitud de 100% con la especie Neoscytalidium dimidiatum, por otra parte, las secuencias de M2, M3, M4, M5, M6 M8, M10, G42, G43, G11, G311 tienen un 100% de similitud con secuencias de la especie Neoscytalidium hyalinum. Al realizar el análisis filogenético de las secuencias y compararlas con secuencias obtenidas de GenBank de las especies N. hyalinum (MK387852.1 y MK387853.1), N. dimidiatum (KP132488.1 y KP132489.1), N. novaehollandiae (NR-111260.1 y MH863173.1) y N. orchidacearum (KY933091.1) se observó que las secuencias estaban mayormente relacionadas con secuencias de los hongos N. hyalinum y N. dimidiatum, así que se consideró que los hongos aislados pertenecen a estas especies (Figura 1).

Figura 1 Árbol filogenético de Neoscytalidium sp., basado en las secuencias de la región ITS 1-2, donde se usó a Amanita muscaria (AB015700.1) como grupo externo (AB015700.1).
Originalmente tanto N. dimidiatum y N. hyalinum formaban parte del género Scytalidium sp., pero actualmente estas y otras especies se clasifican en el género Neoscytalidium sp. que es una mezcla entre Scytalidium sp. y Fusicoccum sp., además tanto N. hyalinum y N. dimidiatum son considerados actualmente la misma especie (Phillips et al., 2013; Machado et al., 2014).
Se realizó el análisis filogenético con las secuencias del 16s RNAr de las bacterias con mayores porcentajes de inhibición de las pruebas in vitro, las secuencias de G11, G21 y G31 se encontraron altamente relacionadas con secuencias de B. amyloliquefaciens (NR-041455.1 y NR-116022.1) considerándose que las bacterias aisladas pertenecen a la especie B. amyloliquefaciens (Figura 2).

Pruebas de patogenicidad y porcentajes de incidencia
En las pruebas de patogenicidad, los hongos del género Neoscytalidium sp., ocasionaron la aparición de manchas de color marrón que se tornaron a un color más obscuro, los cuales son los síntomas iniciales de la ‘pudrición escamosa’ (Souza et al., 2010; Feijo et al., 2019). De los distintos aislamientos de Neoscytalidium sp, el aislamiento G111 presento una mayor incidencia con 93.33% y los aislamientos M10 y G93 tuvieron los menores porcentajes de incidencia con un 33.33%.
Estas diferencias están dadas por la capacidad de la planta para detectar a los patógenos mediante proteínas en las membranas de sus células llamadas proteínas R que sirven para detectar unas proteínas producidas por el patógeno conocidas como proteínas Avr (Ordeñana, 1998; Wiesel et al., 2014). En caso de que exista una compatibilidad entre las proteínas R de la planta y las proteínas Avr del patógeno, se desencadena una reacción de hipersensibilidad que ocasiona la muerte celular programada y la resistencia sistémica adquirida como defensa en toda la planta, ambos mecanismos evitan o reducen el daño causado por el patógeno (Ordeñana, 1998; Durrant y Dong, 2004; Forouhar et al., 2005; Días, 2012).
Antagonismo in vitro
Los radios obtenidos de las pruebas de cultivo dual (Figura 3 y 4) se compararon con una prueba de Anova (0.05), mostrando diferencias significativas entre los tratamientos (p≤ 0.05). Al comparar los tratamientos mediante la prueba de Tukey (0.05) (Cuadro 1) se observó que todas las bacterias reducen significativamente el radio del patógeno al comprar con el testigo y entre los tratamientos la bacteria G11 obtuvo diferencias significativas con respecto al resto de los tratamientos, pues esta consiguió los mayores porcentajes de inhibición del patógeno con una media de 50.09%. Asimismo, los aislamientos G21 y G31 mostraron diferencias significativas con la mayoría de los tratamientos excepto entre sí, ambas tienen una media de porcentaje de inhibición de 41.84% y de 41.73%, respectivamente. Todos los aislamientos bacterianos ocasionaron halos de inhibición debido a que la bacteria libera antibióticos en el medio, siendo los más comunes las surfactinas, iturinas y fengicinas (Villarreal et al., 2018).
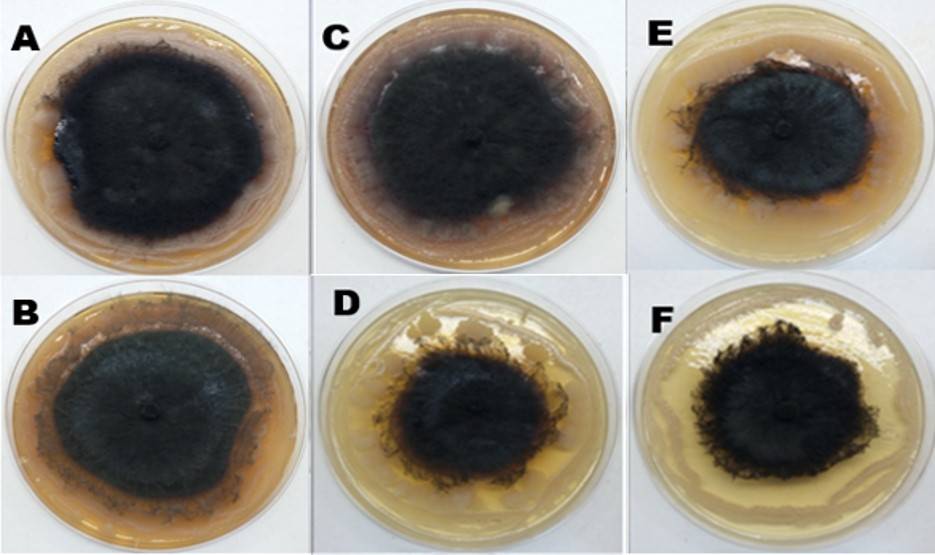
Figura 3 Pruebas de antagonismo in vitro mediante cultivo dual. A) G111vBMN3; B) G111vBMN21; C) G111vBMN22; D) G111vG11; E) G111vG21; y F) G111vG31.

Figura 4 Pruebas de antagonismo in vitro mediante cultivo dual. G) G111vG51; H) G111vG52; I) G111vG53; J) G111vG54; y K) G111 testigo.
Cuadro 1 Resultados de la prueba de Tukey (0.05) de las pruebas in vitro, tratamientos con la misma letra no tienen diferencias significativas.
| Tratamientos | Promedios |
| Testigo (G111) | 4.24 ±0.053 a |
| G51 | 2.83 ±0.053 b |
| NM3 | 2.95 ±0.074 c |
| MN22 | 2.84 ±0.059 b |
| G52 | 2.93 ±0.074 c |
| G21 | 2.46 ±0.069 d |
| G54 | 2.7 ±0.056 e |
| G53 | 2.79 ±0.054 b |
| G11 | 2.11 ±0.065 f |
| MN21 | 2.83 ±0.057 b |
| G31 | 2.47 ± 0.06 d |
Las diferencias en los porcentajes de inhibición observadas en las pruebas in vitro pueden deberse a la cantidad de antibióticos que producen las bacterias por ejemplo Bacillus subtilis dedica un 5% de su genoma a producir antibióticos mientras que Bacillus amyloliquefaciens dedica 10% de su genoma a la producción de estos (Chowdhury et al., 2015; Gond et al., 2015).
Pruebas in vivo de inducción de resistencia
Los diámetros obtenidos en los experimentos fueron usados para realizar una prueba de Anova de una vía (0.05), en donde se encontró que estos presentan diferencias significativas (p≤ 0.05). Al comparar los tratamientos con la prueba de Tukey (0.05) (Cuadro 2) las bacterias mostraron reducir el daño causado por el patógeno de forma significativa. Durante la resistencia inducida se disminuye el tamaño de las heridas causadas por el patógeno debido a la acumulación de proteínas PR como glucanasas y quitinasas (Wiesel et al., 2014; Canchignia et al., 2015). Al comparar entre tratamientos las bacterias G11, G21 y G31 no mostraron diferencias significativas para reducir el daño que causa el hongo N. hyalinum.
Cuadro 2 Resultados de la prueba de Tukey (0.05) de la prueba de inducción de resistencia, tratamientos con la misma letra no tienen diferencias significativas.
| Tratamiento | Promedio |
| Testigo(G111) | 4.27 ±0.069 a |
| G11 | 0.39 ±0.075 b |
| G21 | 0.47 ±0.072 c |
| G31 | 0.45 ±0.07 bc |
La bacteria G11 obtuvo en promedio un valor de 0.39 cm, en el tratamiento con la bacteria G21 se consiguió un promedio de 0.47 cm, en el tratamiento con la bacteria G31 se alcanzó un promedio de 0.45 cm y por último el promedio de los diámetros de las heridas del tratamiento testigo fueron de 4.27 cm. Se desconoce los genes que se activan en el nopal al producirse la inducción de resistencia (Figura 5). En plantas de maíz inoculadas con B. subtilis se activan los genes PR-1 y PR-4, mientras que en plantas de lechuga al ser inoculadas con B. amyloliquefaciens se activan los genes PR-1 y PDF1.2 (Chowdhury et al., 2015; Gond et al., 2015). Esta activación ocasiona un incremento en la producción de enzimas relacionadas a la defensa como la peroxidasa, polifenol oxidasa y el superóxido dismutasa las cuales disminuyen o evitan el daño causado por los patógenos en las plantas (Ongena et al., 2005; Chowdappa et al., 2013).

Conclusiones
Las bacterias aisladas pertenecientes al género Bacillus sp. lograron disminuir el diámetro del patógeno N. hyalinum (G111) en las pruebas in vitro mediante antibiosis destacándose los aislamientos G11, G21 y G31 identificadas como B. amyloliquefaciens. Los resultados obtenidos en las pruebas in vivo muestran que los aislamientos son capaces de reducir el daño causado por N. hyalinum (G111). Pero, aunque los aislamientos de B. amyloliquefaciens logran inducir resistencia en el nopal se desconoce cuáles son los antibióticos que los aislamientos G11, G21 y G31 producen y cuales genes PR logran activar en el nopal, por lo que futuras investigaciones deberían estar enfocadas en cuales son los antibióticos que producen los aislamientos G11, G21 y G31 y los genes PR que inducen en el nopal.

