











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
La calidad de la luz, el flujo de fotones y el fotoperíodo son aspectos de la luz que afectan el desarrollo de las plantas, por los efectos específicos que éstos tienen sobre diferentes tipos de respuestas de las plantas, como la fotosíntesis, fotomorfogénesis y otros procesos fisiológicos y bioquímicos (Hogewoning et al., 2010; Nelson y Bugbee, 2015; Snowden et al., 2016; Yan et al., 2019). La lámpara fluorescente compacta representa una forma simple y económica de reemplazar a la lámpara incandescente utilizada en cámaras de crecimiento (Runkle et al., 2012).
El espectro de luz emitido por una lámpara fluorescente compacta es diferente al de una lámpara incandescente. Una lámpara incandescente emite más luz roja lejana (RL= 700-800 nm) que roja (R= 600-700 nm) y tiene baja proporción R:RL, de 0.7, aproximadamente (Runkle et al., 2012; Gupta y Jatothu, 2013), que provoca alargamiento de tallos, expansión foliar y otras respuestas inducidas por fitocromos (Casal, 2013; Demotes-Mainard et al., 2016). En contraste, la lámpara fluorescente compacta emite más luz azul (A= 400-500 nm) y roja, y poca de rojo lejano, por lo que la proporción R:RL es entre 3 y 8, dependiendo de las bandas de onda utilizadas y el modelo de lámpara, las cuales inducen crecimiento de plantas más compactas (Runkle et al., 2012; Gupta y Jatothu, 2013).
Los diodos emisores de luz (leds) emergieron como una fuente de luz novedosa y eficiente para promover el crecimiento de las plantas en cámaras de investigación con espacio limitado (Cope y Bugbee, 2013). Con leds se puede proporcionar luz roja o azul monocromática, pero ninguna de éstas logra satisfacer el requisito del crecimiento normal de la planta (Wang et al., 2016; Yang et al., 2017). Sin embargo, cuando se suministran adecuadas proporciones R:A, suplementadas o no con otras longitudes de onda: UV-A (350-400 nm), verde (500-600 nm) o rojo lejano, se obtiene mayor compacidad en plántula de tomate (Javanmardi y Emami, 2013; Hernández et al., 2016), mayor biomasa, clorofila y capacidad fotosintética de plántulas de pepino (Hogewoning et al., 2010; Song et al., 2017) y tomate (Xiaoying et al., 2012), óptima producción de plántulas de lechuga (Yan et al., 2019) y papa (Chen et al., 2020).
No obstante, varios estudios han informado mayor crecimiento bajo lámparas fluorescentes en comparación con leds con el mismo flujo fotónico (Lin et al., 2013; Chen et al., 2014; Wang et al., 2014). Las lámparas fluorescentes tienen más luz difusa en comparación con la luz directa de los leds. La luz difusa penetra mejor en el dosel vegetal que la luz directa e incrementa la fotosíntesis y la biomasa seca (Li y Yang, 2015). Las lámparas fluorescentes también aumentan la radiación infrarroja (Nelson y Bugbee, 2015) y tienen algo de radiación RL (Snowden et al., 2016), lo cual aumenta la expansión de hojas y pecíolos y por lo tanto, la captura de radiación.
Está disponible una diversidad de lámparas fluorescentes compactas que difieren en flujo luminoso (lm); eficacia luminosa (lm W-1). vida útil (h), índice de rendimiento de color (IRC) y apariencia de color, más comúnmente descrita como temperatura de color y expresada en unidades Kelvin (K). De la cual hay mayor disponibilidad en tres grupos principales: 2 700 a 3 000 K, que produce luz blanca cálida similar a la de las lámparas incandescentes, 3 500 a 4 100 K de luz blanca neutra o natural y 5 000 a 6 500 K de luz blanca fría, que proporciona luz con tonalidad azul (Saavedra et al., 2016).
La luz es un factor ambiental importante también para los hongos, que tiene un papel importante en las interacciones planta-patógeno y controla varias actividades metabólicas de hongos patógenos (Rahman et al., 2003; Wang et al., 2010). Se ha descubierto que más de 100 especies de hongos, que representan todos los filos, responden al efecto de la luz (Tisch y Schmoll, 2010). Entre las actividades metabólicas que regula están incluidos los ritmos circadianos, la conidiación asexual, la pigmentación, el metabolismo secundario y el desarrollo sexual (Purschwitz et al., 2006; Suzuki et al., 2018).
Aunque la interacción entre la luz y los hongos ha sido estudiada y revisada por varios investigadores (Idnurm y Heitman, 2005; Purschwitz et al. 2006; Chen et al., 2009), los informes sobre el efecto de la calidad de la luz en la patogenicidad o la virulencia de los hongos son limitados. El objetivo de la investigación fue determinar el efecto del espectro luminoso emitido por lámparas fluorescentes compactas de luz blanca fría, neutra y cálida, sobre el crecimiento de plantas de pepino (Cucumis sativus L.) y la severidad de cenicilla de las cucurbitáceas (Oidium sp.).
Materiales y métodos
La investigación se realizó dentro de cámaras de crecimiento de 44 x 70 x 80 cm (246 400 cm3) con malla tejida de 16 x 16 monofilamentos cristalinos de polietileno de alta densidad por cm2, en todos sus lados y los costados revestidos con papel Mylar de alta reflectancia (Figura 1) en la Facultad de Agronomía de la Universidad Autónoma de Sinaloa. Se sembraron semillas de pepino ‘Poinsett 76’ en charolas de poliestireno de 128 cavidades y cuando las plántulas contaron con dos hojas verdaderas, fueron individualmente trasplantadas en vasos de poliestireno de 0.5 L. En ambos tipos de contenedores se utilizó turba (Pro-Mix® FLX, Premier Horticulture, EE UU) como sustrato y fueron regados hasta saturación con una solución fertilizante (pH= 6.5 a 7 y CE= 1.12 a 1.35 dS m-1) compuesta por 0.5 a 1.2 g L-1 de fosfato monopotásico y 0.3 a 0.7 g L-1 de nitrato de potasio.

Figura 1 Plantas de pepino dentro de cámara de crecimiento con iluminación de lámparas fluorescentes de luz blanca fría (LBF, izquierda), neutra (LBN, centro) y cálida (LBC, derecha).
Se utilizaron lámparas fluorescentes (FLE23HLX, GE, EE. UU) de luz blanca fría (LBF= 6 500 K), neutra (LBN= 4 000 K) y cálida (LBC= 2700 K). La medición del flujo de fotones, en el rango de 350 a 1 050 nm, fue realizada con espectrorradiómetro (Field SpecPro®Vnir, ASD, EE. UU), obteniéndose: a) cantidad absoluta de la densidad de flujo de fotones fotosintéticos (DFFF= 400 a 700 nm), luz azul (A= 400 a 500 nm), luz roja (R= 600 a 700 nm) y luz roja lejana (RL= 700 a 800 nm); b) cantidad relativa (% del total de DFFF) de luz azul (A:DFFF) y roja (R:DFFF); y c) cantidad proporcional de luz azul a roja (A:R), azul a roja lejana (A:RL), roja a azul (R:A) y roja a roja lejana (R:RL) (Cuadro 1). El espectro de luz de las tres lámparas fluorescentes se muestra en la Figura 2.
Cuadro 1 Parámetros de calidad de la luz emitida por lámparas fluorescentes de luz blanca fría (LBF), neutra (LBN) y cálida (LBC).
| Parámetro/tipo de lámpara | LBF | LBN | LBC |
| DFFF (400-700 nm)x | 306.4 | 305.25 | 304.37 |
| A (400-500 nm)x | 84.19 | 64.87 | 45.74 |
| R (600-700 nm)x | 81.34 | 100.44 | 122.04 |
| RL (700-800 nm)x | 31.79 | 33.93 | 37.51 |
| A:DFFF [(400-500/400-700 nm)*100]y | 27.48 | 21.25 | 15.03 |
| R:DFFF [(600-700/400-700 nm)*100]y | 26.55 | 32.9 | 40.1 |
| A:R (400-500/600-700 nm)z | 1.04 | 0.65 | 0.37 |
| A:RL (400-500/700-800 nm)z | 2.65 | 1.91 | 1.22 |
| R:A (600-700/400-500 nm)z | 0.97 | 1.55 | 2.67 |
| R:RL (600-700/700-800 nm)z | 2.56 | 2.96 | 3.25 |
DFFF= densidad de flujo de fotones fotosintéticos. A= luz azul; R= luz roja; RL= luz roja lejana. Cantidades absolutasx (µmol m-2 s-1), relativasy (%) y proporcionalesz (adimensional).
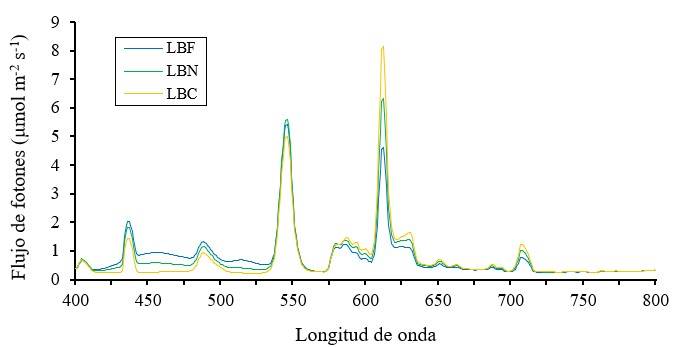
Figura 2 Distribución espectral de la luz emitida por lámparas fluorescentes de luz blanca fría (LBF), neutra (LBN) y cálida (LBC).
El ambiente luminoso presentó medias de 305.3 μmol m-2 s-1 de DFFF y 13.2 mol m-2 d-1 de luz diaria integrada, adecuado para la producción de plántula de hortalizas (Fan et al., 2013). El fotoperiodo fue de 12/12 h de luz/obscuridad. La temperatura y humedad relativa, registradas con termohigrómetros (CM-DT171, Twilight, México), presentaron medias ± error estándar de 24.5 ±0.15 °C y 62.5 ±0.6%, respectivamente. El nivel de CO2 fue de 420 ±20 µmol mol-1, obtenida con medidor de CO2 (CO2-100, Amprobe, Alemania).
Para el estudio del crecimiento de las plantas se utilizó un diseño experimental completamente al azar con tres tratamientos: lámparas fluorescentes compactas de luz blanca fría (LBF), neutra (LBN) y cálida (LBC) y cuatro repeticiones (cuatro plantas por repetición). Las plantas de pepino fueron evaluadas durante 35 días después de la emergencia (dde), período durante el cual fueron expuestas a la luz de las lámparas. El experimento se repitió tres veces, en cada uno de los cuales fueron evaluadas las siguientes variables de respuesta: altura de planta, medida con cinta métrica, diámetro de tallo, obtenido con calibrador digital (6MP, Truper, México), verdor foliar, por medio de un medidor de clorofila (SPAD 502 Plus, Minolta, Japón), área foliar, obtenida con un método no destructivo propuesto por Blanco y Follegati (2003), biomasa fresca y seca de hojas, tallo y raíz por planta, mediante balanza analítica (SA120, Scientech, EE. UU), previo secado en horno (FE293AD, Felisa, México) a 70 °C, hasta peso seco constante.
Para el estudio de severidad de la cenicilla se utilizó un diseño experimental completamente al azar con tres tratamientos: lámparas fluorescentes compactas de luz blanca fría (LBF), neutra (LBN) y cálida (LBC) y 12 repeticiones (una planta por repetición). El inóculo primario de Oidium sp. se obtuvo de plantas de Cucurbita pepo L. infectadas naturalmente.
La suspensión de conidios se preparó cepillando las hojas fuente con agua destilada y se mantuvo con una concentración aproximada de 5.1 x 104 conidios ml-1 mediante hemocitómetro (79003, Cole-Parmer, EE. UU). La inoculación se realizó por medio de la aspersión foliar de 30 ml de suspensión conidial por cada charola de 128 plántulas, el día que desplegaron la segunda hoja verdadera. El experimento se repitió tres veces y cada uno abarcó un período de 35 días después de la inoculación (ddi), durante el cual se determinó el porcentaje de área foliar con síntomas de la enfermedad en cada hoja de las plantas, así como el promedio de la planta completa.
Los datos obtenidos de los tres experimentos fueron promediados y sometidos al análisis de varianza y comparación de medias con la prueba de Tukey (p≤ 0.05), mediante el paquete estadístico Statistica versión 7.0 (StatSoft, 2004).
Resultados y discusión
La calidad de luz emitida por las lámparas fluorescentes (Cuadro 1) ocasionó diferencias significativas (p≤ 0.05) sobre la altura, diámetro de tallo, área foliar y verdor de las plantas de pepino (Cuadro 2). El ambiente luminoso creado por lámparas LBC presentó los valores más altos de cantidad absoluta de luz roja (122.04 µmol m-2 s-1), relación R:DFFF (40.1%) y proporción R:A (2.67), que ocasionó aumentos en altura de planta de 3.3 a 30.2% y 15.4 a 82.3%, en comparación con el efecto causado por lámparas LBN y LBF, las cuales emitieron 17.5 y 45.1% menos luz roja que LBC, respectivamente. El resultado de incremento de altura de planta a causa de la luz roja concuerda con el descrito en Paeonia suffruticosa (Ding et al., 2010), Solanum lycopersicum (Xiaoying et al., 2012), Morus alba (Hu et al., 2016), Camptotheca acuminata (Yu et al., 2017) y Solanum tuberosum (Chen et al., 2020).
Los fitocromos, receptores de luz roja y roja lejana, regulan la elongación del tallo tanto por división como por extensión celular (Neff et al., 2000). La actividad de los fitocromos durante el alargamiento celular se controla mediante la biosíntesis de giberelinas AG1 y AG4, principalmente y auxina AIA (Damayanthi-Ranwala y Decoteau, 1998; Kurepin et al., 2007; Fukuda et al., 2016; Li et al., 2017).
Las lámparas LBF emitieron la mayor cantidad absoluta de luz azul (84.19 µmol m-2 s-1), 29.8 y 90.3% mayor comparada con las respectivas lámparas LBN y LBC, lo cual contuvo el alargamiento del tallo, pues se correlacionó negativamente con altura de planta. Estas lámparas produjeron también la más alta relación A:DFFF (27.48%) y la mayor proporción A:R (1.04), además de la más baja relación R:DFFF (26.55%) y la menor proporción R:A (0.97). Este resultado se relaciona con la capacidad de la luz azul para inhibir el alargamiento del tallo (Ding et al., 2010; Xiaoying et al., 2012; Hu et al., 2016; Yu et al., 2017; Chen et al., 2020), como respuesta a interacciones sinérgicas entre fitocromos y criptocromos, receptores de luz roja y azul, respectivamente, en la promoción o inhibición de la elongación del tallo (Heo et al., 2002).
Las características espectrales de la luz emitida por las lámparas LBF, antes referidas, también incrementaron el diámetro de tallo de 0.1 a 9.1% y 5.1 a 13.2% en comparación con el de las plantas que crecieron con luz de lámparas LBN y LBC, respectivamente, las cuales emitieron luz con relación A:DFFF (15.03 y 21.25%) y proporción A:R (0.37 y 0.65), menores que las de LBF. Li et al. (2017); Chen et al. (2020) señalan que la luz azul estimula una mayor expresión de proteínas asociadas a microtúbulos y tubulina vegetales, las cuales promueven formación de pared celular secundaria y como consecuencia, engrosamiento del tallo.
La calidad de luz emitida por las lámparas LBC, con relevancia en la luz roja, promovió aumentos de 2.2 a 14.2% y 18.6 a 25.5% en el área foliar por plántula, en comparación con el efecto ocasionado por las lámparas LBN y LBF, respectivamente.
Resultado congruente con los obtenidos por Cope y Bugbee (2013) en Raphanus sativus y Glycine max y por Hernández y Kubota (2016) en Cucumis sativus, donde el área foliar creció a medida que la luz azul disminuyó y la luz roja aumentó. Ambas, luz azul y roja, estimulan el flujo de protones en las células, la acidificación apoplástica, elasticidad de la pared celular y la acumulación de solutos para el mantenimiento de la turgencia de las hojas en crecimiento, mediante mecanismos separados. La luz azul induce una interacción directa entre la bomba de protones y un fotorreceptor de luz azul, mientras que la luz roja influye indirectamente en la bomba de protones modulando los canales de calcio y potasio (Staal et al., 1994; Van Volkenburgh, 1999).
Mientras que, los parámetros espectrales en la luz emitida por LBF (Cuadro 2) con relevancia en la luz azul, aumentaron el verdor foliar de las plantas de 2.1 a 8% y 10.1 a 18% en comparación con el efecto ocasionado por las lámparas LBN y LBC, respectivamente. Lo anterior concuerda con investigaciones realizadas en plántulas de Solanum lycopersicum (Hernández y Kubota, 2016) y Cucumis sativius (Hogewoning et al., 2010; Hernández et al., 2016), las cuales manifestaron mayor concentración de clorofila foliar conforme aumentó el flujo de fotones azules, debido al efecto aditivo de criptocromos y fitocromos, comparado con menor biosíntesis de clorofila en plantas cultivadas bajo luz azul o roja monocromática.
Cuadro 2 Altura de planta (AP), diámetro de tallo (DT), área foliar por planta (AFP) y verdor foliar (VF) de plantas de pepino cv Poinsett 76, cultivadas en cámara de crecimiento con lámparas de luz blanca fría (LBF), neutra (LBN) y cálida (LBC).
| Parámetro | Tratamiento | Días después de la emergencia | |||||||||
| 7 | 14 | 21 | 28 | 35 | |||||||
| AP (cm) | LBF | 3.98 | c | 4.5 | c | 11.7 | b | 17.8 | c | 43.3 | b |
| LBN | 5.4 | b | 5.59 | b | 12.5 | b | 26.2 | b | 48.4 | ab | |
| LBC | 6.92 | a | 7.28 | a | 14.7 | a | 32.4 | a | 50 | a | |
| DMSH | 0.97 | 0.96 | 1.5 | 5.1 | 5.48 | ||||||
| DT (mm) | LBF | 2.3 | a | 3.3 | a | 4.4 | a | 5.83 | a | 5.98 | a |
| LBN | 2.1 | b | 3.2 | ab | 4.4 | a | 5.34 | b | 5.51 | b | |
| LBC | 2.1 | b | 2.9 | b | 4.2 | b | 5.21 | b | 5.42 | b | |
| DMSH | 0.13 | 0.32 | 0.16 | 0.2 | 0.31 | ||||||
| AFP (cm2) | LBF | 30 | b | 163.6 | b | 529.5 | b | 936.1 | b | 1319.3 | b |
| LBN | 48.4 | a | 200.9 | a | 579.6 | b | 1065.3 | a | 1525.1 | a | |
| LBC | 37 | b | 205.3 | a | 661.7 | a | 1114.8 | a | 1564.4 | a | |
| DMSH | 8.52 | 27.4 | 59.28 | 103.6 | 119.4 | ||||||
| VF (u SPAD) | LBF | 30.1 | a | 41.6 | a | 43.5 | a | 41.5 | a | 36.4 | a |
| LBN | 27.9 | ab | 39.7 | b | 42 | a | 38.4 | b | 35.7 | a | |
| LBC | 25.5 | b | 37.8 | c | 37.8 | b | 36.1 | c | 33 | b | |
| DMSH | 2.94 | 1.84 | 1.63 | 1.99 | 1.82 | ||||||
Medias con la misma letra son estadísticamente iguales (Tukey, p≤ 0.05). DMSH= diferencia mínima significativa honesta.
La producción de biomasa de hojas, tallo y raíces por las plantas de pepino también presentaron diferencias significativas (p≤ 0.05) debidas a la calidad de luz emitida por las lámparas fluorescentes (Cuadro 3).
Cuadro 3 Peso fresco (PF) y seco (PS) de hojas, tallo y raíz de plantas de pepino cv Poinsett 76, cultivadas en cámara de crecimiento con lámparas de luz blanca fría (LBF), neutra (LBN) y cálida (LBC).
| Parámetro | Tratamiento | Días después de la emergencia | ||||||||||
| 7 | 14 | 21 | 28 | 35 | ||||||||
| PF/hojas (g) | LBF | 0.17 | b | 3.93 | a | 11.52 | b | 16.21 | b | 20.18 | b | |
| LBN | 0.2 | ab | 5.22 | a | 12.24 | ab | 16.25 | b | 20.97 | b | ||
| LBC | 0.23 | a | 5.43 | a | 13.29 | a | 17.99 | a | 22.69 | a | ||
| DMSH | 0.03 | 1.57 | 1.42 | 1.02 | 1.4 | |||||||
| PS/hojas (g) | LBF | 0.02 | b | 0.37 | b | 1.06 | a | 1.58 | b | 2.09 | b | |
| LBN | 0.02 | b | 0.47 | a | 1.11 | a | 1.68 | ab | 2.21 | ab | ||
| LBC | 0.03 | a | 0.47 | a | 1.14 | a | 1.8 | a | 2.49 | a | ||
| DMSH | 0.006 | 0.096 | 0.109 | 0.167 | 0.291 | |||||||
| PF/tallo (g) | LBF | 0.41 | b | 1.58 | b | 5.18 | a | 7.12 | a | 8.06 | b | |
| LBN | 0.43 | ab | 2.36 | a | 5.18 | a | 7.1 | a | 9.02 | a | ||
| LBC | 0.49 | a | 2.8 | a | 5.31 | a | 7.42 | a | 9.53 | a | ||
| DMSH | 0.068 | 0.598 | 1.086 | 0.963 | 0.878 | |||||||
| PS/tallo (g) | LBF | 0.03 | b | 0.04 | a | 0.18 | a | 0.34 | b | 0.5 | b | |
| LBN | 0.03 | b | 0.05 | a | 0.17 | a | 0.37 | ab | 0.56 | a | ||
| LBC | 0.04 | a | 0.05 | a | 0.22 | a | 0.39 | a | 0.58 | a | ||
| DMSH | 0.01 | 0.019 | 0.073 | 0.046 | 0.056 | |||||||
| PF/raíz (g) | LBF | 0.42 | b | 0.99 | b | 2.05 | b | 5.03 | b | 8.01 | b | |
| LBN | 0.55 | a | 1.21 | a | 2.07 | b | 5.33 | b | 8.59 | ab | ||
| LBC | 0.5 | a | 1.2 | a | 2.62 | a | 5.81 | a | 9 | a | ||
| DMSH | 0.058 | 0.12 | 0.15 | 0.429 | 0.712 | |||||||
| PS/raíz (g) | LBF | 0.018 | a | 0.065 | b | 0.085 | b | 0.213 | b | 0.341 | b | |
| LBN | 0.019 | a | 0.07 | ab | 0.105 | a | 0.263 | a | 0.42 | a | ||
| LBC | 0.017 | a | 0.075 | a | 0.1 | a | 0.256 | a | 0.413 | a | ||
| DMSH | 0.004 | 0.006 | 0.006 | 0.023 | 0.04 | |||||||
Medias con la misma letra son estadísticamente iguales (Tukey, p≤ 0.05). DMSH= diferencia mínima significativa honesta.
Los parámetros de luz roja (R= 122.04 µmol m-2 s-1; R:DFFF= 40.9%; R:A= 2.67) de las lámparas LBC provocaron que las plantas aumentaran el peso fresco de hojas, desde 3.9 hasta 15.4% en comparación con el efecto causado por las lámparas LBN (R= 100.44 µmol m-2 s-1; R:DFFF= 32.9%; R:A= 1.55) y desde 8.6 hasta 38% con relación a LBF (R= 81.34 µmol m-2 s-1; R:DFFF= 26.55%; R:A= 0.97). Con LBC también se incrementó el peso seco de hojas de 0 a 12.6% y de 8.9 a 27.4%, con respecto al obtenido de plantas cultivadas con LBN y LBF, respectivamente.
La respuesta morfológica influyó en las de crecimiento de planta, ya que la biomasa fresca y seca de hojas (parámetros de crecimiento) coincidieron estrechamente con el área foliar por planta (parámetro morfológico), acorde a lo observado por Hogewoning et al. (2010); Hernández y Kubota (2016). Estos autores indicaron que las plántulas de Cucumis sativus, cultivadas bajo combinaciones de luz roja y azul, requieren de 15 a 50% de luz azul para tener un crecimiento y desarrollo adecuados, ya que tales proporciones de luz se asociaron con aumentos en la biomasa por unidad de área foliar, contenido de nitrógeno y clorofila foliar, asimilación fotosintética de CO2 y conductancia estomática; las cuales fueron menores con luz roja monocromática donde las plantas mostraron un sistema de fotosíntesis disfuncional.
La acumulación de biomasa en el tallo de las plantas no presentó diferencias significativas (p≤ 0.05) durante los primeros 28 días después de la emergencia (dde). No obstante, a los 35 dde, la biomasa de tallo estuvo más relacionado con su longitud (altura de planta) que, con su diámetro, ya que los parámetros de luz roja de las lámparas LBC causaron incrementos de 5.7 y 18.2% en el peso fresco de tallo y de 0 y 12.2% en el peso seco de tallo, comparados con los obtenidos con las lámparas LBN y LBF, respectivamente; donde los tallos se alargaron menos y engrosaron más.
En este sentido, Ayala-Tafoya et al. (2015) observaron aumentos en el peso seco de hojas y tallo de plantas de Cucumis sativus cultivadas bajo malla roja, debido a la confluencia de más DFFF y luz roja, comparadas con las respuestas a la malla de color azul. De manera semejante, después de 21 dde, con las lámparas LBC el peso fresco de raíz aumentó de 4.7 a 26.7% y de 12.4 a 28.1%, en comparación con las lámparas LBN y LBF, respectivamente. Mientras que, con las lámparas LBC y LBN el peso seco de raíz superó de 17.6 a 21% al obtenido con las lámparas LBF.
Las plantas de pepino cultivadas con lámparas LBF presentaron una severidad de la cenicilla (Oidium sp.) de 0.8% a 10 días después de la inoculación (ddi) y de 3% a los12 ddi, la cual fue menor desde 4.9 y 5.3 veces hasta 3 y 4 veces comparadas con las plantas cultivadas con lámparas LBN y LBC, respectivamente. En el resto del estudio, las lámparas LBF indujeron los valores de severidad de 7.9% a 14 ddi y 62.5% a 35 ddi, menores desde 109.2 y 136.2% hasta 15.7 y 23.6%, en comparación con lámparas LBN y LBC, respectivamente. No obstante, los valores de severidad de la cenicilla obtenidos con los tres tipos de lámparas fluorescentes, durante todo el periodo de estudio, presentaron aumentos lineales con similares coeficientes de determinación (Figura 3).

Figura 3 Severidad de la cenicilla (Oidium sp.) en plantas de pepino ‘Poinsett 76’ cultivadas en cámara de crecimiento con lámparas fluorescentes de luz blanca fría (LBF), neutra (LBN) y cálida (LBC).
En el mismo sentido, otras investigaciones mostraron que la luz azul redujo la severidad de Botrytis cinerea en Solanum lycopersicum (Xu et al., 2017) y Podosphaera xanthii en Cucumis melo (Jing et al., 2018), al aumentar la expresión de genes relacionados con la defensa en las plantas, que indujo acumulación de prolina, H2O2, compuestos fenólicos, flavonoides, taninos y lignina, además de promover una morfología compacta y aumento del grosor de la pared celular en el tejido vegetal.
Conclusiones
Las lámparas fluorescentes compactas de luz blanca cálida promovieron mayor altura, área foliar, peso fresco y seco de hojas, tallo y raíz de las plantas. Mientras que, las lámparas fluorescentes compactas de luz blanca fría indujeron menor altura de planta y mayor grosor de tallo e índice de verdor foliar. Los parámetros espectrales de las lámparas fluorescentes compactas de luz blanca fría también indujeron menor severidad de la cenicilla en las plantas de pepino, en comparación con las lámparas de luz blanca neutra o cálida.

