










Services on Demand
Journal
Article
 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista mexicana de ciencias agrícolas
Print version ISSN 2007-0934
Rev. Mex. Cienc. Agríc vol.13 n.5 Texcoco Jun./Aug. 2022 Epub Oct 10, 2022
https://doi.org/10.29312/remexca.v13i5.2880
Articles
In vivo translocation of Klebsiella variicola PB02 and Klebsiella quasipneumoniae HPA43 in fruits of Solanum lycopersicum cultivar DT-22
1Laboratorio de Microbiología Molecular y Biotecnología Ambiental-Facultad de Ciencias Químico Biológicas-Universidad Autónoma de Guerrero. Av. Lázaro Cárdenas núm. 88, El Centenario, Chilpancingo de los Bravo, Guerrero, México. CP. 39000. Tel. 747 4725503.
2Campo Experimental Zacatepec-INIFAP. Carretera Zacatepec-Galeana s/n, Centro Zacatepec de Hidalgo, Morelos. CP. 62780. Tel. 734 3433820.
3Facultad de Ciencias Agropecuarias y Ambientales-Universidad Autónoma de Guerrero. Iguala-Teloloapan s/n, Ignacio Manuel Altamirano, Iguala de la Independencia, Guerrero. CP. 40040. Tel. 733 3334776.
4Facultad de Ciencias-Universidad Nacional Autónoma de México. Av. Universidad 3000, Circuito exterior s/n, Delegación Coyoacán, Ciudad de México, México. CP. 04510.
Currently, biofertilizers or organic compounds have been implemented in agriculture as fertilizer to increase the yield of crops of commercial interest and restore soil fertility. The use of plant growth-promoting bacteria has been one of the best alternatives that increase production yield and do not cause damage to the environment. Their use is not fully accepted by producers because of the risk they may have to the health of the consumer. Therefore, this work evaluated the effectiveness of two strains K. variicola PB02, K. quasipneumoniae HPA43 in consortium with Trichoderma in the production yield of Solanum lycopersicum cultivar DT22, a variety highly marketed in the national market, the significant increase (p= 0.003) was observed with respect to other commercial biofertilizers and the control group. The microbiological safety of the fruits, the translocation of these bacteria by the vascular system of the plant until reaching the fruit, was evaluated through the clonal profile of each of the isolates, without finding similarities between the inoculated strains and the strains isolated from endophytic tissues of the fruit, however it is necessary to carry out more genetically specific studies that verify that these bacteria have not actually developed mechanisms that allow them to translocate to aerial anatomical sites of plants and fruit, thus guaranteeing quality fruits without representing a risk to the health of the consumer and a high yield in the production of agricultural crops of commercial interest.
Keywords: biofertilizer; clonal profile; safety; translocation
En actualidad la agricultura se han implementado biofertilizantes o compuestos orgánicos como abono para aumentar el rendimiento de los cultivos de interés comercial y restaurar la fertilidad del suelo. El uso de bacterias promotoras de crecimiento vegetal ha sido una de las mejores alternativas que aumentan el rendimiento y no ocasionan daños al ambiente. Su uso no es totalmente aceptado por los productores por el riesgo que pudieran tener a la salud del consumidor. Por ello en este trabajo se evaluó la efectividad de dos cepas K. variicola PB02, K. quasipneumoniae HPA43 en consorcio con Trichoderma en el rendimiento de Solanum lycopersicum cultivar DT22 una variedad comercializada en el mercado nacional, se observó el aumento significativo (p= 0.003) respecto a otros biofertilizantes comerciales y al grupo testigo. Se evaluó la inocuidad microbiológica de los frutos, la traslocación de estas bacterias por el sistema vascular de la planta hasta llegar a fruto, por medio del perfil clonal de cada uno de los aislamientos, sin encontrar similitudes entre las cepas inoculadas y las cepas aisladas de tejidos endófitos del fruto; sin embargo, es necesario realizar estudios genéticamente para comprobar que estas bacterias no han desarrollado mecanismos que le permitan translocar a sitios anatómicos aéreos de las plantas y fruto, de esta manera garantizar frutos de calidad sin que representen un riesgo a la salud del consumidor y alto rendimiento en la producción de cultivos agrícolas de interés comercial.
Palabras clave: biofertilizante; inocuidad; perfil clonal; traslocación
Introduction
Bacteria associated with the rhizosphere, also called plant growth promoters (PGPB), fulfill specific functions in biogeochemical processes, interact with plants, establish protection against phytopathogens, increase nutrient availability and soil fertility level (Martínez-Romero et al., 2018). Among these species, the genus Klebsiella stands out, mainly the species of K. variicola and K. quasipneumoniae.
They fulfill mechanisms such as the modulation of the root system, promote positive phototropism, fulfill specific functions such as the production of indole-3-acetic acid, fix atmospheric nitrogen, solubilize phosphorus (Wei et al., 2013; Vega Celedón et al., 2016), which are essential for the formation of amino acids, nitrogenous compounds for metabolism and structural part of the plant, membrane phospholipid synthesis pathways, root elongation, flowering, part of the metabolism to obtain energy, repair and synthesis of fatty acids (Beltrán, 2014).
Another function of PGPB is to stimulate the development of the root system of the plant and therefore increase the absorption of nutrients, the main absorption pathway is through the root hairs present in the primary and secondary roots, through which a large number of organic compounds and molecules enter for the metabolism of the plant, it has been shown that there are microorganisms present in the soil that have developed the ability to internalize, translocate in the vascular system of the plant and persist in the fruit as is the case of E. coli O157: H7 (Heaton et al., 2008; Ocaña et al., 2018).
The interaction and metabolism with other microorganisms allow the association of microbial communities that establish symbiosis between bacteria with antagonist fungi that provide protection to the plants against phytopathogens present in the soil. Among the antagonist fungi present are Trichoderma, commonly isolated from the soil, being widely studied and marketed as biopesticides, biofertilizers and soil amendments, which allow the balance of the ecological niche (Benítez et al., 2004; Do Nascimento et al., 2017; Hirpara et al., 2017). Several strategies have been used to identify the molecular factors involved in this complex symbiosis interaction, including genomics, proteomics, and more recently, metabolomics, in order to improve our understanding (Vinale et al., 2008).
Biofertilizers based on microbial solutions have been evaluated in vegetables such as Solanum lycopersicum, commonly known as tomatoes (SIAP, 2019). However, it is of vital importance to monitor the internalization, persistence and translocation of these bacteria in the internal tissues of fruits in order to guarantee the health of the product. Therefore, the in vivo translocation of K. variicola and K. quasipneumoniae from the root system to the fruit of S. lycopersicum DT-22 in greenhouse was analyzed.
Materials and methods
Isolation of bacterial strains from various environments
Samples of banana, corn and tomato roots, and various water samples (river, lagoon, wastewater treatment plant and irrigation) were selected from the municipalities of Mochitlán, Acapulco, Chilpancingo and Tixtla, respectively. A sterile water wash of the root was carried out and the nodules were located. The rhizosphere samples were taken directly with a swab and transported to the laboratory in sterile 0.85% saline solution and in vials in tubes with MacConkey agar. Dilutions were made from the water samples up to 1x10-3, following the methodology of Puerta et al. (2010). After 24 h at room temperature, characteristic colonies of the genus Klebsiella, lactose-positive, irregular-edged and highly mucoid, dome-shaped, smooth and convex colonies were selected. Oxidase tests were performed, and the conventional biochemical profile was evaluated: in Kligler, MIO, citrate, urea, lysine to separate the genus.
Molecular differentiation of K. variicola, K. pneumoniae and K. quasipneumoniae by multiplex PCR
The biochemically identified strains of the genus Klebsiella, the extraction of DNA was performed by thermal shock, solubilizing three colonies in 50 μl of sterile water and subjecting to a temperature of 96 °C for 10 min and quickly in ice for 5 min and then centrifuged and 10 μl of the supernatant is taken and resuspended with 90 μl of sterile distilled water and kept at -20 °C. The reaction mixture for the multiplex PCR was carried out at a final volume of 25 μl, with a concentration of 1X of buffer, 2 mM of dNTP’s, 25 mM of MgCl, final of 1X of buffer, 2 mM of dNTP’s, 25 mM of MgCl, 1 μl of the oligonucleotides of K. variicola 2006 at a concentration of 10 pmol DNA, 2 μl of K. quasipneumoniae 10441 at a concentration of 5 pmol and 2 μl of the oligonucleotides of K. pneumoniae 13883 at a final concentration of 5 pmol, 0.5 μl of Taq DNA polymerase and 3 μl of the 1% DNA solution.
The reaction mixture was subject to an initial denaturation step of 2 min, at 92 °C, followed by 30 cycles of 30 s, at 92 °C, 30 s, at 60 °C, 40 s, at 72 °C and a final extension of 3 min, at 72 °C. The PCR products were subjected to electrophoresis in a 1.5% agarose gel at 120 V for 1 h. A fragment of 275 bp of K. variicola 2006, one of 372 bp K. quasipneumoniae 10 441 and one of 650 bp of K. pneumoniae 13 883 were used as controls (Martínez et al., 2018).
Detection of plant growth promotion mechanisms: nitrogen fixation
The strains were inoculated with a sterile wooden stick in modified Rennie medium free of N2 (Rennie, 1986) and incubated at 30 °C for 24-48 h. Growth is an indicator of nitrogen fixation. K. variicola PB was used as positive control and E. coli DH5-α was used as negative control.
Phosphate solubilization
The ability to solubilize inorganic phosphorus was evaluated in the National Botanical Research Institute’s Phosphate (NBRIP) medium with tricalcium phosphate following the methodology of Nautiyal (1999), the diameter of the halo and the colony (mm) was measured for the calculation of the solubilization index (SI), which relates the sum of the halo and colony diameters, and the diameter of the colony. K. variicola PB was used as positive control and E. coli DH5-α as was used negative control.
Quantification of indole acetic acid
The production of indole acetic acid (IAA) was evaluated in trypticasein soy broth supplemented with tryptophan following the methodology and interpretation of Lara et al. (2011). To quantify the production of IAA, in supernatants of the strains evaluated, a standard curve was made with different concentrations of commercial IAA (4 to 140 μg ml-1) and the Salkowski reagent in a 2:1 ratio. Absorbance was measured at 542 nm in a Stat Fax-2100 reader, using K. variicola PB as positive control and E. coli DH5-α as negative control.
Siderophore production
The production of siderophores was evaluated by growing the strains in F broth, added with glycerol 10 ml L-1, from a pure culture of 18-24 h, it was incubated for 24-72 h at 37 °C and the growth was examined under a faint ultraviolet light of 260 nm in a mini BioRad transilluminator. It was interpreted according to Villa et al. (2017), using K. variicola PB as positive control and F broth without inoculation as negative control.
Plant material and treatment design
The seeds of S. lycopersicum variety Saladette cultivar DT-22 were germinated in seedbeds of 54 cavities in December 2018, at an age of 39 days post-germination, they were transplanted in greenhouse without any fertilizer. For their study, a design of randomized complete blocks with seven plants each was carried out in accordance with NOM-032-SAG/FITO-2014, this for each treatment and distributed as shown in Figure 1, with a total of 252 plants in a greenhouse with an area of 500 m2, under controlled conditions of temperature, moisture and irrigation (the temperature was measured with a digital thermometer inside the greenhouse, when it reached 40 °C, corrective measures were taken, such as opening the wings of the greenhouse, the same for moisture, and irrigation was measured by two tensiometers that measure the effort made by the roots to extract moisture from the soil, one was 60 cm and another 30 cm from the soil and these turned on the drip irrigation when that of 30 cm exceeded 5 kPa (kilopascals) and the irrigation was turned off when that of 60 cm reached 0 kPa, all this was to avoid generating water stress in the plants). In addition, a strain of Trichoderma sp., X2, provided by Dr. Sergio Ramírez of INIFAP, Zacatepec, Morelos, was added in order to evaluate together with the strains of Klebsiella. It should be noted that T1 is the control group, T6 a commercial biofertilizer as positive control.
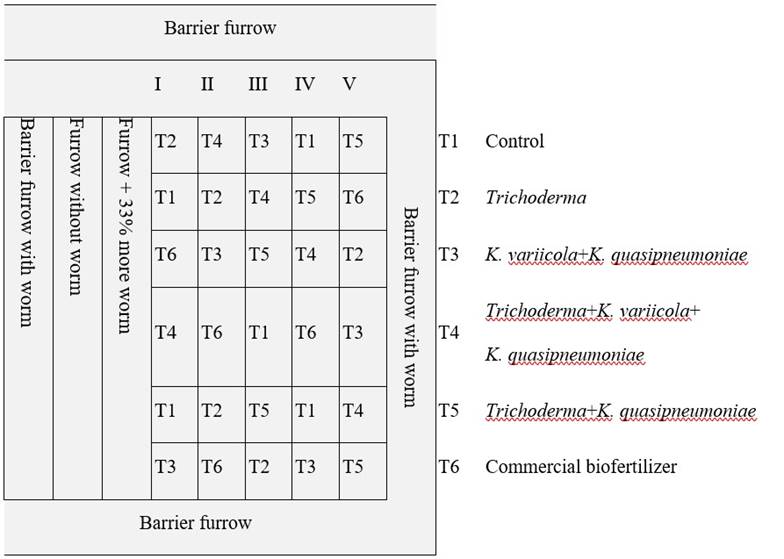
The tests of biological effectiveness and translocation to fruit were conducted during the agricultural cycle from January to June 2019. Environmental conditions were monitored throughout the cycle until the end of the production stage.
Preparation of microbial treatments
The selection of the strains PB02 and HPA43 used in this work was in accordance with their previous characterization as nitrogen-fixing, phosphate-solubilizing, indole-3-acetic acid-producing bacteria, molecularly identified by sequencing the rpoB gene as K. variicola and K. quasipneumoniae, respectively, and evaluated as root growth promoters in vitro in Solanum lycopersicum (Gutiérrez et al., 2017). As well as the results obtained when they were evaluated in vivo in other crops of commercial interest by this work team. A pre-inoculum of the strains was made individually in nutrient broth, adjusting an optical density of 1.5x108 microorganisms and the inoculum in saline solution at a final volume of 1.5 L per each treatment, at the same concentration of microorganisms.
Inoculation of microbial treatments in the root of S. lycopersicum cultivar DT-22
After transplantation, 50 ml of the inoculum was added with sterile syringe at the root level, this was to each of the plants by block, a total of 3 inoculations were performed for periods of every 30 days.
Production yield of S. lycopersicum fruits exposed to microbial treatments
The number of fruits produced was quantified and classified by class based on weight (g): those of first class weighed 100 g or more, second class from 50 to 99 g and third class with weight less than 49 g, the absolute weight (kg) of the production by block was also obtained with respect to each treatment; it is worth mentioning that this classification is carried out based on the experience of the producer; Eng. Rubén Ocampo Espín, who has produced and placed his product for more than 15 years in the national market. Absolute weight of production, weight by class (I, II and III) were analyzed as response variables. The statistical analysis was performed in the statistical program SAS version 9.4. A bifactorial Anova test, comparison of means by the Tukey test.
Microbiological safety of the fruits of S. lycopersicum cultivar DT-22 exposed to the various microbial treatments
Three fruits per treatment were randomly collected in three different events, with a total of 54 fruits per greenhouse. The collection and preparation of the samples was conducted executing the specifications of NOM-109-SSA1-1994. Once the fruits were transported to the laboratory, the surface of the fruits was disinfected with iodine at a concentration of 0.5% in sterile water for 5 minutes, after this, the peel of the fruit was removed.
For the processing of the samples, 10 g of the internal tissue of the fruit was taken and diluted in 90 ml of peptone H2O with sterile NaCl at 0.85%, the processing was carried out based on NOM-113-SSA1-1994 and NOM-210-SSA1-2014, from the resulting suspension, 100 μl were taken and inoculated in Petri dishes (culturable area 100 mm) in Mac Conkey agar, using the glass bead dispersion technique under sterility conditions for the count of CFU g-1 as indicated in NOM-110-SSA1-1994. For the interpretation of the result, the reference values established in NOM-113-SSA1-1994 were taken, which indicates that the maximum permissible limit (MPL) for anaerobic mesophiles is 150 000 CFU g-1 and for fecal coliforms is 100 000 CFU g-1 in food samples, together with ISO 21528-2:2017. The isolates were biochemically identified by the Api20E® Biomerix system following the manufacturer’s instructions.
Determination of the colonization of the root of S. lycopersicum DT-22 plants
Once the production stage finished, the S. lycopersicum plants were sacrificed to obtain the root, they were transported to the laboratory for their microbiological analysis, the root surface was disinfected with a 20% Tween solution for 10 min (modified from Bueno et al., 2007). One cubic centimeter was taken from the internal tissue of the apical meristem of the primary root. It was macerated with a sterile glass rod and diluted in 9 ml of peptone water with sterile 0.85% NaCl and vortexed at 3 500 rpm for 1 min. Subsequently, dilutions were made up to 10-8, in a time not exceeding 20 min, 100 μl of each dilution were plated in Mac Conkey agar plates in duplicate, incubating at 25-30 °C for 18 h. After the incubation time, the CFU g-1 count was performed. The isolates were identified by characteristic colonial morphology and biochemically.
Evaluation of the translocation by genomic fingerprint of each Klebsiella isolate
Genomic DNA was extracted from the total strains identified as Klebsiella by thermal shock and they were identified based on genetic patterns by PCR of enterobacterial repetitive intergenic consensus (ERIC) sequences to evidence their clonal origin. The oligonucleotides used were: ERIC-1 (5’-ATGTAAGCTCCTGGGGATTCAC-3’) and ERIC-2 (5’- AAGTAAGTGACTGGGGTGAGCG-3’), under the following conditions: initial denaturation at 95 °C for 3 min, followed by 30 cycles of 93 °C for 45 s, 50 °C for 1 min, 65 °C for 8 min and a final extension at 65 °C for 16 min (Versalovic et al., 1991). ERIC-PCR products were analyzed in 1% agarose gels at 75 V for 2 h and stained with ethidium bromide (3 μl 100 ml-1 of agarose) and visualized under UV light (365/302 nm). The genomic pattern of the strains isolated from the fruit and root were compared with the type strains of K. variicola PB02 and K. quasipneumoniae HPA43. Finally, the interpretation of the patterns obtained was carried out according to Tenover’s criteria (Tenover et al., 1995).
Results and discussion
A total of 13 strains isolated from the rhizosphere of various agricultural crops (banana, corn and tomato) and water were collected. The species of K. variicola, K. pneumoniae and K. quasipneumoniae were molecularly differentiated. Obtaining 37.5% (3/13) confirmed as K. pneumoniae, strains RS10, GN7 and KA2, 6.5% (1/13) K. variicola PB02, and 12.5% (2/13) identified as K. quasipneumoniae HPA4-3 and PB06. K. quasipneumoniae and K. variicola have had a great impact, however, a bad approach, lack of resources, ignorance of the diversity among species, lack of updating of the databases of the equipment used for bacterial identification in hospitals hinder their differentiation and diagnosis.
Therefore, it is important to use molecular methods to confirm the identification and differentiation of species within the microbiological routine, given that studies by Garza-Ramos et al. (2015) mention that it is not possible to differentiate K. quasipneumoniae and K. variicola from K. pneumoniae by biochemical and phenotypic tests, but by molecular methods (Figure 2).
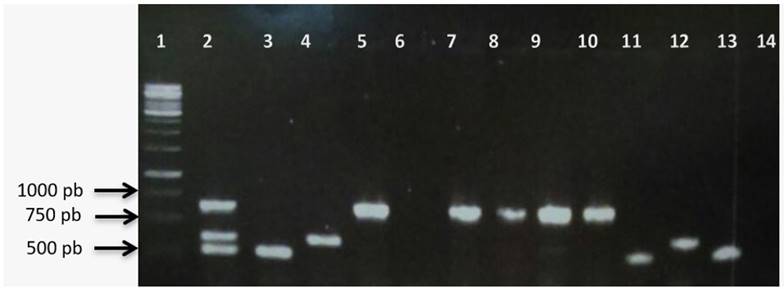
Figure 2 Differentiation of the strains of Klesiella sp. Lane 1= molecular weight marker 1kb DNA; lane 2= positive control; DNA K. variicola, K. quasipneumoniae, K. pneumoniae, lane 3= DNA K. variicola 2006 (control); lane 4= DNA K. quasipneumoniae 10442 (control); lane 5= DNA K. pneumoniae 13883 (control); lane 6= negative control (distilled water); lane 7= DNA strain KC1; lane 8= DNA strain KC2; lane 9= DNA strain KC3; lane 10= DNA strain RS10; lane 11= DNA strain PB02 (K. variicola); lane 12= DNA strain HPA4-3 (K. quasipneumoniae); lane 13= DNA strain PB06 (K. variicola).
Bernabé (2016) reports K. quasipneumoniae from the rhizosphere of Opuntia ficus identified by metagenomics directed at the 16S rRNA gene. This being the first report of K. quasipneumoniae in an environmental area, the HPA4-3 strain isolated from blue river water and the PB06 strain located in the corn rhizosphere. It is worth mentioning that 43.75% (7/13) of the isolates did not amplify for any species, but for the genus.
Biological nitrogen fixation
The strains identified as K. variicola PB02 and K. quasipneumoniae HPA4-3 are able to fix atmospheric nitrogen in 100%, since they present a nitrogenase complex activity at 24 h, evaluated in vitro. The latter is confirmed by the study of Fouts et al. (2008) that reports that K. pneumoniae has in its genome the regulon (KPK-1696-KPK-1715) of nitrogen fixation, indicating that the nitrogenase enzyme is present in the strains, which catalyzes the conversion of N2 to NH4. Carcaño et al. (2006) report Klebsiella sp. with a nitrogenase activity in a range of 42-122.46 nmol of ethylene per ml, where when compared with the genus Azospirullum sp. (8.62-70.08 nmol), it shows a significant difference in the nitrogenase activity of ethylene per ml, this indicates that the genus Klebsiella sp. has great potential as a nitrogen-fixing bacterium.
Phosphate solubilization
One hundred percent of the strains are solubilizers. The one with the highest index was K. quasipneumoniae PB06 with indices of 10.7 mm, followed by K. pneumoniae KA-2 with 9.66 mm and K. quasipneumoniae HPA-4-3 with 7.97 mm, Tereja et al. (2013) evaluated strains of Bacillus sp., where 29.31% are phosphate solubilizers, this places Klebsiella as a genus with great potential to perform this phenomenon. The release of soluble phosphorus is essential for plant development; it plays a fundamental role in DNA synthesis. Lara et al. (2011) reports a solubilization index with a maximum range between 3 and 4.2 mm in diameter of a total of 20 strains, among which are P. putida, P. luteola, E. sakasaki and E. cloacae, evaluated under the same conditions of this working document. The difference in these results is related to the ability to fix atmospheric nitrogen. High rates of solubilization have been recorded in the presence of ammonium as a nitrogen source, which is related to the percentage of fixation in the strains evaluated in our work (Beltrán, 2014).
Production of indole acetic acid
In the production of IAA, it was observed that 100% of the strains are producers of IAA, among which the following stand out: K. pneumoniae RS10 and K. quasipneumoniae PB06, with a high concentration of 3.241 and 2.738 μg ml-1, respectively, which is statistically significant with respect to the positive control K. variicola PB with 2.169 μg ml-1. The concentration of IAA in the plant is responsible for increasing the root system and foliage of the plant (Taiz-Zeiger, 2013). In addition, this phytohormone is related to the induction of stem elongation and fruit development.
The results obtained in this study, compared to those of Bautista-Gallardo (2008), are lower. They worked with A. vinelandii and obtained a high IAA production. Carcaño et al. (2006) report strains of Azospirillum and Klebsiella with a production of up to 26.55 μg ml-1, which confirms that species of the genus Klebsiella are producers of this auxin, however, in this study it can be compared that the strains of Klebsiella sp. that do not have nitrogenase activity have a higher production of indoles and vice versa, this confirms the results obtained given that, in the nitrogen-fixing strains, a production of IAA is obtained in low concentrations. Vega et al. (2016) mention that IAA-producing strains have a greater growth promoting effect, increasing the elongation of the primary root and the number of secondary roots. However, those that do not produce IAA delay root elongation.
Siderophore production
The production of siderophores was carried out on the molecularly identified strains: 100% of the strains evaluated are producers of siderophores. This agrees with what was mentioned by Arena et al. (2015), where they show that K. pneumoniae produces siderophores: enterobactin and aerobactin. This gives it competitive advantages in the colonization of the root, providing protection through the biocontrol that it establishes, due to the uptake of iron by siderophores, being attracted to the bacterial membrane, thus limiting the availability of iron for microorganisms that lack the assimilation system. This is consistent with what Molina et al. (2015) mentioned in relation to the antagonism between B. altitudinis and Thanatephorus cucumeris.
K. quasipneumoniae HPA4-3 and PB06 and K. variicola PB02 were selected as candidates together with Trichoderma sp. to be inoculated in S. lycopersicum Saladette cultivar DT-22, although the strain of K. pneumoniae RS10 shows several characteristics of plant promotion, it was ruled out because of the various published studies that demonstrate the alterations in human health.
Production yield in Saladette cultivar DT-22
A total of 11 harvests (one per week) were made, five harvests were taken at random in different events to analyze yield. The total estimated production harvest was: 1 392 kg for all treatments. The results obtained show that the treatments evaluated do not have the same effect on the production of first class tomatoes, the Anova comparative analysis gives us a value of p= 0.0008*, complete data are observed in Figure 3, where it is classified according to the Tukey grouping, treatments with the same letter do not have significant differences in their effect, T4 has the highest average value (p= 0.0007*) in production (39.618 kg) per harvest, with respect to the control group (T1). Unlike the treatments where they were evaluated individually, T2 does not show a significant difference with respect to the control (p= 0.464), T3 has a lower production (p= 0.999*) compared to the other treatments and the control (T1), on the other hand, T5 does not show a significant difference (p= 0.999 ) with respect to the control group (T1), the latter consortium was carried out because the HPA43 strain was able to promote in vitro root growth of S. lycopersicum L. (Gutiérrez et al., 2017).
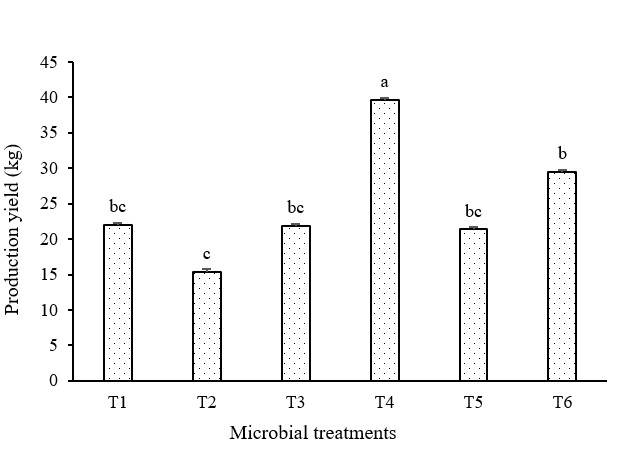
Figure 3 Graph of comparison of means of the production yield (kg) of Saladette tomato cultivar DT-22. The graphs show the standard error bar and the classification according to the Tukey grouping, which indicates that the treatments with the same letters do not present significant differences.
In addition to the microbial consortia, a commercial biofertilizer T6 was evaluated, already standardized and it was found that it does increase the production yield of the crops with respect to T1; however, it is not significant compared to our T4. These microorganisms are able to establish a mutualistic association (bacteria-fungus-plant) and potentiate the production yield, stimulate the development of the root system of the plant, influencing cell differentiation at the level of the meristem, internalizing themselves in the zone of differentiation, and they influence the differentiation of cells according to the needs of the plant and environmental conditions, therefore there is a greater number of root hairs which allow the entry of nutrients and minerals into the vascular system of the plant (Scharf et al., 2016; Calzavara et al., 2018).
Microbiological analysis of fruits exposed to microbial treatments
Table 1 shows the microbiological count of the fruits by treatment, the means and standard deviations of the Log of CFU g-1 of three repetitions in three different events, the number of microorganisms g-1 present in the sample is reported, as ISO 21528-2:2017, the values obtained from the count are within the maximum permissible limits.
Table 1 Microbiological count in fruits of S. lycopersicum DT-22 in each treatment.
| Treatment | Microbiological count Log of CFU g-1 (X ±SD) | N (microorganisms g-1) | Pr(>F) | Reference |
| T3 | 2.6 ±2.5 | 3262 a | p= 0.469 | NOM-113-SSA1-1994, ISO 4833-1:2013, ISO 21528-2:2017 |
| T4 | 2.2 ±2.1 | 1595 a | ||
| T5 | 2.5 ±2.5 | 2828 a | ||
| T1 | 2.2 ±2 | 1545 a |
X= mean; SD= standard deviation; N= number; T3= K. variicola and K. quasipneumoniae; T4= K. variicola, K. quasipneumoniae, Trichoderma sp.; T5= K. quasipneumoniae, Trichoderma sp.; T1= control. The subscript a in each value belongs to the Tukey grouping, equal letters indicate that there is no statistical difference between the treatments.
From the CFU g-1 count found, it is inferred that it is the endemic microflora that is associated with the anatomical sites of the fruit, which have affinity for specific sites of the plant according to the nutritional composition of each anatomical site (Ottesen et al., 2013). It is of vital importance the microbiological surveillance and the study of the microbial ecology of these crops of commercial interest, such as tomato DT-22, which is marketed directly to the market and is highly consumed fresh, because the disinfectants used are of superficial use in the fruit, they do not exert an effect on the internal tissues (Barak et al., 2012). We must watch that the fruits do not pose a risk to the health of the consumer.
Molecular analysis of genomic fingerprint of K. variicola PB02 and K. quasipneumoniae HPA43
A total of 114 strains (12 of fruit and 102 of root) were obtained, identified phenotypically and biochemically as Klebsiella sp. The profiles obtained based on ERIC-PCR allowed classifying clones (38) and subclones (16) of the strains of K. variicola PB02 and K. quasipneumoniae HPA43, all from root, of the strains isolated from fruits, no isolation was determined with the same pattern of bands similar to that of the control strains, as observed in Figure 4.
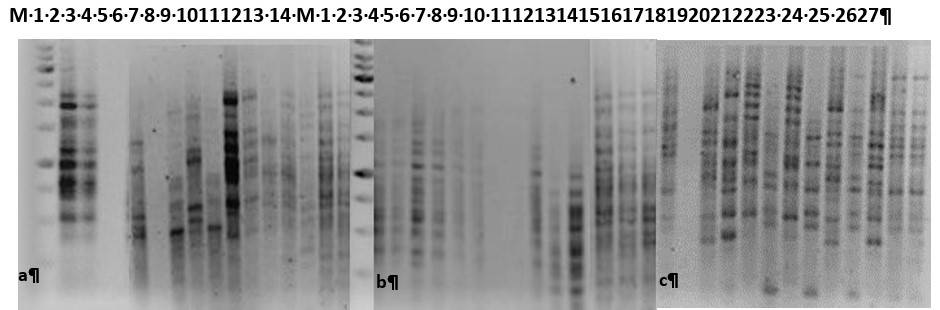
Figure 4 The clonality patterns of enterobacterial repetitive intergenic sequences are observed. M= molecular weight marker of 1Kb; 1= strain K. variicola PB02; 2= K. quasipneumoniae HPA43; 3= negative control. Fruit strains; 4= T5R3, 5= S2, 6= R4, 7= V4, 8= P3, 9= Z3, 10= T3R2, 11= R3, 12= N3. Root strains; 13= T5R5, 14= T5R1, 15= T3R4, b) M= molecular weight marker of 1Kb; 1= 3R1, 2= 3R2, 3= 3R4, 4= 3R4, 5= 3R5, 6= 5R1, 7= 5R2, 8= 5R3, 9= 5R4, 10= 5R5, 11= T5R3, 12= T5R4, 13= T3R2, 14= V5, 15= W5, 16= T5, 17= T3R1, 18= T3R4, 19= T3R5, 20= T3R, 21= 5R1, 22= A1, 23= A6, 24= B6, 25= C6, 26= D6, 27= E6.
This method has been used mainly to determine hospital outbreaks with the aim of verifying whether phenotypically similar isolates have a genetic relationship, given the high rate of genetic variability in prokaryotes caused by environmental factors, intergenic sequences conserved in enterobacteria that allow to identify by means of their fingerprint that they are derived from a common ancestor, in the same place and in a defined period, are amplified (Tenover et al., 1995). These profiles are useful for analyzing the spread and distribution of bacteria in the environment, how they move from one place to another and the conditions under which they persist.
In the results obtained, genetic similarity was only found between the Klebsiella strains isolated from the root and the strains K. variicola PB02, K. quasipneumoniae HPA43 inoculated; however, more specific molecular studies are needed to confirm that these bacteria or other PGPB do not actually translocate to the fruits, that they do not have integrated adaptation mechanisms by horizontal transfer that allow them translocation to the fruit, as is the case of E. coli, in response to environmental or genetic factors (Ottesen et al., 2013). To better understand the interaction of the microbial ecology present in the fruit and determine if the colonization of these bacteria is a determining factor in developing a health problem to consumers.
Conclusions
The use of plant growth-promoting bacteria in consortia with antagonist fungi potentiates the production yield of agricultural crops of commercial interest, helping to meet consumption demands and reducing production costs in a shorter time compared to conventional agriculture. There is a need for further studies on the effects of the amount of inoculum used on fruit yield and to use finer molecular tools of the strains isolated from the fruits to corroborate the clonal pattern and propose them with greater certainty for the study of translocation of PGPB from the rhizosphere to the fruits.
Acknowledgements
The participation of Dr. Yanet Romero Ramírez, Dr. Arturo Ramírez Peralta, Dr. Ulises Garza Ramos García, Q. Alejandro Sánchez and QBP. Brenda Celeste Rodríguez Chacón in the review and technical support of the project is appreciated
REFERENCES
Arena, F.; Henrici De Angelis, L.; Pieralli, F.; Di Pilato, V.; Giani, T.; Torricelli, F. and Rossolini, G. M. 2015. Draft genome sequence of the first Hypermucoviscous Klebsiella quasipneumoniae subsp. quasipneumoniae Isolate from a Bloodstream Infection. Genome Announc. 3(5):e00952-15. http://doi.org/10.1128/genomeA.00952-15. [ Links ]
Barak, D. J. and Schroeder, K. B. 2012. Interrelationships of food safety and plant pathology: the life cycle of human pathogens on plants. Annual Review Phytopathol. 50(1):241-266. DOI:10.1146/annurev-phyto-081211-172936. [ Links ]
Bautista, L. X. C. y Gallardo, I. R. 2008. Estandarización de métodos de detección para promotores de crecimiento vegetal (ácido indol acético y giberelinas) en cultivos microbianos. Tesis de grado. Pontificia Universidad Javeriana. 159 p. [ Links ]
Beltrán, M. E. 2014. Phosphate solubilization as a microbial strategy for promoting plant growth, Corpoica Cienc. Tecnol. Agropecuaria. 15(1):101-113. [ Links ]
Benítez, T.; Rincón, A. M.; Limón, M. C. y Codón, A. C. 2004. Biocontrol mechanisms of Trichoderma strains. International microbiology: the official J. Spanish Soc. Microbiol. 7(4):249-260. [ Links ]
Bernabé, S. L. 2016. Identificación molecular de bacterias cultivadas y no cultivadas asociadas a la rizósfera de Opuntia ficus-indica (L.) mil. (cactácea) en ecosistemas áridos. Tesis de maestría. Universidad Nacional de Tumbes. 31-33 pp. http://repositorio.untumbes.edu.pe/ handle/UNITUMBES/186. [ Links ]
Calzavara, A. K.; Godoy, P. P. H.; Cavanha, G. L.; Martínez, O. L. A.; Milanic, K.; Caixeta, O. H.; Bianchini, E.; Pimenta, J. A.; Neves, O. M. C.; Dias, P. J. and Stolf, M. R. 2018. Associative bacteria influence maize (Zea mays L.) growth, physiology, and root anatomy under different nitrogen levels. Plant Biol. 20(5):870-878. doi:10.1111/plb.12841. [ Links ]
Carcaño, M. M. G.; Ferrera, C. R.; Pérez, M. J.; Molina, J. D. and Bashan, Y. 2006. Nitrogenase activity, production of phytohormones, siderophores and antibiosis in strains of Azospirillum and Klebsiella isolated from Maize and Teosintle. Terra Latinoam. 24(4):493-502. [ Links ]
do Nascimento, M. O.; de Almeida, S. R.; dos Santos, G. R.; de Oliveira, C. A. and de Sousa, D. J. 2017. Antagonism of Trichoderma isolates against Leucoagaricus gongylophorus (Singer) Möller. J. Basic Microbiol. 57(8):699-704. doi: 10.1002/jobm.201600755. [ Links ]
Fouts, D. E.; Tyler, H. L.; Deboy, R. T.; Daugherty, S.; Ren, Q.; Jonathan, H. and Methe, B. A. 2008. Complete genome sequence of the N2 -fixing broad host range endophyte Klebsiella pneumoniae 342 and virulence predictions verified in mice. PLoS Genet. 4(7):e1000141. 1-8 pp. doi: 10.1371/journal.pgen.1000141. [ Links ]
Garza, R. U.; Silva, S. J.; Martínez, R. E.; Tinoco, P.; Pina, G. M. and Barrios, H. 2015. Development of a multiplex-PCR probe system for the proper identification of Klebsiella variicola. BMC Microbiol. 15(64):1-24. doi: 10.1186/s12866-015-0396-6. [ Links ]
Gutiérrez, I. G. y Nava, C. G. 2017. Evaluación de Klebsiella variicola, Klebsiella quasipneumoniae y Klebsiella pneumoniae como reguladoras de crecimiento radicular de Solanum lycopersicum. Tesis de Licenciatura. Facultad de Ciencias Químicas de la Universidad Autónoma de Guerrero. 22-45 pp. [ Links ]
Heaton, J. C. and Jones, K. 2008. Microbial contamination of fruit and vegetables and the behaviour of enteropathogens in the phyllosphere: a Review. J. Appl. Microbiol. 104(3):613-626. doi:10.1111/j.1365-2672.2007.03587. [ Links ]
Hirpara, D. G.; Gajera, H. P.; Hirapara, J. G. and Golakiya, B. A. 2017. Inhibition coefficient and molecular diversity of multi stress tolerant Trichoderma as potential biocontrol agent against Sclerotium rolfsii Sacc. Infection, genetics, and evolution. J. Mol. Epidemiol. Evol. Gen. Infectious Dis. 55(29):75-92. https://doi.org/10.1016/j.meegid.2017.08.029. [ Links ]
Lara-Celia, C.; Oviedo, L. y Alemán, A. 2011. Aislados nativos con potencial en la producción de Ácido Indol Acético para mejorar la agricultura. Biotecnología en el Sector Agropecuario y Agroindustrial. 9(1):17-23. [ Links ]
Martínez-Romero, E.; Rodríguez-Medina, N.; Beltrán-Rojel, M.; Toribio-Jiménez, J. and Garza-Ramos, U. 2018. Klebsiella variicola and Klebsiella quasipneumoniae with capacity to adapt to clinical and plant settings. Salud Pública de México. 60(1):29-40. https://doi.org/10.21149/8156. [ Links ]
Nautiyal-Shekhar, C. 1999. An efficient microbiological growth medium for screening phosphate solubilizing microorganisms. Microbiology Letters. 170:265-270. [ Links ]
Ottesen, A. R.; González-Peña, A.; White, J. R.; Pettengill, J. B.; Li, C.; Allard, S.; Rideout, S.; Allard, M.; Hill, T.; Evans, P.; Strain, E.; Musser, S.; Knight, R. and Brown, E. 2013. Baseline survey of the anatomical microbial ecology of an important food plant: Solanum lycopersicum (tomato). BMC Microbiology . 13(114):1-11. https://doi.org/10.1186/1471-2180-13-114. [ Links ]
Puerta-García, E. A. and Mateos-Rodríguez, F. 2010. Enterobacterias. Medicine. 10(1):3426-3431. [ Links ]
Rennie, R. J. 1986. Advantages and disadvantages of nitrogen-15 isotope dilution to quantify dinitrogen fixation in field-grown legumes. A critique. In: Hauck R. D. and Weaver, R. W. Field measurement of dinitrogen fixation and denitrification. Soil Science. Society of America, Madison, Wisconsin, USA. Special Publication 8. (Ed.). 43-58 pp. [ Links ]
Scharf, B. E.; Hynes, M. F. and Alexandre, G. M. 2016. Chemotaxis signaling systems in model beneficial plant-bacteria associations. Plant Mol. Biol. 90(6):549-559. https://doi.org/ 10.1007/s11103-016-0432-4. [ Links ]
Taiz, L. and Zeiger, E. 2013. Fisiologia Vegetal. 5th Edition. Artmed, Porto Alegre. 918 p. [ Links ]
Tereja-Hernández, R. B. 2013. Aislamiento de Bacillus solubilizadores de fosfatos. Agron. Mesoamer. 24(2):357-364. [ Links ]
Tenover, F. C.; Arbeit, R. D.; Goering, R. V.; Mickelsen, P. A.; Murray, B. E.; Persing, D. H. and Swaminathan, B. 1995. Interpreting chromosomal DNA restriction patterns produced by pulsed-field gel electrophoresis: criteria for bacterial strain typing. J. Clinical Microbiol. 33(9):2233-2239. https://doi.org/10.1128/JCM.33.9.2233-2239.1995. [ Links ]
Versalovic, J.; Koeuth, T. and Lupski, J. R. 1991. Distribution of repetitive DNA sequences in eubacteria and application to fingerprinting of bacterial genomes. Nucleic. Acids. Res. 19(24):6823-6831. [ Links ]
Vega-Celedón, P.; Canchignia-Martínez, H.; González, M. y Seeger, M. 2016. Biosíntesis de ácido indol-3-acético y promoción del crecimiento de plantas por bacterias. Cultivos Tropicales. 37(1):33-39. [ Links ]
Villa, P. M.; Frías, A. y González, G. 2017. Evaluación de cepas de Pseudomonas sp. para el control de hongos fitopatógenos que afectan cultivos de interés económico. ICIDCA. Sobre los derivados de la caña de azúcar. XXXIX(3): 40-44. [ Links ]
Vinale, F.; Sivasithamparam, K.; Ghisalberti, E. L.; Marra, R.; Woo, S. L. and Lorito, M. 2008. Trichoderma-plant-pathogen interactions. Soil Biol. Biochem. 40(1):1-10. https://doi.org/10.1016/j.soilbio.2007.07.002. [ Links ]
Wei, C.; Lin, L.; Luo, L.; Xing, Y.; Hu, C.; Yang, L.; Li, Y. and An, Q. 2013. Endophytic nitrogen fixing Klebsiella variicola strain DX120E promotes sugarcane growth. Biology and Fertility of Soils. 50(1):657-666. Doi: 10.1007/s00374-013-0878-3. [ Links ]
Received: March 01, 2022; Accepted: June 01, 2022
 Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
