











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
Para el crecimiento y desarrollo de los árboles frutales, los portainjertos juegan un papel importante en la absorción y transporte de agua y nutrimentos, así como la translocación a sus diferentes órganos (Fallahi et al., 2000), que está influenciado por la morfología y distribución espacial de las raíces (Gregory et al., 2013). Los portainjertos pueden ser selectivos en la absorción de nutrientes, por ello se deben seleccionar con cuidado para superar condiciones desfavorables (Fazio et al., 2013). En plantaciones de alta densidad, el requerimiento nutrimental difiere con respecto a las plantaciones convencionales debido a que los portainjertos enanizantes inducen precocidad, calidad del fruto y mayor rendimiento (Cheng y Raba, 2009) y controlan el vigor del árbol debido a su eficiencia para reducir o retrasar la competencia por el espacio disponible (Chalmers et al., 1981; Lo Bianco et al., 2003).
Los portainjertos M.27, M.9 y M.26, pueden reducir el área de la sección transversal del tronco y el volumen del dosel, con efectos positivos en la calidad del fruto en comparación con los portainjertos vigorosos (Autio et al., 1996; Fallahi et al., 2002).
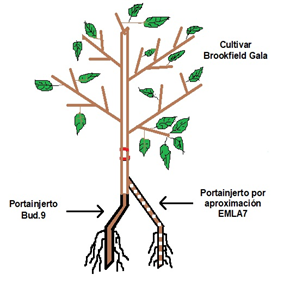
Los portainjertos enanizantes en condiciones de suelo y agua limitantes no siempre presentan la misma eficiencia con el cultivar injertado, debido a la deficiencia de nutrimentos. En el establecimiento de un huerto con un cultivar adaptado a las condicionen ambientales del lugar, pero no del portainjerto, se puede afectar seriamente el potencial productivo. Tal es el caso de un huerto en Aquixtla, Puebla, en el cual el cultivar Brookfield Gala está injertado en los portainjertos Bud.9 y Emla7. Los árboles en el primer portainjerto presentan deficiencias nutrimentales temporales y bajo vigor. Una alternativa para este problema es replantar con un portainjerto adecuado, sin embargo, ésta es costosa y significa pérdida de tiempo.
En árboles frutales se ha utilizado técnicas como el interinjerto para la resistencia de plagas y enfermedades o aumentar el rendimiento, con resultados positivos en algunos cultivares. Así también, el uso del injerto por aproximación en replantaciones con incidencias fitopatógenas se ha observado que puede mejorar la absorción de nutrimentos.
El injerto por aproximación consiste en plantar plántulas jóvenes de otro portainjerto junto a un árbol injertado y después injertarlas en el tronco de los árboles de la variedad (Bové y Ayres, 2007). También se ha utilizado con portainjertos enanizantes para mejorar el vigor de la variedad mediante el apoyo de portainjertos vigorosos (Hartmann et al., 2002). En condiciones deficientes de boro (B) en la naranja ‘Newhall’ esta técnica incrementa la concentración de B en el follaje y mejora el desarrollo de las raíces del árbol (Wang et al., 2016).
Por lo tanto, el objetivo de este estudio fue evaluar el estatus nutrimental y el vigor de manzano cv. Brookfield Gala injertado en el portainjerto Bud.9, apoyado con el portainjerto Emla7 por injerto de aproximación, en un suelo somero del Municipio de Aquixtla, Puebla. El portainjerto Emla7 fue seleccionado debido a que funciona bien con el cv Brookfield Gala, por lo que se espera mejore significativamente el estatus nutrimental y vigor de los árboles ‘Brookfield Gala’/Bud.9.
Materiales y métodos
Sitio experimental
El trabajo se realizó en los años 2018 y 2019 en un huerto comercial de manzano ubicado en el Municipio de Aquixtla, que se localiza en la parte Noreste del estado de Puebla, con coordenadas geográficas de 19°48’ 05” de latitud norte y 97° 56’ 32” de longitud oeste. El clima es templado subhúmedo con lluvias en verano (600-900 mm) y temperatura anual de 12-18 °C, a una altitud de 2 291 m (INEGI, 2009).
De acuerdo con WRB (2006) el suelo es un Tecnosol que se caracteriza por su material original de toba (tepetate) modificado por el agricultor, que mediante el subsoleo fue roturado y fragmentado y con la mezcla de suelo de otra procedencia, materia orgánica y abonos fue posible su aprovechamiento en la agricultura. El material de toba contiene 45-60% de óxido de silicio, por lo que hay mayor presencia de olivinos, anfíboles y piroxenos que se degradan fácilmente quedando disponibles sus elementos para la nutrición de las plantas.
En cuanto al estudio físico, el perfil del suelo mostró cinco estratos, a las profundidades de 0-18, 18-30, 30-41, 41-57 y 57-80 cm, respectivamente. Desde el segundo hasta el cuarto estrato está compuesto de tepetate fragmentado con materiales mezclados, compacto que no tiene espacios porosos. La textura es franco-arcillosa. A la profundidad de 0-30 cm se encontraron altas concentraciones de K, Mg, Fe y Cu y bajas concentraciones de N y Zn; 30-60 cm de profundidad altas concentraciones de Fe y Mn y bajos niveles en N, P, Ca, Zn y Na.
Material vegetal
El huerto se estableció en marzo de 2014 con el cultivar Brookfield Gala injertado en los portainjertos Bud.9 y Emla7 en un sistema de producción intensiva. El cultivar Brookfield Gala es una mutación natural del cultivar Royal Gala, originado en Nueva Zelanda. Bud.9, es un portainjerto enanizante, de bajo vigor con menor número de ramas y espolones cortos. Induce producción precoz en el segundo a tercer año y es resistente a plagas y enfermedades, así como a bajas temperaturas. El portainjerto Emla7, es semienano emite muchos retoños y espolones largos, también induce precocidad al tercer o cuarto año con alto rendimiento. Es tolerante a la pudrición del cuello, resistente a tizón de fuego y tolerante a sequía y frío.
Tratamientos y diseño experimental
Se definieron cuatro tratamientos en función del cultivar injertado en el portainjerto y al año en que se realizó el injerto de aproximación, de la siguiente manera: 1) ‘Brookfield Gala’/Bud.9; 2) ‘Brookfield Gala’/EMLA7; 3) ‘Brookfield Gala’/Bud.9+Emla7-2016; y 4) ‘Brookfield Gala’/Bud.9+Emla7-2017. Los cuatro tratamientos se distribuyeron en el campo de acuerdo con un diseño experimental de bloques completos al azar, con tres repeticiones. Los bloques se conformaron con relación al vigor de los árboles midiendo el diámetro (cm) del tronco (vigor bajo, medio y alto). En una densidad de 4 000 árboles ha, la unidad experimental fue un árbol. Como testigo se consideró el tratamiento 2; es decir, ‘Brookfield Gala’/Emla7, dado que presenta el mejor desempeño hortícola.
Manejo y conducción del experimento
Los árboles están establecidos a una distancia de plantación de 1 m entre plantas y 2.5 m entre hileras. Los árboles tenían una altura de 1.5 m en el portainjerto Bud.9 y 2.2.5 m en el portainjerto Emla7, están formados con el sistema de conducción de Tall Spindle, que tiene como objetivo aprovechar todas las ramas laterales para fructificación (Robinson et al., 2014).
El riego se realizó por goteo, la nutrición mediante fertirriego aplicando 40 kg nitrógeno y 60 kg de potasio por hectárea, una aplicación antes de la brotación vegetativa, y la aplicación de micronutrimentos vía foliar con la fórmula 20-20-20 y 0.5 kg de sulfato de magnesio en 100 L de agua, cada 20 días desde la brotación vegetativa hasta antes de la cosecha (aproximadamente 30 días). Además, se hacen dos aplicaciones de boro, al inicio de la apertura de las yemas vegetativas y en la apertura de yemas florales. La polinización se realiza con árboles del cv Granny Smith y colocación de ‘floreros’ con cultivares de la región.
Estatus nutrimental
Se determinó mediante el análisis foliar para lo cual se realizaron dos muestreos en el ciclo 2018; el primero a los 24 días después de plena floración (DDPF) y el segundo a los 106 DDPF; es decir, veinte días antes de la cosecha. Se obtuvo una muestra compuesta de hojas maduras de la parte media de la copa del árbol en brotes de espolones de crecimiento del año, cortando 30 hojas con peciolo de la parte este y 30 hojas del oeste. Las muestras se procesaron previo al análisis.
Las hojas se lavaron con agua corriente, destilada y desionizada, después se secaron en una estufa de convección (Imperial V Mechanical and gravity convention ovens Dubuque Iowa, EUA) a 68 °C durante 72 h. Posteriormente se molieron en un molino de acero inoxidable (General Electric Modelo AC 160, EUA). El nitrógeno (N) se determinó por el método Micro-Kjeldahl (Bremmer, 1965). La concentración de P, K, Ca, Mg, Fe, Mn, Zn, Cu y B se determinó por espectrometría de emisión atómica de inducción por plasma acoplado (ICP-OES Varian 725ES, Australia).
En el ciclo 2019, solamente se realizó el muestreo a los 24 DDPF mediante el mismo procedimiento del año 2018, justo cuando se presentaron las deficiencias temporales en los árboles, se determinaron los mismos nutrimentos con el procedimiento empleada en 2018.
Área de la sección transversal del tronco (ASTT)
Se determinó en los años 2018 y 2019, midiendo el diámetro (cm) del tronco del cultivar, 15 cm arriba del punto de unión entre el cultivar y el portainjerto de cada tratamiento: ‘Brookfield Gala’/Bud.9, ‘Brookfield Gala’/Emla7 y ‘Brookfield Gala’/Bud.9+Emla7 establecido en 2016 y 2017 (Figura 1), usando un vernier digital marca Truper Stainless y la fórmula ASTT= C2/416π, en la cual, ASTT= área de la sección transversal del tronco en cm2 y C= circunferencia del tronco en cm.
Resultados y discusión
Estatus nutrimental
En el año 2018, a los 24 días después de la plena floración (DDPF) la concentración de N, P, K, Ca, Mg, Zn, Mn y B no fue afectada significativamente por los tratamientos, pero sí la de Fe y Cu (Cuadro 1). Los resultados de las concentraciones de los nutrimentos se compararon con los valores del cv Gala/Emla7 reportado por Nachtigall y Dechen (2006) y el cv Gala/M.26 por Cheng y Raba, (2009), los cuales coinciden con las mismas fechas de muestreo. En la concentración de Fe, el tratamiento 2 (222.25 mg kg-1) fue superior con respecto al resto de los tratamientos cuya concentración varió de 156.33 a 170.65 mg kg-1.
Cuadro 1 Efecto del injerto de aproximación en la concentración mineral de macro y micronutrimentos en hojas jóvenes de árboles del manzano ‘Brookfield Gala’, 24 días después de plena floración en el año 2018.
| Tratamientos | N | P | K | Ca | Mg | Fe | Cu | Zn | Mn | B | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| (mg g-1) | (mg kg-1) | ||||||||||
| 1 | 27.3 a | 3.95 a | 12.1 a | 7.33 a | 2.9 a | 156.33 b | 9.5 aby | 20.03 a | 58.73 a | 76.2 a | |
| 2 (testigo) | 27.75 a | 3.7 a | 13.15 a | 6.95 a | 3 a | 222.25 a | 12.3 a | 16.95 a | 56.8 5a | 72.2 a | |
| 3 | 25.73 a | 3.63 a | 12.25 a | 6.96 a | 2.73 a | 170.65 b | 8.5 b | 19.06 a | 68.16 a | 73.63 a | |
| 4 | 25.15 a | 3.86 a | 11.13 a | 7.13 a | 2.76 a | 156.8 b | 8.43 b | 18.56 a | 64.9 a | 73.86 a | |
| DHS | 5.52 | 0.54 | 4.06 | 2.59 | 0.52 | 66.98 | 3.41 | 4.26 | 19.81 | 11.81 | |
y= diferentes letras dentro de las columnas indican diferencias significativas (p≤ 0.05). Tratamientos: 1) ‘Brookfield Gala’/Bud.9; 2) ‘Brookfield Gala’/Emla7; 3) ‘Brookfield Gala’/Bud.9+Emla7-2016; y 4) ‘Brookfield Gala’/Bud.9+Emla7-2017.
Esto indica que el portainjerto Emla7 es más eficiente en la absorción de hierro, y se debe a la mayor cantidad que raíces que tiene, lo cual permite una mayor exploración del suelo. Estos valores son superiores a los encontrados por Nachtigall y Dechen (2006); Cheng y Raba, (2009). La concentración de Cu, el tratamiento 2 (testigo) fue superior (12.3 mg kg-1) con respecto a los tratamientos 3 y 4 (8.43 y 8.5 mg kg-1, respectivamente en los años 2016 y 2017), el tratamiento 1 resultó con un valor intermedio (9.5 mg kg-1). Estos valores son superiores a los reportados por Nachtigall y Dechen (2006), lo que podría deberse a las aplicaciones foliares de micronutrimentos.
A los 106 días después de plena floración del mismo año, la concentración de N, P, K, Ca, Fe, Cu y Zn no fue afectada significativamente por los tratamientos, pero sí la de Mg, Mn y B (Cuadro 2). La concentración de magnesio, el tratamiento 2 fue superior (4.7 mg g-1) con respecto a los tratamientos 1 y 4 (3.83 y 3.86 mg g-1). El tratamiento 3 la concentración fue intermedia (4.03 mg g-1), lo cual refleja el efecto del año de realización del injerto por aproximación. Estos resultados son similares a los reportados en el cv Gala/Emla7 (Nachtigall y Dechen, 2006); sin embargo, en los tratamientos 1 y 4 los valores son menores, lo que puede sugerir que el portainjerto Bud.9 tiene baja eficiencia en la absorción de Mg.
Cuadro 2 Efecto del injerto de aproximación en la concentración mineral de macro y micronutrimentos en hojas maduras de árboles del manzano ‘Brookfield Gala’, 106 días después de plena floración en el año 2018.
| Tratamientos | N | P | K | Ca | Mg | Fe | Cu | Zn | Mn | B | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| (mg g-1) | (mg kg-1) | ||||||||||
| 1 | 22.83 a | 2.43 a | 10.63 a | 17.63 a | 3.83 by | 152.25a | 8 a | 17.8 a | 101.07 b | 63.8 ab | |
| 2 (testigo) | 24.6 a | 2.2 a | 10.8 a | 17.15 a | 4.7 a | 163.95 a | 9.9 a | 16.15 a | 98.35 b | 58.05 b | |
| 3 | 24.96 a | 2.2 a | 9.63 a | 17.6 a | 4.03 ab | 175.8 a | 7.4 a | 15.43 a | 153.5 a | 62.73 ab | |
| 4 | 24.46 a | 2.4 a | 10.53 a | 17.73 a | 3.86 b | 162.2 a | 7.4 a | 18.16 a | 105.4 b | 66.03 a | |
| DHS | 5.16 | 0.6 | 2.02 | 2.67 | 0.92 | 64.49 | 2.94 | 10.04 | 43.35 | 7.59 | |
y= diferentes letras dentro de las columnas indican diferencias significativas (p≤ 0.05). Tratamientos: 1) ‘Brookfield Gala’/Bud.9; 2) ‘Brookfield Gala’/Emla7; 3) ‘Brookfield Gala’/Bud.9+Emla7-2016; y 4) ‘Brookfield Gala’/Bud.9+Emla 7-2017.
La concentración de manganeso, el tratamiento 3 fue superior (153.5 mg kg-1) con respecto al resto de los tratamientos cuya concentración varió de 98.35 a 105.4 mg kg-1 respectivamente. Estos resultados indican que mediante el injerto por aproximación se puede incrementar la concentración de Mn en el dosel del árbol. Las concentraciones de estos tratamientos son superiores a los reportados por Cheng y Raba (2009), en el cv Gala/M.26; no obstante, en el cv Gala/Emla7, estos valores son inferiores con excepción del tratamiento 3 que se encuentra dentro del intervalo reportado.
En el caso del boro, el tratamiento 4 fue superior (66.03 mg kg-1) con respecto al tratamiento 2 (testigo) (58.05 mg kg-1), lo cual indica que no influyó el año en que se realizó el injerto por aproximación. Sin embargo, los tratamientos 1 y 3 tuvieron una concentración intermedia (63.8 y 62.73 mg kg-1), por lo que puede inferirse que el portainjerto Bud.9 es eficiente en la absorción de boro. Estas concentraciones de B son superiores a lo reportado en el cv Gala/Emla7 (Nachtigall y Dechen, 2006) y ‘Gala’/M.26 (Cheng y Raba, 2009).
Los resultados del análisis foliar indican niveles normales para N, P, Mg, Fe, Cu y Mn; minerales bajos para K y Zn y niveles altos para Ca y B (Jones et al., 1991). La deficiencia de potasio coincidió con la sintomatología observado en campo a 24 DDPF, de una coloración rojiza a púrpura con mayor visión en el envés de los brotes maduros y nuevos con desarrollo posterior de márgenes necróticos, que coincide con lo reportado por Burrell y Boynton (1943); Basile et al., (2003).
Los niveles bajos de potasio repercuten en el desarrollo y crecimiento, dado que la concentración inicial en hojas jóvenes es más alta que en hojas viejas (Huber, 1985), así también disminuye a medida que avanza su fenología y la extracción por el fruto (Faust, 1989; Brown, 1994). La deficiencia de potasio afecta la translocación de carbohidratos y la regulación en las relaciones planta-agua (Grace et al., 2012). Mpelasoka et al. (2003) mencionan que las variaciones en el contenido de K foliar pueden deberse a la capacidad de absorción de las raíces y variaciones en la incorporación de iones de K en el xilema y su translocación de la raíz hacia los brotes. Así también, esta deficiencia podría ser debido al antagonismo con el boro, como se reporta en olivo (Olea europea) (Chatzissavvidis et al., 2005).
La deficiencia de zinc se debe al bajo contenido en el suelo. Swietlik (2002) menciona que la deficiencia de zinc ocurre en gran medida porque en el suelo no se encuentra disponible o está inmóvil. La concentración elevada de calcio puede explicarse por la acumulación de acuerdo con la etapa fenológica de las hojas, así también ésta alta concentración de Ca puede explicar la deficiencia de potasio, dado que el calcio bloquea al potasio causando el antagonismo Ca/K.
Estos resultados no se relacionan con el contenido de Ca en el suelo, debido a que los análisis mostraron contenidos de medio a moderadamente bajos y en los estratos del perfil la presencia de CaCO3 fue de mediana a baja. Sin embargo, Fallahi et al. (2001), también encontraron altas concentraciones de Ca con el portainjerto Bud.9 que en EMLA7. La importancia del Ca es por la función que tiene en el alargamiento y división celular (Bush, 1995).
La alta concentración de boro puede deberse a las dos aplicaciones que se realizan durante la apertura de yemas vegetativas y florales, aunado a las aplicaciones foliares de micronutrimentos. Así también, la concentración en las plantas está influenciada por los portainjertos utilizados (Wojcik, 2003). El boro juega un papel importante en la transpiración, ya que es transportado a través del xilema a los órganos superiores (Paparnakis et al., 2013). Wojcik et al. (2003), encontraron altas concentraciones de B con portainjertos M.9 y M.26, similares a lo encontrado en los portainjertos de este estudio. Así también, la alta concentración puede deberse al pH del suelo que es moderadamente ácido (Paparnakis et al., 2013).
La deficiencia o exceso de B suele reducir la carga de cosecha como se observó en la reducción del porcentaje de flores en olivo (Chatzissavvidis et al., 2005), además de la sincronización con la polinización. Los síntomas de exceso de boro se reflejan en el bajo vigor de los árboles, desarrollo retardado y reducción en el número y peso de los frutos (Reid y Fitzpatrick, 2009; Herrera-Rodríguez et al., 2010), así como la disminución de la clorofila en la hoja, alteración del metabolismo y la necrosis de los tejidos maduros (Reid et al., 2004; Paparnakis et al., 2013).
La concentración de N, P y K a los 24 DDPF disminuyó a los 106 DDPF, mientras que la de Ca, Mg y Mn aumentó. Este resultado coincide con lo reportado en el cv Gala/EMLA7 en fechas similares de muestreo (Nachtigall y Dechen, 2006); sin embargo, la concentración de los elementos Fe, Cu, Zn y B disminuyó. Nachtigall y Dechen (2006); Mengel y Kirby (2001), mencionan que las hojas jóvenes suelen presentar mayores concentraciones de N, P y K y las hojas maduras tienen mayores concentraciones de Ca, Mn, Fe y B, principalmente. La disminución en la concentración de nutrimentos a lo largo del ciclo vegetativo, pueden estar relacionadas con un efecto de dilución que se produce en las hojas y la redistribución hacia otros órganos del árbol.
En el año 2019, a los 24 días después de plena floración, la concentración de N, K, Ca, Fe, Zn y Mn no fue afectada significativamente por los tratamientos, pero sí de los elementos P, Mg, Cu y B (Cuadro 3). La concentración de P fue superior en los tratamientos 1 y 4 (2.38 y 2.33 mg g-1) con respecto al tratamiento 2 (2.07 mg g-1), lo cual indica el efecto que tiene el portainjerto Bud.9 en la concentración de P, aunque también se observa el efecto del injerto por aproximación sin importar el año de su aplicación. La concentración de P en este estudio es superior a lo reportado en el cv Gala/Emla7 (Nachtigall y Dechen, 2006) e inferiores con respecto al cv Gala/M.26 (Cheng y Raba, 2009).
Cuadro 3 Efecto del injerto de aproximación en la composición mineral de macro y micronutrimentos en hojas jóvenes de árboles del manzano ‘Brookfield Gala’, 24 días después de plena floración en el año 2019.
| Tratamientos | N | P | K | Ca | Mg | Fe | Cu | Zn | Mn | B | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| (mg g-1) | (mg kg-1) | ||||||||||
| 1 | 22.15 a | 2.38 a | 10.61 a | 3.7 a | 1.56 by | 64.9 a | 6.8 b | 7.85 a | 30.71 a | 39.33 a | |
| 2 (testigo) | 20.35 a | 2.07 b | 11 a | 3.47 a | 1.87 a | 67.75 a | 6.46 b | 8.95a | 34.32 a | 26.2 b | |
| 3 | 23.11 a | 2.26 ab | 9 a | 3.36 a | 1.63 b | 72.2 a | 5.13 b | 7.98 a | 34.2a | 30.84 ab | |
| 4 | 22.26 a | 2.33 a | 10.76 a | 3.46 a | 1.63 b | 68.38 a | 11.86 a | 8.33 a | 33.05 a | 35.38 a | |
| DHS | 3.62 | 0.25 | 2.46 | 0.53 | 0.16 | 11.63 | 3.62 | 2.57 | 5.81 | 12.99 | |
y= diferentes letras dentro de las columnas indican diferencias significativas (p≤ 0.05). Tratamientos: 1) ‘Brookfield Gala’/Bud.9; 2) ‘Brookfield Gala’/Emla7; 3) ‘Brookfield Gala’/Bud.9+Emla7-2016; y 4) ‘Brookfield Gala’/Bud.9+Emla7-2017.
La concentración de Mg fue superior en el tratamiento 2 (1.87 mg g-1) con respecto al resto de los tratamientos, que varió de 1.56 a 1.63 mg g-1. Por lo tanto, el portainjerto Emla7 influyó en la concentración de magnesio. Sin embargo, estos valores son inferiores a lo reportado en el cultivar Gala/Emla7 (Nachtigall y Dechen, 2006) y en el cv Gala/M.26 (Cheng y Raba, 2009). En el cv BC-2 Fuji/Emla7, también se reportan altas concentraciones y bajos con el portainjerto Bud.9, lo cual puede deberse a la baja fotosíntesis neta (Fallahi et al., 2001).
La concentración de Cu, en el tratamiento 4 fue superior (11.86 mg kg-1) al resto de los tratamientos (5.13 a 6.8 mg kg-1). Estos valores son inferiores a lo reportado en el cv Gala/Emla7 (Nachtigall y Dechen, 2006).
Respecto al B, los tratamientos 1 y 3 (39.33 y 35.38 mg kg-1) fueron superiores al tratamiento 2 (26.2 mg kg-1), lo que podría explicar que el portainjerto Bud.9 absorbe y concentra mayor cantidad de boro, así como con el injerto por aproximación. En el cv Gala/M.26, se ha reportado concentraciones menores (Nachtigall y Dechen, 2006).
La concentración de nutrimentos refleja los factores que influencian la disponibilidad y suministro del suelo, así como la variación del clima y la carga de cosecha (Faust, 1989). La deficiencia de nutrimentos limita el crecimiento y desarrollo en la fructificación y con ello la reducción de la calidad, mientras que concentraciones altas producen síntomas de toxicidad (Hoying et al., 2004).
Así también, la absorción de nutrimentos por los portainjertos puede explicarse por las diferencias en la distribución y función de las raíces y las posibles diferencias en la anatomía de raíces y tallo que afectan la tasa de absorción y el movimiento hacia el xilema y las hojas (Zarrouk et al., 2005).
Área de la sección transversal del tronco (ASTT)
En el año 2018, el área de la sección transversal del tronco fue superior en el tratamiento 2 (12.8 cm2) con respecto al resto de los tratamientos que varió de 6.9 a 7.6 cm2, respectivamente. Esto refleja el vigor del portainjerto Emla7. En el año 2019, los resultados fueron similares, el tratamiento 2 (13.6 cm2) superó al resto de los tratamientos (Cuadro 4). Los tratamientos con menor ASTT podría explicarse por el menor tamaño y área foliar que corresponde al vigor de los árboles.
Cuadro 4 Área de la sección transversal del manzano ‘Brookfield Gala’ en los años 2018, 2019.
| Tratamientos | ASTT (cm2) | ||
|---|---|---|---|
| 2018 | 2019 | Incremento | |
| 1 | 7.6 b | 8.51 b | 0.91 |
| 2 (testigo) | 12.8 a | 13.6 a | 0.8 |
| 3 | 6.9 b | 8.22 b | 1.32 |
| 4 | 7.2 b | 8.27 b | 1.07 |
| DHS | 2.59 | 2.76 | |
y= diferentes letras dentro de las columnas indican diferencias significativas (p≤ 0.05). Tratamientos: 1) ‘Brookfield Gala’/Bud.9; 2) ‘Brookfield Gala’/Emla7; 3) ‘Brookfield Gala’/Bud.9+Emla7-2016; y 4) ‘Brookfield Gala’/Bud.9+Emla 7-2017.
El incremento en crecimiento de un ciclo a otro en las combinaciones sin injerto de aproximación fue menor de 1 cm; sin embargo, en los otros con injerto de aproximación de 2016 y 2017 fueron mayor a 1 cm. Aunque es mínima, refleja efecto del año de realización del injerto de aproximación.
En tanto, el acumulativo del ASTT presentó significancia en los tratamientos, siendo superior el tratamiento 2 (25.36 cm2) con respecto a los otros. Estos resultados se relacionan con lo que se ha reportado en varios cultivares, a medida que aumenta el vigor de los portainjertos aumenta el rendimiento acumulativo/árbol, pero disminuye la eficiencia de rendimiento acumulativo (Barrit et al., 1995). Webster y Wetheim (2003), indican que los portainjertos enanos son más eficientes que los portainjertos vigorosos porque inducen el tamaño de los frutos de la variedad.
Conclusiones
La respuesta del estatus nutrimental y el vigor del árbol, con el injerto por aproximación para mejorar el desempeño hortícola de los árboles de manzano injertadas en Bud.9, el efecto es incipiente, aunque muestra una tendencia a la mejora, lo que se puede atribuir al año de su establecimiento basados en el estatus nutrimental y área de la sección transversal del tronco, así como en el rendimiento (datos no publicados).
Con base al grado de uso del portainjerto Bud.9 en plantaciones intensivas, en condiciones de un suelo toba fragmentado su comportamiento limita el crecimiento y desarrollo del cultivar. Sin embargo, las prácticas culturales como la incorporación de materia orgánica, humus, riego constante y fertilización pueden contribuir a la mejora hortícola de los árboles.

