










Servicios Personalizados
Revista
Articulo
 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de ciencias agrícolas
versión impresa ISSN 2007-0934
Rev. Mex. Cienc. Agríc vol.11 no.spe24 Texcoco abr./may. 2020 Epub 07-Mayo-2021
https://doi.org/10.29312/remexca.v0i24.2360
Articles
Water relations and gas exchange in lucerne under drought conditions
1Colegio de Postgraduados-Campus Montecillo. Carretera México-Texcoco km 36.5, Montecillo, Texcoco, Estado de México, México. CP. 56230. Tel. 595 9520200, ext. 1587. (luna.milton@colpos.mx; sergiomp@colpos.mx; queroadrian@colpos.mx; haro@colpos.mx; meoc@colpos.mx.
2Universidad Autónoma Chapingo. Chapingo, Texcoco, Estado de México, México. CP. 56230. Tel. 595 9521500, ext. 5093. (pedroarturo@correo.chapingo.mx).
The effect of the edaphic water deficit on the water relations and gas exchange of ten varieties of lucerne (M. sativa L.) in the greenhouse with irrigation (I) and drought (D) were studied. A randomized complete block design was used with four replications in I and four in D. The experimental unit was an individual plant in a 4” PVC tube and 1 m high. The seedlings were transplanted into the tubes, 20 d after germination in storage trays. Fertilization was done by applying formula 60-140-00 at 44, 240 and 420 dds. In I the soil water content remained close to CC [20-406 ddt, I1 and 406-688 ddt, I2] and in S the application of water was suspended for 61 d (345-406 ddt, D1) and 68 d (620-688 ddt, D2). The lowest water potential (ψ), osmotic (π) and turgidity (p) and the rate of assimilation (A) and transpiration (E), and the lowest stomatal conductance (g) in I1 were ψ= -3.1, π= -3.5 and p= 0.4 MPa; A= 4.4 µmol CO2 m-2 s-1, E= 0.022 mol H2O m-2 s-1 and g= 0.7 mmol H2O m-2 s-1), and in S2 ψ = -3.5, π = -3.7 and p= 0.2 MPa; A = 4 µmol CO2 m-2 s-1, E= 0.019 mol H2O m-2 s-1 and g= 0.7 mmol H2O m-2 s-1. The osmotic adjustment (OA) and the efficiency in the use of water (W) showed final values of 1.7 MPa and 0.0058 mmol CO2 mmol-1 H2O-1 in S1 and 1.96 MPa and 0.0061 mmol CO2 mmol-1 H2O-1 in S2. The Genex, Jupiter, Atlixco and Milenia varieties with high levels of OA, A, E, g and W showed better behavior and tolerance to water stress.
Keywords: assimilation; efficiency in the use of water; osmotic adjustment; transpiration
Se estudió el efecto del déficit hídrico edáfico en las relaciones hídricas e intercambio gaseoso de diez variedades de alfalfa (M. sativa L.) en invernadero con riego (R) y sequía (S). Se utilizó un diseño de bloques completos al azar con cuatro repeticiones en R y cuatro en S. La unidad experimental fue una planta individual en un tubo de PVC de 4” y 1 m de alto. Las plántulas se trasplantaron en los tubos, 20 d después de la germinación en charolas para almacigo. La fertilización se hizo al aplicar la fórmula 60-140-00 a los 44, 240 y 420 dds. En R el contenido hídrico del suelo se mantuvo cercano a CC [20-406 ddt, R1 y 406-688 ddt, R2] y en S la aplicación de agua se suspendió por 61 d (345-406 ddt, S1) y 68 d (620-688 ddt, S2). El potencial hídrico (ψ), osmótico (π) y de turgencia (p) y la tasa de asimilación (A) y transpiración (E), y la conductancia estomática (g) más bajos en S1 fueron ψ= -3.1, π= -3.5 y p= 0.4 MPa; A= 4.4 µmol CO2 m-2 s-1, E= 0.022 mol H2O m-2 s-1 y g= 0.7 mmol H2O m-2 s-1), y en S2 ψ = -3.5, π = -3.7 y p= 0.2 MPa; A = 4 µmol CO2 m-2 s-1, E= 0.019 mol H2O m-2 s-1 y g= 0.7 mmol H2O m-2 s-1. El ajuste osmótico (AO) y la eficiencia en el uso del agua (W) mostraron valores finales de 1.7 MPa y 0.0058 mmol CO2 mmol-1 H2O-1 en S1 y 1.96 MPa y 0.0061 mmol CO2 mmol-1 H2O-1 en S2. Las variedades Genex, Júpiter, Atlixco y Milenia con altos niveles de AO, A, E, g y W mostraron mejor comportamiento y tolerancia al estrés hídrico.
Palabras clave: ajuste osmótico; asimilación; eficiencia en el uso del agua; transpiración
Introduction
The frequent presence of periods of water deficit in the soil during the lucerne life cycle and the increase in global atmospheric temperature, due to climate change, can cause severe reductions in the quality and yield of dry matter. Also, the high cost and the increasing scarcity of quality water for irrigation make forage production increasingly difficult in large areas of Mexico and the world (Orloff, 2015). In dry years when irrigation availability is low, lucerne is the first crop exposed to water deficiencies, considering that it is a crop with high water demand. However, lucerne can survive short periods of drought, without irrigation and can recover, and produce yield normally in subsequent years (Orloff, 2015).
Under these conditions the growth of the plants depends to a great extent on the gas exchange, which involves the loss of water from the tissue (transpiration) and the entry of CO2 into the interior of the leaf (photosynthesis). This exchange is regulated by stomatal conductance, so that to achieve greater efficiency in the use of water (W), when it is scarce, plants must reduce their stomatal conductance more to the loss of water than to the assimilation of CO2 (López-Castañeda, 1993).
The hydric status of the plant is closely related to the transpiration rate and the processes of water flow in the vascular system of the plant, in such a way that a hydric deficit in the soil can cause a reduction in stomatal conductance and the expansion of the vegetable canopy, and a rearrangement of the leaves in a more vertical position (Sim et al., 2017). The water deficit will be more serious with the increase in temperature in the future, putting crop yields under greater threat (Harrison et al., 2014).
For this reason, the development of varieties with greater resistance to drought and heat is a promising alternative in the selection of genotypes better adapted to stress conditions. Likewise, the discovery of the genetic, physiological and molecular bases of stress resistance mechanisms will be an indispensable tool for the design of new crops and varieties tolerant to drought and heat (Zhao et al., 2016; Zhang et al., 2019).
The active accumulation of solutes in the cell in response to the water deficit is reflected in a decrease in the osmotic potential of the cell, due to the increase in the concentration of solutes, such as protective osmotic compounds, antioxidants and growth regulators (Fahad et al., 2015). These cell-produced solutes contribute to plants' adaptation to environmental stress by mastering the activity of multiple stress response genes that exhibit attractive possibilities for the application of molecular breeding (Morran et al., 2011).
In addition, the synthesis of aquaporins and dehydrins have been reported as factors involved in drought tolerance of numerous species (Perdiguero et al., 2015). Plants' response to water deficit can help to exhibit the genetic-physiological mechanisms that determine resistance to drought. This study was carried out with the objective of evaluating the phenotypic response of 10 commercial varieties of lucerne in their water and gas exchange relationships in irrigation and drought under greenhouse conditions.
Materials and methods
Location
The study was carried out under greenhouse conditions in the drought resistance area of the Postgraduate College in Montecillo, Texcoco, State of Mexico (19° 29’ north latitude, 98° 53’ west longitude and altitude of 2 250 m).
Vegetal material
The experiment included 10 varieties of lucerne obtained from different commercial houses. Four groups of 100 seeds in each variety were chosen at random and germinated in Petri dishes with Whatman® brand filter paper, under laboratory conditions (temperature of 20 °C). The germination percentage ranged from 95 to 98% for all varieties.
Experimental design and soil moisture treatments
A randomized complete block experimental design with four replications and two soil moisture treatments (irrigation and drought) was used; the blocks were rotated inside the greenhouse every week to reduce the effect of the microenvironment on the plants. The experimental unit was an individual plant grown in a 1m high and 4” diameter PVC tube, with a 1.1 m high cylindrical poly bag and the same diameter as the PVC tube.
The sowing was carried out in boxes for storage, on March 15, 2017, by placing five seeds of the same weight in each cell; when the seedlings were 20 days after sowing (dds), the most vigorous seedling in each cell was chosen to be transplanted into the PVC tubes.
The PVC tubes were filled with soil dried in the sun for two weeks; the soil used is of a sandy loam texture with an apparent density of 1.12 T m-3, field capacity (CC) of 41.6% and percentage of permanent wilting (PMP) of 28.2% (Central University Laboratory, Autonomous University Chapingo, Texcco, Mexico, 2016). Fertilization was done 44 days after transplantation (ddt) with the formula 60-140-00, using urea and triple calcium superphosphate as sources of nitrogen and phosphorus; a second and third fertilization were done with the same formula at 240 and 420 ddt.
The soil moisture treatments were: irrigation, where the soil water content remained close to CC from the transplant date (20 dds) to 406 ddt (I1) and from 406 ddt to the conclusion of the experiment (688 ddt) (I2) and drought, where the application of water was suspended for a period of 61 days [345 to 406 ddt, March to May 2018; (D1)] and a second 68-day period [620-688 ddt; November 2018 to February 2019; (D2)].
Measured and calculated variables
The foliar water potential (ψ), osmotic (π) and turgidity (P), the photosynthesis rate (A), transpiration rate (E) and stomatal conductance (g), were measured at 345, 360, 377, 391 and 406 ddt in all plants in irrigation (I1) and drought (D1), and at 620, 635, 658, 673 and 688 ddt in I2 and S2.
ψ (MPa) was measured on a complete stem with the upper leaflets fully expanded and exposed to solar radiation, between 11:00 and 13:00 h, using the pressure pump or Scholander pump (Barrios-Gómez et al., 2011).
The π (MPa) was measured in the samples of the leaflets used to measure the ψ; the leaflets were placed in 5 ml plastic microtubes, which were placed in a thermos with liquid nitrogen at a temperature of -195 °C for 5 minutes. The π was determined in a 100 µl aliquot of each sample, using a Typ 6M 10/25 µl model osmometer, Löser brand (Germany) in the laboratory. The P (MPa) was calculated as the difference between the potentials ψ and π (p= ψ - π), for each sample analyzed (Begg and Turner, 1976). The osmotic adjustment (OA, MPa) of the plants was calculated as the difference between π determined in irrigation (πI) and π measured in drought (πD), for each variety (OA = πI - πD) Morgan (1995).
The net photosynthesis rate (A, µmol CO2 m-2 s-1), the transpiration rate (E, mmol H2O m-2 s-1) and the stomatal conductance (gi, mol H2O m-2 s-1) were measured by placing a fully expanded sheet inside the assimilation chamber of the infrared gas analyzer (IRGA, model LI-6400, LI-COR Inc; Lincoln, Nebraska, USA), configured for an open system, where measurements they are based on the differences in the concentration of CO2 and H2O in the leaf and the concentration of CO2 and H2O in a stream of air flowing through the leaf. The transpiratory efficiency or efficiency in the use of the instantaneous water of the entire plant, measured by the gas exchange (W, mol CO2 mol H2O-1) was calculated by dividing A by E (W= A/E) (Farquhar, 1991).
Temperature data
The maximum and minimum daily temperature (°C) of the air inside the greenhouse was measured with a maximum and mine mercury column thermometer, Taylor brand (England). The maximum and minimum mean temperatures recorded in the greenhouse during the entire experiment were 31.9 and 8.4 °C, respectively.
Moisture content in the soil
The soil water content was determined by the gravimetric method every third day with a Tor-Rey brand electronic scale, PCR Series model, adding the amount of evapotranspirated water in each experimental unit, to keep the soil moisture content close to CC in all the plants, except during the drought treatments (D1 and D2), in which the application of water was suspended and only the weight of each tube was recorded, to estimate the evapotranspiration occurred in the plants subject to water deficit.
Statistical analysis
The data obtained was analyzed with the GLM procedure (SAS Inst. Inc., version 9.12), version for Windows 10, under a split plot design with two main factors; the greatest effect was the soil moisture treatment and the least treatment was the genotype (Hinkelmann and Kempthorne, 2008).
The means of the soil moisture treatments and genotypes were compared using the minimum significant difference (MSD, p< 0.05), under the following statistical model: Yij= µ + Ti + Gj + T*Gjj + Eij. Where: Yij= value of the response variable in soil moisture treatment (T) of level i, genotype (G) of level j; µ= general mean, Ti = effect of soil moisture treatments at level i = irrigation and drought, and Gj = effect of the genotype of the different varieties of lucerne at level j= San Miguel, Oaxaca, Genex, Júpiter, Milenia, San Isidro, Aragón, Cuf 101, Victoria and Atlixco, and Eij= experimental error.
Results and discussion
Soil water content
In irrigation (I1 and I2) the water content of the soil remained close to CC during the experiment, while in drought (D1 and D2), the availability of soil moisture decreased below the PMP (percentage of soil moisture, where the availability of humidity for the plant is reduced to zero) between 361 to 407 ddt in D1 and 634 to 687 ddt in D2, and it remained in this condition for longer in D2 (53 d) than in D1 (46 d).
A severe decrease in soil moisture content below the root absorption limit (PMP), can greatly affect lucerne morphology, physiology, and yield (Pembleton et al., 2009). However, dehydration of the plant can be delayed by a longer absorption period or reduced transpiration; when root growth and absorption stop in a drying soil, lucerne can maintain water deficit by reducing it (Erice et al., 2010) and then stop its vegetative growth (Sheaffer et al., 1988).
Water relations
The drought was more severe and lasted longer in D2 than in D1; ψ, π and p in irrigation (I1 and I2) were consistently higher (p≤ 0.01) than in drought (D2 and D2) on average of all genotypes. In I1 the ψ, π and p remained at an average of -0.6, -1.6 and 1 MPa during the period of water deficit (Figures 1a, b and c), while in D1the ψ, π and p decreased (p≤ 0.01) from - 0.6, -1.6 and 1 MPa, at the beginning of the water deficit period (345 ddt) at -3.1, -3.5 and 0.4 MPa at the end of the drought period (406 ddt) (Figures 1d, e and f).
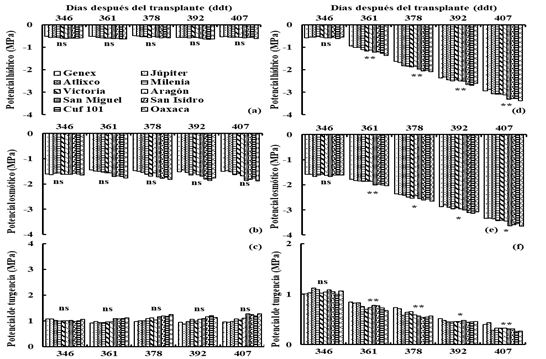
Figure 1 Water, osmotic and turgor potential in irrigation (I1) (a, b and c), and drought (D1) (d, e and f), for 10 varieties of lucerne in greenhouse conditions. Montecillo, Texcoco, State of Mexico.
In D1 the Júpiter variety had high (p≤ 0.05) ψ at 392 and 407 ddt (Figure 1d), and π at 361, 378 and 407 ddt; the Atlixco variety had high (p≤ 0.05) π at 361 and 407 ddt and the Victoria variety showed high (p≤ 0.05) π at 361 ddt (Figure 1e).
The varieties Júpiter, Atlixco and Milenia showed ψ similar to Genex at 361 ddt, Júpiter showed ψ similar to 361 ddt, Genex showed ψ similar to Jupiter at 392 and 407 ddt (Figure 1e).
The Genex, Milenia and Aragón varieties had π similar to Jupiter, Atlixco and Victoria at 361 ddt, the Atlixco and Victoria varieties had π similar to Jupiter at 378 ddt; the Júpiter, Atlixco and Milenia varieties showed π similar to Genex at 392 ddt and the Milenia, Victoria and Aragón varieties were π similar to Genex, Júpiter and Atlixco (Figure 1e). The Milenia variety showed p similar to Genex at 378 ddt (Figure 1f).
In I2 the ψ, π and p were -0.6, -1.6 and 1 MPa on average during the period of soil water deficiencies (Figures 2a, b and c) while in D2, the ψ, π and P decreased (p≤ 0.01) from - 0.6, -1.6 and 1 MPa, at the beginning of the drought (620 ddt) to -3.5, -3.7 and 0.2 MPa at the end of the water deficit period (688 ddt) (Figures 2d, e and f).
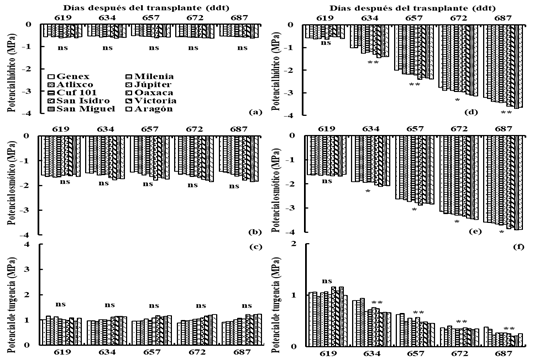
Figure 2 Water, osmotic and turgor potential in irrigation (I2) (a, b and c) and drought (D2) (d, e and f), for 10 varieties of lucerne under greenhouse conditions. Montecillo, Texcoco, State of Mexico.
In D2 the Genex variety showed high (p≤ 0.05) ψ at 634, 672 and 687 ddt (Figure 2d) and π at 657, 672 and 687 ddt (Figure 2e). The Atlixco variety had high (p≤ 0.05) ψ at 634 ddt (Figure 2d), π at 634 ddt (Figure 2e) and p at 634 ddt (Figure 2f). The Milenia variety presented high (p ≤ 0.05) ψ at 634, 657 and 687 ddt (Figure 2d), π at 672 ddt (Figure 2e) and p at 657 ddt (Figure 2f).
The Genex variety showed ψ similar to Milenia at 657 ddt; Atlixco and Milenia varieties had ψ similar to Genex at 672 ddt and Atlixco showed ψ similar to Genex and Milenia at 687 ddt (Figure 2d). The Milenia variety had π similar to Atlixco at 634 ddt; the Atlixco and Milenia varieties had π similar to Genex at 657 ddt, the Atlixco variety had π similar to Genex and Milenia at 672 ddt and the Atlixco and Milenia varieties were similar in π to Genex at 687 ddt (Figure 2e). The Genex and San Isidro varieties showed p similar to Atlixco at 634 ddt (Figure 2f).
A water deficit in the soil (≈ 60% of the extractable moisture) in adult plants of the varieties Cuf 101, Moapa 69 and WL 318 of lucerne, linearly reduced the ψ of all genotypes until the availability of humidity was very low to a depth of 2.4 m (≈ 0.14 m3 m-3), with a differential response between varieties (Grimes et al., 1992). The reduction in ψ of -0.5, -0.5 and -0.35 MPa with respect to the well-irrigated lucerne control, decreased dry matter production by 29, 23 and 20% (Godoy-Ávila et al., 2003). However, it has been determined that resistance to drought in lucerne germplasm is associated with a high capacity of tolerance to wilting of the plant, maintenance of the green leaf area and greater number of stems per plant under stress conditions, in addition to a wide genetic variability in physiological and morphological characters of the plant (Anicchiarico et al., 2013)
Osmotic adjustment
The OA increased (p≤ 0.05) as the drought worsened with values from 0.39 to 1.7 MPa in D1 (361 to 407 ddt) and from 0.46 to 1.96 MPa in D2 (634 to 687 ddt), on average of all genotypes; the longest drought period in D2 was reflected in a greater capacity for osmotic adjustment (Figure 3). The OA is one of the adaptive cellular mechanisms to drought, salinity and freezing in plants, the dehydration of the cell enters a state of osmotic tension due to the increase in the cellular concentration of solutes (Blum, 2015).
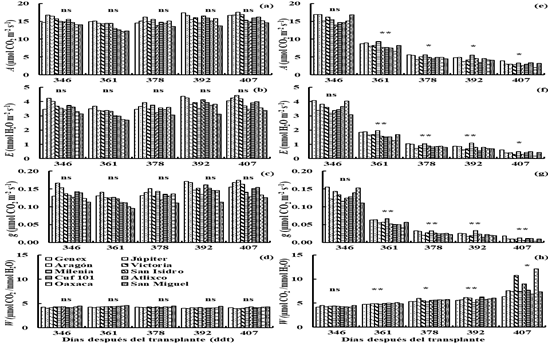
Figure 3 Osmotic adjustment capacity of the plant in D1 (a) and D2 (b) and their respective irrigation periods (I1 and I2), for 10 varieties of lucerne under greenhouse conditions. Montecillo, Texcoco, State of Mexico.
In D1 the Genex, Jupiter and Atlixco varieties had high OA values ( p≤ 0.05) at 361, 378, 392 and 407 ddt, the Victoria variety showed higher OA (p≤ 0.05) at 361 and 378 ddt (p≤ 0.05) and the variety Aragon had a higher OA (p≤ 0.05) at 407 ddt (Figure 3a).
In D2, the Genex, Milenia and Atlixco varieties showed high OA (p≤ 0.05) at 634, 657 and 672 ddt, the Oaxaca variety showed high OA (p≤ 0.05) at 672 and 687 ddt, the Cuf 101 variety showed high OA (p≤ 0.05) at 634 ddt, and the Jupiter variety had high OA (p≤ 0.05) at 657 ddt (Figure 3b). The Genex, Milenia and Atlixco varieties showed an OA similar to Oaxaca at 687 ddt (Figure 3b).
Drought significantly reduced leaf dry weight in three lucerne populations (Chemini, Ganouch, and Tebelbou), with a significant increase in the content of proline, total soluble sugars, amino acids, and potassium in leaves and roots; the Chemini population showed the highest content of solutes, while Tebelbou exhibited the lowest content of proline and the other osmolytes (Abid et al., 2016).
The maintenance of the physiological processes and the hydric status of the leaf suggests that lucerne has some important mechanisms to maintain cellular function when the plant is subject to hydric deficit, in such a way that the maintenance of cellular function under hydric deficit can be related to the processes of antioxidation and OA (He et al., 2012).
An examination of the individual behavior of the lucerne genotypes used in the present study under water deficit conditions, indicated that the Genex, Atlixco and Milenia varieties in D1 and the Genex, Jupiter, Atlixco and Milenia varieties in D2 maintained higher values for the potential ψ, π and p. The high OA capacity observed in the two drought periods (D1 and D2), possibly was one of the mechanisms that contributed to maintaining the survival of the plant, the turgidity of plant organs and tissues, and consequently, the growth and accumulation of dry matter.
Gaseous exchange relationships
The rate of A, E and g in irrigation (I1 and I2) were higher (p≤ 0.05) than in drought (D1 and D2), while W in irrigation (Figure 4) was lower (p≤ 0.05) than in drought (Figure 5) on average of all genotypes. In D1 (345 to 406 ddt) the A, E and g decreased (p≤ 0.05) to 4.4 µmol CO2 m-2 s-1, 0.022 mol H2O m-2 s-1 and 0.7 mmol H2O m-2 s-1 (p≤ 0.05) with respect to the values observed at the beginning of the drought treatment (A=15.3 µmol CO2 m-2 s-1, E= 3.6 mmol H2O m-2 s-1 and g= 0.133 mol H2O m-2 s-1) (Figures 4a, b and c) and the W increased (p≤ 0.05) from 0.0043 to 0.0058 mmol CO2 mmol H2O-1 (Figure 4d).
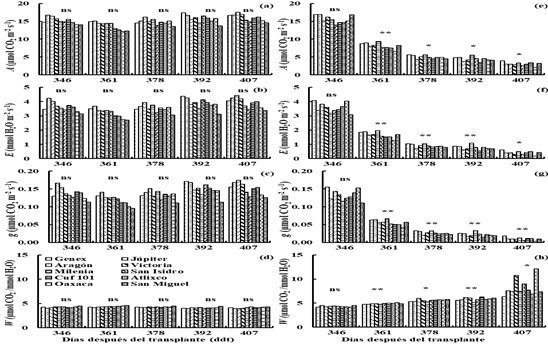
Figure 4 Net assimilation rate (a), transpiration rate (b), stomatal conductance (c) and efficiency in the use of water (d) in irrigation (I1) and drought (D1) (e, f, g and h), for 10 varieties of lucerne in greenhouse conditions. Montecillo, Texcoco, State of Mexico.
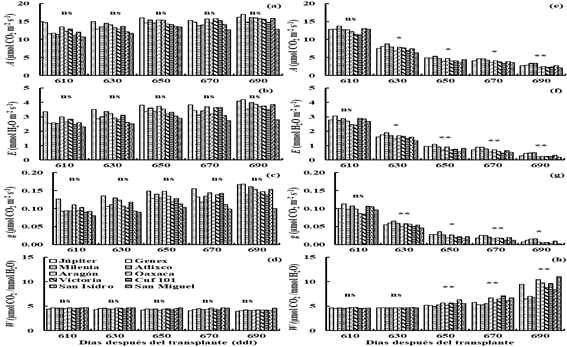
Figure 5 Net assimilation rate (a), transpiration rate (b), stomatal conductance (c) and efficiency in the use of water (d) in irrigation (I2) and drought (D2) (e, f, g and h), for 10 varieties of lucerne in greenhouse conditions. Montecillo, Texcoco, State of Mexico.
In D1 the Milenia variety showed the highest values (p≤ 0.05) of At 361, 392 and 407 ddt (Figure 4a); E at 361, 378, 392 and 407 ddt (Figure 4b) and g at 361, 378, 392 and 407 ddt (Figure 4c). The Genex variety had high values (p≤ 0.05) of A, E and g at 378 ddt (Figure 4a, b and c). The Oaxaca variety showed high value (p≤ 0.05) for W at 361 and 407 ddt, the Aragón variety had high value (p≤ 0.05) for W at 378 ddt and the Cuf 101 variety showed high value (p≤ 0.05) for W at 392 ddt (Figure 4d).
The Jupiter variety had A similar to Milenia at 361 ddt, the Milenia variety had A similar Genex at 378 ddt, the Genex and Jupiter varieties had A similar to Milenia at 392 ddt and 407 (Figure 4a), the Genex and Júpiter were similar to Milenia in E at 361, 392 and 407 ddt and the Júpiter variety was similar to Genex and Milenia in E at 378 ddt (Figure 4b). The Genex and Jupiter varieties were similar to Milenia in g at 361, 392 and 407 ddt, the Jupiter variety was similar in g to Genex and Milenia at 378 ddt (Figure 4c). The Cuf 101 variety was similar to Oaxaca in W at 361 ddt, the San Miguel variety had a W similar to Aragon at 378 ddt, the Aragon variety was similar in W to Cuf 101 at 392 ddt and the Victoria variety had a W similar to Oaxaca at 407 ddt (Figure 4d).
In D2 (620-688 ddt), the rate of A, E and g decreased (p≤ 0.05) to 4 µmol CO2 m-2 s-1, 0.019 mol H2O m-2 s-1 and 0.7 mmol H2O m-2 s-1 with respect to the values observed at the beginning of the drought treatment (A= 12.5 µmol CO2 m-2 s-1, E= 2.8 mmol H2O m-2 s-1 and g= 0.100 mol H2O m-2 s-1) (Figures 5a, b and c), and the W increased from 0.0046 to 0.0061 mmol CO2 mmol-1 H2O-1 (Figure 5d).
The Milenia variety showed the highest values (p≤ 0.05) for A, E and g at 634, 657, 672 and 687 ddt; the Genex variety had high values (p≤ 0.05) for A, E and g at 672 and 687 ddt (Figures 5a, b and c). The San Isidro variety showed the highest value (p≤ 0.05) for W at 657 ddt, the Cuf 101 variety had a high value (p≤ 0.05) of W at 672 ddt and the San Miguel variety showed high value (p≤ 0.05) for W at 687 ddt (Figure 5d).
The Genex variety showed values of A, E and g similar to Milenia at 634 (Figures 5a, b and c), Jupiter had A similar to Milenia at 657 ddt, the varieties Jupiter, Atlixco and Oaxaca showed A similar to Genex and Milenia at 672 ddt and the varieties Júpiter and Oaxaca had A similar to Genex and Milenia at 687 ddt (Figure 5a). The Júpiter, Genex, Atlixco and Oaxaca varieties had an E similar to Milenia at 657 ddt (Figure 5b). The varieties Jupiter and Genex had Milenia had a g similar at 657 ddt (Figure 5c) and the Atlixco variety showed E and g similar to Genex and Milenia at 672 and 687 ddt (Figures 5b and c). The Victoria variety had a W similar to Cuf 101 at 672 ddt and San Miguel at 687 ddt (Figure 5d).
In another study, it was observed that the soil water deficit severely reduced the net assimilation rate, stomatal conductance, water use efficiency, relative water content, leaf area and chlorophyll content a and b in various lucerne populations, with an increase in the efficiency in the use of water and internal concentration of CO2; additionally, it was determined that the most drought tolerant population reduced their leaf area as much as possible and maintained the highest values for water use efficiency (Abid et al., 2016a).
Lucerne is generally considered a highly drought-sensitive species, especially in the vegetative and reproductive stages, and cultivated in dry areas is moderately drought-tolerant, but under conditions of severe water stress suffers considerable production losses. The main effects of water stress are manifested in a reduction in germination and growth, severe damage to the photosynthetic apparatus, and a decrease in net photosynthesis and nutrient absorption (Nadeem et al., 2019).
Conclusions
The soil water deficit reduced the plant’s water and osmotic potentials to levels that favored a high active accumulation of solutes or osmotic adjustment, which allowed plants to maintain favorable turgidity levels for gas exchange (CO2 molecules/molecules of H2O) of the leaf area.
The Genex, Júpiter, Atlixco and Milenia varieties with high levels of osmotic adjustment, photosynthetic rate, transpiration rate, stomatal conductance and efficiency in the use of water showed better behavior and tolerance to water stress.
A high osmotic adjustment capacity, gas exchange and efficiency in the use of water are genetic-physiological characteristics of the plant, which could be useful in the selection of new lucerne genotypes with drought tolerance.
Literatura citada
Abid, M.; Mansour, E.; Khaled, A. B.; Bachar, Kh. D.; Yahya, B. L. and Ferchichi, A. 2016. Induced osmotic adjustment in alfalfa plants confers tolerance to water stress. Bulg. J. Agric. Sci. 22(3):415-425. [ Links ]
Abid, M.; Haddad, M.; Khaled, A. B.; Mansour, E.; Bachar, K.; Lacheheb, B.; Ferchichi, A. 2016a. Water relations and gas exchange in alfalfa leaves under drought conditions in southern Tunisian oases. Pol. J. Environ. Stud. 25(3):917-924. [ Links ]
Annicchiarico, P.; Pecetti, L. and Tava, A. 2013. Physiological and morphological traits associated with adaptation of lucerne (Medicago sativa) to severely drought-stressed and to irrigated environments. Ann. Appl. Biol. 162(1):27-40. Doi:10.1111/j.1744-7348.2012.00576.x. [ Links ]
Barrios-Gómez, E. J.; López-Castañeda, C. y Kohashi-Shibata, J. 2011. Relaciones hídricas y temperaturas altas en frijol del tipo ‘Flor de Mayo’. Agron. Costarr. 35(1):131-145. [ Links ]
Begg, J. E. and Turner, N. C. 1976. Crop water deficits. Adv. Agron. 28(C):161-217. [ Links ]
Blum, A. 2015. Stress, strain, signaling, and adaptation-not just a matter of definition. J. Exp. Bot. 67(3):562-565. [ Links ]
Fahad, S.; Hussain, S.; Matloob, A.; Khan, F. A.; Khaliq, A. and Saud, S. 2015. Phytohormones and plant responses to salinity stress: a review. Plant Growth Reg. 75(2):391-404. Doi: 10.1007/s10725-014-0013-y. [ Links ]
Farquhar, G. D. 1991. Use of stable isotopes in evaluating plant water use efficiency. In: Proceedings series of the International symposium on the use of stable isotopes in plant nutrition, soil fertility and environmental studies, organized by the International Atomic Energy Agency, Vienna (Austria) and the Food and Agriculture Organization of the United Nations, Rome (Italy). Vienna, Austria, 1-5 October, 1990. 475-488 pp. [ Links ]
Godoy-Ávila, C.; Pérez-Gutiérrez, A.; Torres-E, C. A.; Hermosillo, L. J. y Reyes-J, I. 2003. Uso de agua, producción de forraje y relaciones hídricas en alfalfa con riego por goteo subsuperficial. Agrociencia. 37(2):107-115. [ Links ]
Grimes, D. W.; Wiley, P. L. and Sheesley, W. R. 1992. Alfalfa yield and plant water relations with variable irrigation. Crop Sci. 32(6):1381-1387. [ Links ]
Harrison, M. T.; Tardieu, F.; Dong, Z.; Messina, C. D. and Hammer, G. L. 2014. Characterizing drought stress and trait influence on maize yield under current and future conditions. Glob. Change Biol. 20(3):867-878. Doi:10.1111/gcb.12381. [ Links ]
He, S., Liu, G. and Yang, H. 2012. Water use efficiency by alfalfa: Mechanisms involving anti-oxidation and osmotic adjustment under drought. Russ. J. Plant Physiol. 59(3):348-355. [ Links ]
Hinkelmann, K. and Kempthorne, O. 2008. Design and analysis of experiments. Volume 1: Introduction to experimental design. A John Wiley and Sons, Inc., United States of America. 631 p. [ Links ]
López-Castañeda, C. 1993. Eficiencia en el uso del agua y discriminación de carbono en cereales. Agrociencia. 4(4):7-32. [ Links ]
Morgan, J. M. 1995. Growth and yield of wheat lines with differing osmoregulative capacity at high soil water deficit in seasons of varying evaporative demand. Field Crops Res. 40(3):143-152. [ Links ]
Morran, S.; Eini, O.; Pyvovarenko, T.; Parent, B.; Singh, R. and Ismagul, A. 2011. Improvement of stress tolerance of wheat and barley by modulation of expression of DREB/CBF factors. Plant Biotech. J. 9(2):230-249. Doi: 10.1111/j.1467-7652.2010.00547.x. [ Links ]
Nadeem, M.; Li., J.; Yahya, M.; Sher, A.; Ma, C.; Wang, X. and Qiu, L. 2019. Research progress and perspective on drought stress in legumes: a review. Inter. J. Mol. Sci. 20(1):1-32. [ Links ]
Orloff, S. 2015. Drought strategies for alfalfa. University of California, Agriculture and Natural Resources, Publication 8522, July 2015. http://anrcatalog.ucanr.edu. [ Links ]
Pembleton, K. G.; Rawnsley, R. P.; Donaghy, D. J. and Volenec, J. J. 2009. Water deficit alters canopy structure but not photosynthesis during the regrowth of alfalfa. Crop Sci. 49(2):722-731. [ Links ]
Perdiguero, P.; Soto, Á. and Collada, C. 2015. Comparative analysis of Pinus pinea and Pinus pinaster dehydrins under drought stress. Tree Genetics & Genomes 11:70. Doi: 10.1007/s11295-015-0899-1. [ Links ]
Sheaffer, C. C.; Tanner, C. B. and Kirkham, M. B. 1988. Alfalfa water relations and irrigation. In: Hanson, A. A.; Barnes, D. K. and Hill, Jr. R. R. (Eds.). Alfalfa and alfalfa improvement. Madison, WI, USA: ASA-CSSA-SSSA. 373-409 pp. [ Links ]
Sim, R. E.; Brown, H. E.; Teixeira, E. I. and Moot, D. J. 2017. Soil water extraction patterns of lucerne grown on stony soils. Plant Soil. 414:95-112. Doi 10.1007/s11104-016-3112-x. [ Links ]
Zhang, C.; Shi, S.; Liu, Z.; Yang, F. and Yin, G. 2019. Drought tolerance in alfalfa (Medicago sativa L.) varieties is associated with enhanced antioxidative protection and declined lipid peroxidation. J. Plant Physiol. 232(1):226-240. [ Links ]
Zhao, P., Liu, P., Yuan, G., Jia, J., Li, X., Qi, D., Chen, S., Ma, T., Liu, G. and Cheng, L. 2016. New insights on drought stress response by global investigation of gene expression changes in Sheepgrass (Leymus chinensis). Front. Plant Sci. 7:954. Doi: 10.3389/fpls.2016.00954. [ Links ]
Received: January 01, 2020; Accepted: March 01, 2020
 Este es un artículo publicado en acceso abierto bajo una licencia
Creative Commons
Este es un artículo publicado en acceso abierto bajo una licencia
Creative Commons
