











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
Los cítricos (Citrus spp.) son considerados como la fruta universal ya que están presentes en más de 100 países y son el grupo de frutas más importante económicamente, abarcando 20% del mercado mundial de frutas. Actualmente, la mayor parte de la cosecha de cítricos proviene de 10 países que aportan 77% de la producción, de los cuales China y Brasil producen 42% y México ocupa la cuarta posición después de los EE. UU. (Lerma et al., 2015).
Las enfermedades constituyen un factor limitante en la producción de estos cultivos en las zonas productoras de México, causadas por alrededor de 50 patógenos que provocan diversos síntomas tales como gomosis, agrietamiento, caída de frutos, mancha grasienta, roña, muerte de ramas, entre otros (Orozco et al., 2013). El hongo Lasiodiplodia spp., ha sido reportado como causante de muerte regresiva y otros síntomas en cítricos y en una amplia gama de cultivos en diversos países (Adesemoye et al., 2014; Rodríguez et al., 2016).
Recientemente Polanco et al. (2019) reportaron a Lasiodiplodia theobromae y Fomitopsis meliae como los hongos causantes de la muerte regresiva de los cítricos en árboles de huertos de los estados de Nuevo León y Tamaulipas, México. Por lo general, el método de control de este hongo es mediante aplicaciones de fungicidas sintéticos. Los fungicidas químicos pueden producir efectos colaterales como la resistencia en plagas y enfermedades, la reducción de las poblaciones de microorganismos benéficos y la contaminación del ambiente (Adeniyi et al., 2013; Tovar et al., 2013; Álvarez, 2015).
Los agentes utilizados para biocontrol reducen significativamente las poblaciones de fitopatógenos y con su uso se pretende equilibrar las perturbaciones ecológicas provocadas por los agroquímicos (Badii y Abreu, 2006) mediante diversos mecanismos como la producción de toxinas (Carisse et al., 2001), la competencia por nutrientes y espacio (Mutawila et al., 2011), inhibición del crecimiento micelial del patógeno y micoparasitismo (Cheng et al., 2012).
Entre los grupos de microorganismos utilizados para biocontrol se encuentran los actinomicetos, los cuales han sido ampliamente estudiados por su capacidad de producir compuestos antibióticos y antifúngicos (Lee et al., 2002), especialmente contra fitopatógenos de importancia agrícola (Sreevidya y Gopalakrishnan 2012; Abdallah et al., 2013; Dávila et al., 2013). Los actinomicetos son bacterias Gram positivas, halladas generalmente en suelo, ríos, lagos, material orgánico en descomposición, entre otros lugares. Muchos son saprofitos y algunas especies son patógenas (Quiñones et al., 2016), como Streptomyces scabiei causante de la sarna común del cultivo de papa.
Desde hace varias décadas, el número de actinomicetos utilizados en la agricultura ha estado en constante aumento como un medio para controlar o reducir las poblaciones de fitopatógenos (Dávila et al., 2013). Aunque los microorganismos han sido estudiados ampliamente en el control de hongos fitopatógenos, son pocos los trabajos realizados en el control de los hongos Lasiodiplodia theobromae y Fomitopsis meliae. Sajitha y Florence (2013); Kamil et al. (2018), evaluaron la capacidad antagónica de actinomicetos para el control de Lasiodiplodia theobromae causantes de daño a la madera del árbol de caucho y la muerte regresiva de plantas de mango respectivamente, con buenos resultados; además Caro et al. (2019) trabajando con actinomicetos obtuvieron resultados promisorios para el control de Lasiodiplodia sp., en el cultivo de papa.
Aun así, es poco lo que se conoce de trabajos de control de los principales hongos causantes de la muerte regresiva de los críticos, Lasiodiplodia theobromae y Fomitopsis meliae, a los cuales se trata normalmente con el uso de productos químicos o practicas sanitarias del cultivo (podas). Por lo anterior, el objetivo de este trabajo fue aislar cepas de microorganismos de suelo y evaluar su capacidad antagónica in vitro frente a los fitopatógenos Lasiodiplodia theobromae y Fomitopsis meliae, causantes de la muerte regresiva de los cítricos.
Materiales y métodos
Cepas fúngicas fitopatógenas
Las cepas fúngicas Lasiodiplodia theobromae y Fomitopsis meliae fueron proporcionadas por la Facultad de Agronomía de la Universidad Autónoma de Nuevo León (FAUANL).
Aislamiento de microorganismos del suelo
Se realizaron 17 muestreos de suelo con diferentes tipos de vegetación en zonas del estado de Nuevo León (Cuadro 1), se colectaron muestras de la rizósfera a una profundidad de 10 a 15 cm, después de eliminar la capa superficial. Para el muestreo se seleccionaron al azar cinco puntos diferentes para formar una muestra. Las muestras se colocaron en bolsas de plástico, se sellaron y almacenaron en el laboratorio en condiciones de clima fresco.
Cuadro 1 Relación de lugares de muestreo y tipo de vegetación predominante.
| Localización | Coordenadas | Tipo de vegetación |
| FAUANL, Campus Marín, municipio Marín | 25° 52’ 25.76” latitud norte; 100° 3’ 21.25” latitud oeste | Cultivo de cítricos y terreno baldío |
| FAUANL, Campus La Ascensión, municipio Aramberri | 24.32° latitud norte; 99.91° latitud oeste | Cultivo de ajo |
| Municipio de Mina | 25° 52’ a 26° 44’ latitud norte; 100° 26’ a 101° 13’ latitud oeste | Plantas silvestres |
Un gramo de cada muestra se disolvió en 9 mL de agua destilada estéril. Las diluciones fueron tratadas con calor (50 °C) durante 10 min. De cada suspensión se prepararon diluciones seriales 1:100, 1:1 000 y 1:10 000. De cada dilución se transfirieron 100 µL a cajas Petri con agar papa dextrosa adicionado con extracto de levadura (0.3%) y extracto de malta (0.3%) (PDLMA) mediante el método de extensión. Las cajas Petri se incubaron a 25 ±2 ºC. Se aislaron y purificaron microorganismos que presentaron halos de inhibición (Oskay et al., 2004).
Caracterización morfológica y genética de actinomicetos y bacterias con actividad antagónica
Las cepas de actinomicetos se identificaron de acuerdo con sus características de crecimiento macroscópicas (forma, color del micelio aéreo, textura y la producción de pigmentos en el medio de cultivo). Adicionalmente, se realizaron observaciones microscópicas utilizando para ello la tinción de Gram. Para la identificación de las bacterias se consideraron forma, color y textura de las colonias, para la visualización microscópica se utilizó la tinción de Gram.
Para la identificación molecular de los actinomicetos se extrajo el ADN utilizando el DNeasy Plant Mini KitMR (Qiagen Inc.) y se amplificó por PCR utilizando los iniciadores 8F (5’-agagtttgatcctggctcag-3’) y 1492R (5’-ggttaccttgttacgactt-3’) (Sacchi et al., 2002). Las reacciones de amplificación se llevaron a cabo en un termociclador ThermoMR, las cuales consistieron en una desnaturalización a 94 ºC por 1 min, seguido por 35 ciclos a 94 ºC por 30 s, un anillamiento a 48 ºC por 50 s y una extensión a 72 ºC por 80 s y una extensión final a 72 ºC por 4 min. Los productos se visualizaron por electroforesis en gel de agarosa al 1% teñido con 0.5 ng µl-1 de bromuro de etidio. Se utilizó el marcador de peso molecular ladder-100MR (Axygen).
Caracterización morfológica y genética de los hongos con actividad antagónica
La identificación morfológica de los aislados obtenidos estuvo basada en las características de las colonias (color, tipo y forma) además de la morfología de conidios y conidióforos utilizando un microscopio óptico (Úrbaez et al., 2013). Además, se realizaron montajes en laminillas utilizando el colorante lactofenol azul de algodón como solución, y se describieron las características particulares de las estructuras. Los hongos se identificaron siguiendo los criterios de Barnett y Hunter (1998); Phillips et al. (2013).
Para la caracterización genética, se extrajo el ADN genómico a partir de micelio siguiendo las instrucciones del fabricante del kit DNeasy Plant Mini KitMR (Qiagen Inc.). El ADN se cuantificó con un espectrofotómetro Take3MR (Bioselec). Se amplificó por PCR la región ITS-1, el gen ribosomal 5.8S y el ITS-2 utilizando los iniciadores ITS-1fu 5`-tccgtaggtgaacctgcgg-3’ e ITS-4 5’-tcctccgcttatttgatatgc-3’ (White et al., 1990). Las reacciones de amplificación se llevaron a cabo en un termociclador TermoMR, las cuales consistieron en una desnaturalización a 94 ºC por 1 min, un anillamiento a 55 ºC por 25 s, una polimerización a 72 ºC durante 50 s, con 35 ciclos, seguido de una extensión final a 72 ºC por 4 min. Los productos de PCR fueron secuenciados y comparados con secuencias del GenBank para confirmar la identidad de las cepas a nivel de especie.
Prueba de antagonismo in vitro
Se realizó una prueba preliminar para seleccionar las cepas que presentaran mayor efecto inhibitorio sobre los hongos patógenos. Las confrontaciones se realizaron con cada cepa de los microorganismos aislados contra los hongos fitopatógenos L. theobromae y F. meliae. Se sembró una azada de cada uno de los microorganismos en los cuatro puntos cardinales (3 cm de distancia del centro, en cajas Petri) en el medio PDLMA. Cuarenta y ocho horas después un explante de 0.5 cm de crecimiento de cada hongo fitopatógeno fue colocado en el centro. Las cajas fueron incubadas a 25 ±2 ºC durante 5 días. Cada microorganismo se tomó como un tratamiento con tres repeticiones, considerando cada caja Petri como una repetición y un control negativo con cada hongo sin antagonista. El ensayo se realizó dos veces.
La capacidad inhibitoria de los microorganismos se cuantificó midiendo el diámetro del crecimiento fúngico con un vernier, usando dos diámetros cardinales previamente dibujados en la parte baja de cada caja Petri como referencia.
El porcentaje de inhibición fue calculado usando la siguiente fórmula: (%) de inhibición = [(A1-A2)/A1] x 100. Donde: A1= crecimiento de la colonia del fitopatógeno (testigo); A2 = crecimiento de la colonia fungosa del fitopatógeno creciendo frente al actinomiceto.
Con base a los resultados de las pruebas preliminares de las confrontaciones in vitro, se seleccionaron las cepas M1-101 y M104 para un segundo ensayo por presentar los valores más altos de inhibición. Las cepas antagonistas fueron cultivadas en caldo papa dextrosa adicionado con extracto de levadura y extracto de malta al 0.3% (CPDLM), e incubadas a 25 ±2 ºC y 125 rpm durante 10 días. Posteriormente, el contenido fue centrifugado, filtrado y colocado en frascos estériles.
La actividad de inhibición de cada antagonista fue evaluada a concentraciones de 25, 50 y 100% del filtrado. La técnica consistió en esparcir por extensión 100 µl del antagonista. Posteriormente, fue colocado un disco de 5 mm del crecimiento micelial del fitopatógeno en el centro de la caja Petri. Además, se confrontó cada antagonista con los dos hongos fitopatógenos al mismo tiempo, para lo cual se esparcieron 100 µl del antagonista por el método de extensión en cajas e inmediatamente un disco del crecimiento micelial de cada hongo de 5 mm de diámetro fue colocado a una distancia de 4 cm entre ellos. Se realizaron observaciones diarias durante cinco días, al final de los cuales se midió el crecimiento micelial de los hongos, cuando el crecimiento del control cubrió por completo la caja Petri.
Inhibición de la germinación de conidios y micelio
Con base en los resultados preliminares se seleccionó la cepa M1-101 para la prueba de inhibición de la germinación de conidios y micelio de los hongos F. meliae y L. theobromae, respectivamente. La cepa M1-101 se sembró en 50 ml de caldo CPDLM mediante la adición de un cuadro de la cepa en agar de aproximadamente 1 cm2 por cada matraz Erlenmeyer bafleado de 250 ml. Los matraces se incubaron en agitación a 125 rpm y 25 ±2 ºC durante 10 días. Al término del periodo de incubación, se filtró al cultivo y el filtrado se diluyó hasta tener concentraciones de 25, 50 y 100%.
Para la preparación del inóculo de los hongos fitopatógenos se usaron cepas incubadas por 6 días en papa dextrosa agar (PDA). Se agregaron 10 ml de agua destilada estéril a cada caja Petri para resuspender los conidios de F. meliae y el micelio de L. theobromae raspando la superficie de la caja con un asa bacteriológica. El inoculo se conservó en tubos cónicos de 15 ml.
La actividad de inhibición de la germinación del antagonista fue evaluada a concentraciones de 25, 50 y 100%. La técnica consistió en extender con ayuda de un asa de vidrio, 100 µl de cada concentración en cada caja Petri. Después de absorción, se añadieron 100 µl del inoculo de los fitopatógenos de la solución previamente preparada (conidios y micelio) de los fitopatógenos. Se evaluaron tres réplicas de cada concentración y las cajas Petri se incubaron a 25 ±2 ºC. Cada 24 h se realizaron observaciones para ver la inhibición de la germinación de conidios y los posibles cambios en la morfología del micelio.
El recuento de conidios germinados y no germinados fue realizado usando un microscopio compuesto (Leica DM500). Se contabilizaron 100 conidios cm-2, 500 conidios en total (germinados y no germinados). Para determinar el porcentaje de conidios germinados se utilizó la siguiente fórmula: % germinación = (a/a+b) x 100. Donde: a = número de conidios germinadas; y b= número de conidios sin germinar. Con L. theobromae, sólo se observaron los cambios en la morfología del micelio inducido por la presencia de los compuestos antifúngicos, producidos por el antagonista a un nivel microscópico.
Resultados
Se aislaron 70 cepas de microorganismos de las cuales 15 presentaron actividad inhibitoria, y de estas, ocho presentaban características típicas de actinobacterias, cinco bacterias bacilares y dos hongos (Cuadro 2).
Cuadro 2 Microorganismos aislados de suelo de diferentes zonas del estado de Nuevo León con actividad antagónica.
| Cepas | Lugar de muestreo | Clasificación |
| A7 | Mina | Bacilo |
| A14 | Mina | Bacilo |
| A18 | Mina | Bacilo |
| A20 | Mina | Bacilo |
| A23 | Mina | Bacilo |
| CH 102 | La Ascensión | Actinomiceto |
| CH 103 | La Ascensión | Actinomiceto |
| M 104 | Marín | Hongo |
| M1-101 | Marín | Actinomiceto |
| M2 R1 | Mina | Actinomiceto |
| M2 R2 | Mina | Actinomiceto |
| M3 | Marín | Actinomiceto |
| M4 | Marín | Actinomiceto |
| M4 R | Mina | Actinomiceto |
| M5-102 | Marín | Hongo |
Caracterización morfológica y genética de los actinomicetos y bacterias bacilares
Las características de crecimiento colonial de los actinomicetos en cultivo fueron masas aéreas secas, polvosas, con bordes irregulares, de pigmentaciones variadas, observándose tonalidades de color amarillo, gris y otras de color anaranjado. Al microscopio (40X) mostraron las ramificaciones típicas de los actinomicetos. Por tinción Gram se observaron estreptobacilos esporulados Gram positivos.
Las bacterias mostraron tonalidades variables como amarillo y crema (Figura 1), con textura acuosa y bordes regulares. Mediante tinción Gram se observaron bacilos Gram positivos y Gram negativos. No se logró la identificación molecular de las cepas de actinomicetos por una falta de amplificación por PCR atribuida posiblemente a que los iniciadores o primers han sido más utilizados con otras bacterias y tal vez no tengan la especificidad suficiente para unirse a las secuencias de actinomicetos.
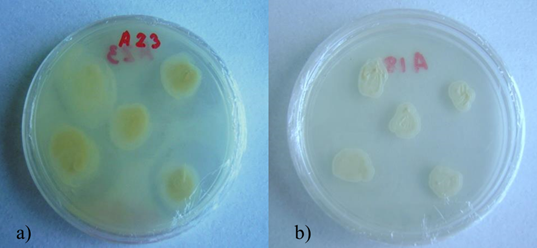
Las bacterias bacilares obtenidas fueron amplificadas por PCR y se secuenció parte del gen 16S. Las secuencias consenso de las bacterias bacilares fueron comparadas con secuencias depositadas en el GenBank a través del programa Blast; la cepa A18 correspondió a Achromobacter xylosoxidans con un 96% de homología y A23 mostró 96% de similitud con Delftia sp.
Caracterización morfológica y genética de los hongos con actividad antifúngica
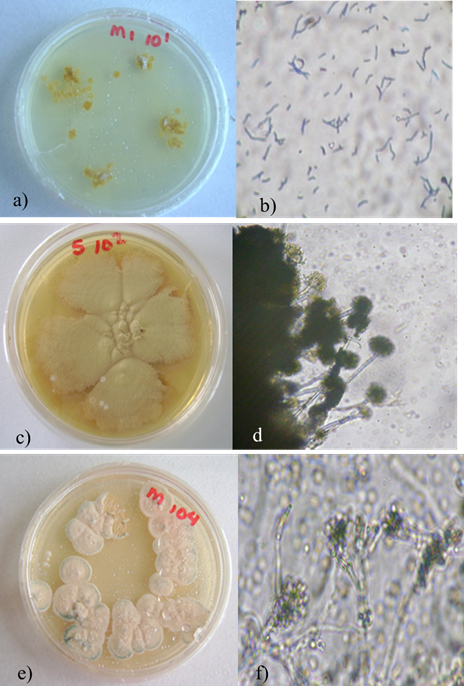
Las colonias de la cepa M104 en medio de cultivo fueron de color blanco, compactas y con bordes irregulares, y el micelio de la cepa M5-102 de color café (Figura 2). La cepa M104 fue identificada como Penicillium sp. Las características del conidióforo mostraban forma de escobilla y conidios hialinos, unicelulares en forma cilíndrica. La cepa M5-102 presentó conidióforos y cabezas aspergilar, conidios hialinos, unicelulares y cilíndricos; esta cepa fue identificada como Aspergillus sp. La identificación morfológica fue confirmada por su secuencia de ADN, correspondiendo a Penicillium citrinum y Aspergillus sp.
Prueba de antagonismo in vitro
Acromobacter xylosoxidans redujo el crecimiento micelial de L. theobromae 63% y a F. meliae 59%. Por otro lado, Delftia sp. inhibió en un 39% a ambos hongos fitopatógenos. Aspergillus sp., redujo 82% el crecimiento de L. theobromae y a F. meliae en 83%. La cepa identificada como P. citrinum fue capaz de reducir el crecimiento micelial de L. theobromae en 83% manteniéndolo inhibido durante más 20 días. Los valores más altos de inhibición se obtuvieron con el hongo P. citrinum (M104) y acinomicetos M4R, y M1-101, los cuales mostraron porcentajes de inhibición de 85, 84 y 91% contra L. theobromae y 91, 83, y 91%, contra F. meliae respectivamente.
En la prueba de inhibición con las diferentes concentraciones de los filtrados del actinomiceto y P. citrinum (25, 50 y 100%), hubo diferencias estadísticamente significativas (p< 0.001) en el porcentaje de inhibición de los patógenos. El control presentó el crecimiento micelial más alto a los 5 días, después de que los hongos se colocaron en PDA. Los hongos L. theobromae y F. meliae tuvieron un crecimiento similar, no mostrando diferencias significativas entre ellos. Los tratamientos que mostraron mayor inhibición de los patógenos fueron L. theobromae y F. meliae frente a P. citrinum al 100%, seguido por L. theobromae + P. citrinum y F. meliae + P. citrinum con la concentración al 100%. No hubo diferencias entre las tres concentraciones de la cepa M101 contra F. meliae. En la prueba de inhibición con las diferentes concentraciones de los filtrados del actinomiceto y P. citrinum (25, 50 y 100%), hubo diferencias estadísticamente significativas (p= 0) en el porcentaje de inhibición de los patógenos.
Inhibición de la germinación de conidios
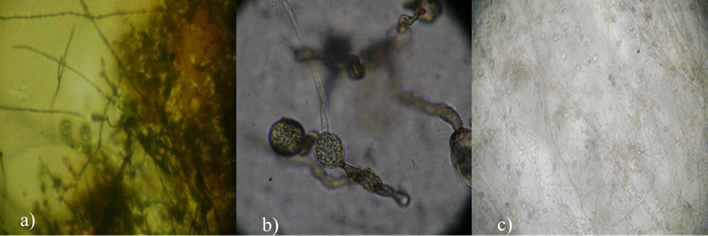
L. theobromae a las 96 h de incubación cubrió por completo la caja Petri sin obtenerse ninguna estructura reproductiva. Sin embargo, en presencia del antagonista, L. theobromae produjo clamidosporas de color café oscuro y células conidiógenas (Figura 3). Con la confrontación, los conidios de F. meliae son inhibidos y no fueron capaces de germinar, se observó que los conidios se agrupan y engrosan tendiendo a formar clamidosporas (Figura 3).
Discusión
En este estudio la bacteria A. xylosoxidans fue efectiva en reducir el crecimiento in vitro de L. theobromae y F. melie. La capacidad que presentó esta bacteria de biocontrolar estos patógenos es comparable con los resultados reportados por Moretti et al. (2008), quienes usaron esta bacteria para controlar a Fusarium oxysporum causante del marchitamiento del tomate. Bagheri y Ahmadzadeh (2016) también reportaron que A. xylosoxidans puede inhibir a Curvularia lunata y Bipolaris sorikinian presentes en el cultivo de trigo.
Por otro lado, algunas especies del género Delftia han sido reportadas por Han et al. (2005) inhibiendo a Xanthomonas oryzae pv. oryzae, Rhizoctonia solani y Pyricularia oryzae. Adicionalmente, estas bacterias desempeñan un papel importante en tecnologías de biorremediación (Ubalde et al., 2012). Jergensen et al. (2009) citan a Delftia con capacidad de romper los enlaces glicosídicos en la quitina y los péptidoglucano, con lo cual pudieran mostrar mayor capacidad de inhibición de los hongos fitopatógenos como los usados en este estudio.
Por lo general, las especies del género Aspergillus están consideradas como hongos saprófitos u oportunistas (Cortés y Mosqueda, 2013). Algunas razas de Aspergillus sp., son consideradas como atoxigénicas debido a que no producen aflatoxinas y han sido registradas como cepas biocontroladoras, cuyo mecanismo de acción es la exclusión competitiva (Cotty, 1994). Debido a esas cualidades, una cepa de Aspergillus fue formulada y ha sido comercializada como agente de biocontrol de la especie Aspergillus flavus productora de aflatoxinas (Dorner, 2009).
En este estudio, Aspergillus sp. fue capaz de inhibir el crecimiento de L. theobromae y F. meliae. Estos resultados son confirmados por los hallazgos obtenidos por Adeniyi et al. (2013) quienes reportaron a A. niger como agente biocontrolador de L. theobromae, aunque sugieren que se debe explotar su potencial en otros fitopatógenos y manejar los posibles efectos adversos en los cultivos.
A pesar de que el hongo P. citrinum puede causar enfermedades en cultivos en campo y productos postcosecha (Marquardt, 1996), de acuerdo con nuestros resultados, algunas especies de Penicillium también poseen potencial de inhibir patógenos. Estos resultados pueden ser explicados de acuerdo con Quiroz et al. (2008), quienes reportaron que diferentes especies de hongos poseen mecanismos tanto de antagonistas como patógenos.
De las 15 cepas que se aislaron y evaluaron, los actinomicetos mostraron los valores más altos de inhibición (M4 R y M1-101), siendo M1-101 la cepa con mayor actividad antagónica, con porcentajes de inhibición contra L. theobromae y F. meliae de 84 y 91%, respectivamente. Sánchez et al. (2011); Wang et al. (2015) aislaron actinomicetos con capacidad de inhibir a fitopatógenos de importancia agrícola como Fusarium equiseti.
Los resultados obtenidos en este estudio demuestran que la habilidad de los microorganismos para inhibir patógenos puede ser incrementada o disminuida, dependiendo de las condiciones de las estructuras de sobrevivencia que puedan producir los últimos y que el antagonista logra reducir su crecimiento por la producción de compuestos antifúngicos o exclusión competitiva.
Conclusiones
Se logró el aislamiento de actinomicetos a partir de muestras de suelo, capaces de inhibir el crecimiento de los patógenos causantes de la muerte regresiva de los cítricos L. theobromae y F. meliae. Las cepas M4R y M104 presentaron porcentajes de inhibición superiores a 84% contra los fitopatógenos anteriores.
Los hongos Aspergillus sp. y P. citrinum fueron capaces de reducir el crecimiento de L. theobromae y F. meliae. Por la capacidad antagonista demostrada en el presente estudio, las cepas de actinomicetos M4R y M104 y los hongos Aspergillus sp. y P. citrinum podrían ser considerados como potenciales candidatos a ser utilizados en programas de control de L. theobromae y F. meliae, pero se necesita continuar con los estudios de esta capacidad antagónica in vivo.

