










Serviços Personalizados
Journal
Artigo
 texto em
texto em  Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista mexicana de ciencias agrícolas
versão impressa ISSN 2007-0934
Rev. Mex. Cienc. Agríc vol.11 no.2 Texcoco Fev./Mar. 2020 Epub 15-Mar-2021
https://doi.org/10.29312/remexca.v11i2.1917
Articles
Diurnal physiological response of tomato cultivation to the application of silicon under salinity conditions
1Faculty of Agronomy-Autonomous University of Sinaloa. Culiacán-El Dorado highway km 17.5, Culiacán, Sinaloa, Mexico. CP. 80000. (lopezcuen13@hotmail.com).
2Institute of Agricultural Sciences-Autonomous University of Baja California. Highway to Delta s/n, Ejido Nuevo León, Baja California, Mexico. CP. 21705. (daniasaf@gmail.com; onecimo.grimaldo@uabc.edu.mx; isabel.escobosa@uabc.edu.mx; victorcardenas@uabc.edu.mx; roberto-soto@uabc.edu.mx; mariacristina@uabc.edu.mx).
Silicon (Si) has beneficial effects on the tolerance of plants to various biotic and abiotic stresses. The objective of this study was to determine the effect of Si nutrition in tomato grown under conditions of induced saline stress (S), on leaf temperature, chlorophyll fluorescence and stomatal conductance (gs). The experiment was conducted under greenhouse conditions during the spring-summer 2017 cycle. A randomized distributed plot design was used. In which the main plot was salinity [with NaCl or with standard nutrient solution (SNS)] and the subplot was the application of Si (0, 4, 8 g L-1). The variables were measured during the day. The results showed that the leaf temperature increased at dawn in plants with NaCl and at sunset the effect was reversed and the highest values were in plants with SNS. At noon, Si × S interaction was identified in the photosynthetic efficiency (Fv/Fm) and minimal fluorescence (Fo) variables. The maximum fluorescence (Fm) was more affected by the type of salinity, with the highest values of plants with SNS. It was concluded that the diurnal physiological response of the tomato plant grown under saline conditions, will depend on the type of salt used and the dose of Si, especially at early and late hours of the day.
Keywords: greenhouse; mineral nutrition; plant physiology; saline stress; sodium chloride
El silicio (Si) tiene efectos benéficos sobre la tolerancia de las plantas a diversos estreses bióticos y abióticos. El objetivo de este estudio fue determinar el efecto de la nutrición con Si en tomate cultivado bajo condiciones de estrés salino inducido (S), sobre la temperatura de las hojas, la fluorescencia de la clorofila y la conductancia estomática (gs). El experimento se realizó bajo condiciones de invernadero durante el ciclo primavera-verano de 2017. Se utilizó un diseño de parcelas divididas distribuidas al azar. En el cual la parcela principal fue la salinidad [con NaCl o con solución nutritiva estándar (SNS)] y la subparcela fue la aplicación de Si (0, 4, 8 g L-1). La medición de las variables se realizó durante el día. Los resultados mostraron que la temperatura de la hoja se incrementó al amanecer en plantas con NaCl y al atardecer el efecto se invirtió y los valores mayores fueron en plantas con SNS. Al medio día, se identificó interacción Si × S en las variables de eficiencia fotosintética (Fv/Fm) y fluorescencia mínima (Fo). La fluorescencia máxima (Fm) resultó más afectada por el tipo de salinidad, presentando los valores mayores plantas con SNS. Se concluyó que la respuesta fisiológica diurna de la planta de tomate cultivado bajo condiciones salinas dependerá del tipo de sal empleada y de la dosis de Si, sobretodo en horas tempranas y tardías del día.
Palabras clave: cloruro de sodio; estrés salino; fisiología vegetal; invernadero; nutrición mineral
Introduction
Soil salinity is the main abiotic limiting factor in the productivity of agricultural crops especially in arid and semi-arid regions due to low rainfall, high temperatures and high evapotranspiration, in addition to poor soil and water management practices (Munns and Tester, 2008). Around 20% of the total arable area is affected by this factor with an annual increase of 1 to 2% due to high concentrations of salts (Munns and Tester, 2008; Plaut et al., 2013; Rizwan et al., 2015).
Salinity affects plant growth by being associated with low osmotic potential of the soil solution (water stress), ionic stress and nutritional imbalance (Munns and Tester, 2008; Parvaiz and Satyawati, 2008; Horie et al., 2012). High concentrations of toxic ions such as Na+ and Cl- form an ionic imbalance in the plant cell, affect the physiological and enzymatic processes causing alterations in the metabolism and decrease the absorption of essential ions such as K+ and Ca2+ (Hajiboland et al., 2010).
Saline stress mainly affects the process of photosynthesis by damaging the photosynthetic apparatus. Not only does it affect the opening and closing of stomata, but it also decreases the assimilation of CO2 (Mehta et al., 2010; Ashraf and Harris, 2013; Gupta and Huang, 2014). In addition, it damages photosynthetic pigments including chlorophyll and carotenoids and the leaf area (Brugnoli and Lauteri, 1991; Gong et al., 2005). On the other hand, an immediate response of the plant when exposed to high concentrations of salinity is the stomatic closure (Tang et al., 2015).
Although stomatic closure implies a reduction in CO2 fixation, under fluorescence conditions it can cause overexcitement of the photosystem II (PSII) reaction centers (Ahmed et al., 2009; Ashraf and Harris, 2013). Likewise, when plants are affected by high concentrations of salinity, they deteriorate the complex and reaction centers of PSII (Naumann et al., 2010). So that studying the impact of abiotic stress such as salinity, drought, low and high temperatures in the photosynthetic efficiency of the plant, is through the measurement of chlorophyll fluorescence (Zobayed et al., 2005) and gas exchange in the leaves (Shahid et al., 2011; Li et al., 2015).
Chlorophyll fluorescence is a non-destructive measure, easy to use and immediate response. This method provides information on the identity of several pigments, their structure and the reactions of electron transfers specific to PSII (Ashraf and Harris, 2013).
Hattori et al. (2005) found a higher photosynthetic rate, perspiration, stomatal conductance, larger stomata in sorghum plants (Sorghum bicolor L.) grown under saline stress conditions when treated with Si, compared to plants that did not receive Si. Similar results were found in corn cultivars (Zea mays) (Parveen and Ashraf, 2010) and in okra plants (Albemoschus esculentus) (Abbas et al., 2015).
On the other hand, Chen et al. (2011) reported an increase in the photochemical efficiency of PSII, photosynthetic rate and perspiration rate in rice plants (Oryza sativa L.) fertilized with Si under conditions of saline stress. On the other hand, Wang et al. (2015) found a higher photosynthetic rate and perspiration, water content in the leaves and hydraulic conductance of the root in cucumber plants fertilized with Si in salinity conditions. Similar results have been reported by Shi et al. (2016) in tomato plants. All these researchers concluded that the Si applied to the cultures induced a decrease in Na+ and an increase in the concentration of K+ in the leaves.
Si is the second most abundant mineral element in the soil after oxygen and forms 31% of the earth's crust (Gong et al., 2006; Epstein, 2009). In the soil solution it is in the form of monosilicic acid (H4SiO4) in concentrations ranging from 0.1 to 0.6 mM (Epstein, 1999; Ma and Yamaji, 2008). H4SiO4 is absorbed by plants and transported in the plant through the xylem (Epstein, 1999; Kazunori and Ma, 2003). Later it is deposited on the cell wall as amorphous silica (SiO2 nH2O) interacting with pectins and calcium improving its stiffness and resistance (Epstein, 1999; Ma, 2004; Ma and Yamaji, 2008).
Plants contain Si in concentrations of 0.1 to 10% dry weight (Ma and Yamaji, 2008). Si acts as a physical and mechanical barrier in plants. Not only is it deposited on cell walls, but also participates in metabolic and physiological activities when plants are under stress conditions (Ma, 2004; Liang et al., 2007). Although, Si is not considered an essential element for plant growth and metabolism, it is beneficial under stressful conditions (Liang et al., 2007; Ma and Yamaji, 2008; Epstein, 2009; Kaur et al., 2016). Some authors have reported tolerance to salinity by fertilizing Si in different crops such as rice, wheat (Triticum durum), tomato, cucumber, barley (Hordeum vulgare L.).
Most Si is deposited in the epidermal cells of the roots, leaves and stems, reducing the absorption of Na+ by the roots, Si decreases the permeability of cell membranes to Na+, resulting in a low level of Na+ and a high level of K+ in the cytosol (Gong et al., 2006). The Si function reduces the absorption of Na+ by decreasing perspiration in rice (Yeo et al., 1999; Gong et al., 2006), improves the hydric state of tomato (Romero-Aranda et al., 2006), increases activity enzyme antioxidant in cucumber (Zhu et al., 2004) and increases the activity of the H+ ATPase of the plasma membrane in barley (Liang et al., 2006). The objective of this study was to determine the effect of Si nutrition in tomato grown under conditions of saline stress induced with NaCl or with standard nutrient solution (SNS), on leaf temperature, photochemical efficiency and stomatal conductance.
Materials and methods
Establishment of the experiment and environmental conditions
The experiment was conducted in the experimental field of the Institute of Agricultural Sciences of the Autonomous University of Baja California located in the common Nuevo Leon, Mexicali in the period from March to June 2017. A rooftop greenhouse with a polycarbonate roof was used. Low technology, no temperature control and no heating. The Amalia tomato saladette cultivar was used. The transplant was performed on March 31 using two plants per 9 L pots. The substrate used was stream sand with granulometry less than 0.5 cm. The application of the nutrient solution was through pressurized irrigation spaghetti type with an expense per dropper of 100 mL min-1. There were between one and three irrigations per day. The irrigation drainage was adjusted to 30%.
Experimental design
The experimental design used was divided plots randomly distributed with five repetitions. The main plot was two salinity conditions at a value of 4 dS m-1 (SNS and saline solution induced with NaCl) and the subplots were doses of Si (0, 4, 8 g L-1 of SiO2), for a total of twelve treatments As a source of Si, 94% silicon dioxide was used (Diatomix, Bio Agrinor®, Zapopan, Jalisco Mexico). Each main plot consisted of 12 plants, while the subplot was 4 plants.
Treatment Management
The SNS used was the one recommended by Gomez and Sanchez (2003) and the following nutrient concentrations N 224, P 47, K 281, Ca 212, Mg 65, Fe 2, Mn 0.55, Zn 0.33, Cu 0.05, were handled. B 0.28 and Mo 0.05 mg L-1. At the time of transplantation, all SNSs were sufficiently diluted in water to reach an electrical conductivity of 2 dS m-1.
After 15 days after the transplant, the application of the treatments began. For factor one, in the plots of the SNS treatment, a gradual increase was made for two days in the concentration of the macro and micronutrients until a salinity value of 4 dS m-1 was reached. The same happened in the salinity treatment with NaCl. The increase in salinity of the solution was achieved by adding in addition to the SNS to an electrical conductivity of 2.0 dS m-1, the amount of ≈2.6 g L-1 of NaCl and reaching the value of 4 dS m-1. On the other hand, the doses of Si were applied daily to each pot manually at 12:00 noon.
The pH of the nutrient solution was maintained between 5.5-6 units. During the time the experiment lasted, the relative humidity of the greenhouse was achieved by wetting the floor with running water at 10:00 and 14:00 h. The maximum temperature inside the greenhouse at 15 hours a day reached 45 °C and the relative humidity at that same time was 14.5% and were monitored using a CEM DT-172 digital thermohygrometer (Twin Light Instruments, Monterrey, Mexico) placed at 20 cm just above the plants.
Variables evaluated
The physiological variables evaluated were leaf temperature, photochemical efficiency and stomatal conductance. The measurements were made at 34 days after the transplant at 9:00, 11:00, 13:00, 15:00 and 17:00 a day. For all variables, the fourth or fifth leaf counted from the apex down was used as a reference. All measurements were made on three plants per previously labeled treatment.
Leaf temperature
The leaf temperature was determined with an infrared digital laser gun thermometer (DAN-tronics model P045440, Mexico). The measurements were made at a distance of 20 cm between the sensor and the leaf.
Photochemical efficiency
The maximum photochemical efficiency of PSII (Fv/Fm), minimum fluorescence (Fo) and maximum fluorescence (Fm) was determined with a portable fluorometer (Chlorophyll Fluorometer OS-30p, OPTI-SCIENCE, USA) using an actinic light intensity of 2 100 µmol photon m-2 s-1 intensity for a period of 2 s. Prior to the determination, the leaves were acclimatized for 30 min of darkness with light exclusion clamps to ensure that all reaction centers were open (González et al., 2008).
Results and discussion
Leaf temperature
A response in the temperature of the leaves of the tomato crop was not identified by the Si × S interaction (p> 0.05) (Table 1). Likewise, the addition of Si also showed no significant effect on leaf temperature in all treatments (p> 0.05). However, salinity treatments did show response to the increase in leaf temperature (p> 0.05). At 9:00 am, the application of NaCl, increased the leaf temperature by more than 2 °C than plants treated with SNS-induced salinity, while at 1:00 p.m. and 5:00 p.m. was inverted.
Table 1 Effect of the application of Si in two forms of salinity (SNS and NaCl) on the temperature of the hydroponic tomato leaf.
Factor |
9:00 h |
11:00 h |
13:00 h |
15:00 h |
17:00 h |
Temperature (°C) | |||||
Silicon (Si) |
|
|
|
|
|
0 |
31.8 |
34.76 |
38.38 |
36.74 |
35.79 |
4 |
31.59 |
34.85 |
38.3 |
36.72 |
35.67 |
8 |
31.33 |
34.91 |
38.35 |
36.64 |
35.7 |
Probability |
0.72 |
0.938 |
0.718 |
0.954 |
0.537 |
Salinity (S) |
|
|
|
|
|
SNS 4 dS m-1 |
30.43 |
34.62 |
39.84 |
36.82 |
35.92 |
NaCl 4 dS m-1 |
32.71 |
35.06 |
36.85 |
36.58 |
35.51 |
Probability |
< 0.001 |
0.212 |
< 0.001 |
0.414 |
< 0.001 |
Si × S probability |
0.79 |
0.775 |
0.594 |
0.963 |
0.95 |
In this sense, the plants subjected to SNS-induced salinity had the ability to stay cooler in the morning due to greater water availability or better perspiration (Shahenshah and Isoda, 2010); however, in the afternoon the plants with NaCl remained fresher than those salinized with SNS. The above happens when the crop is subjected to water stress due to lack of water in the substrate or a poor root system (Sánchez-Blanco et al., 2014). In either case, in this study salinity restricted stomatal conductance and consequently foliar temperature increased (Ben-Asher et al., 2006).
Maximum photochemical efficiency of PSII (Fv/Fm)
Table 2 shows the results of maximum photochemical efficiency of PSII (Fv/Fm) due to the application of Si and two forms of salinity. The Si × S interaction was significant between 11:00 am and 3:00 pm (p≤ 0.004). At that same time, the application of Si had a significant effect on the Fv/Fm of the leaves of tomato plants (p≤ 0.005). Throughout the day, the salinity caused by the SNS significantly affected the Fv/Fm (p≤ 0.001) maintaining higher values than with the salinity caused by NaCl.
Table 2 Effect of the application of Si in two forms of salinity (SNS and NaCl) on the maximum photochemical efficiency of PSII (Fv/Fm) in hydroponic tomato leaves.
Factor |
9:00 h |
11:00 h |
13:00 h |
15:00 h |
17:00 h |
Fv/Fm | |||||
Silicon (Si) |
|
|
|
|
|
0 |
0.75 |
0.741 |
0.672 |
0.749 |
0.734 |
4 |
0.742 |
0.714 |
0.713 |
0.726 |
0.729 |
8 |
0.741 |
0.683 |
0.624 |
0.695 |
0.727 |
Probability |
0.364 |
˂ 0.001 |
0.002 |
0.005 |
0.615 |
Salinity (S) |
|
|
|
|
|
SNS 4 dS m-1 |
0.821 |
0.797 |
0.745 |
0.795 |
0.815 |
NaCl 4 dS m-1 |
0.667 |
0.628 |
0.595 |
0.652 |
0.646 |
Probability |
˂ 0.001 |
˂ 0.001 |
˂ 0.001 |
˂ 0.001 |
˂ 0.001 |
Si × S Probability |
0.146 |
˂ 0.001 |
˂ 0.001 |
˂ 0.004 |
0.134 |
The reduction in the Fv/Fm values in the plants subjected to salinity with NaCl, would indicate a reduction in the photochemical efficiency of the PSII and a disturbance or damage to the photosynthetic apparatus caused by the specific salinity of the NaCl and not by that caused by the SNS (Jimenez-Suancha et al., 2015). The interactive Si × S effect at 11, 13 and 15:00 h, was characterized by high Fv/Fm values when using the higher dose of Si (Figure 1). At 11:00 h, no difference was found between the type of salinity when Si was not applied. However, as used, if the difference in Fv/Fm was smaller in salinity induced by NaCl. At 13:00 the same effect was repeated, but only with the highest dose of Si [(8 g) (Figure 1b)]. Likewise, at 15:00 (Figure 1c) the effect of Si at doses of 4 and 8 g was the same as during the morning of the day.
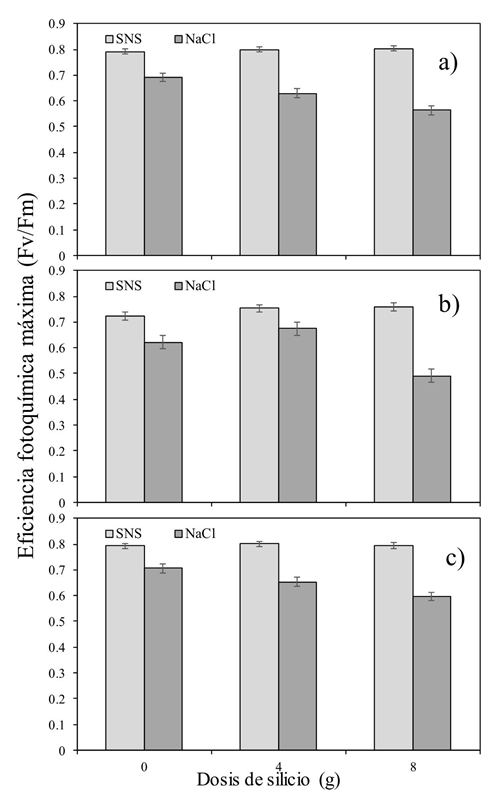
Figure 1 Silicon versus salinity (Si × S) interactive effect on the maximum photochemical efficiency of tomato cultivation during the day [a) 11:00 h; b) 13:00 h; and c) 15:00 h].
In general, lower Fv/Fm values were found at higher doses of Si, especially in the treatment of salinity induced by NaCl. These results agree with those reported by Cao et al. (2015), who, when applying Si to tomato seedlings subjected to saline stress, found that as time progressed, the Fv/Fm values were lower than those obtained in plants without the addition of Si.
Minimum fluorescence (Fo)
Table 3 shows the effect of the application of Si and the type of S on the Fo in tomato crop leaves. A significant effect was found in the interaction between Si × S for most of the day (p˂ 0.001). The highest dose of Si (8 g) applied to the NaCl solution was what made the difference from the rest of the treatments (Figure 2). Possibly the combination of the high dose and the NaCl caused that reaction. High values of Fo mean damage to the PSII reaction center or a reduction in the ability to transfer the excitation energy from the antenna to the reaction center (Baker, 2008; Khan et al., 2016). Contradictory data are those reported by Maghsoudi et al. (2015) who studied the Fo on the application of Si in wheat under water stress. They explain that the stress caused by the lack of water decreased Fo values.
Table 3 Effect of the application of Si in two forms of salinity (SNS and NaCl) on the minimum fluorescence (Fo) in hydroponic tomato leaves.
Factor |
9:00 h |
11:00 h |
13:00 h |
15:00 h |
17:00 h |
Fo | |||||
Silicon (Si) |
|
|
|
|
|
0 |
252.7 |
248.1 |
267.6 |
225.7 |
208 |
4 |
264.2 |
255.1 |
257.5 |
229.5 |
202 |
8 |
342.4 |
364.9 |
336.5 |
226 |
236.6 |
Probability |
˂ 0.001 |
˂ 0.001 |
0.003 |
0.913 |
0.007 |
Salinity (S) |
|
|
|
|
|
SNS 4.0 dS m-1 |
263.5 |
252.7 |
272 |
225.2 |
205 |
NaCl 4.0 dS m-1 |
309.3 |
326 |
302.3 |
229 |
226.1 |
Probability |
0.019 |
0.004 |
0.094 |
0.651 |
0.02 |
Si × S probability |
˂ 0.001 |
˂ 0.001 |
˂ 0.001 |
0.409 |
˂ 0.001 |

Maximum fluorescence (Fm)
Significant differences were observed in the Si factor and in the Si × S interaction, over the Fm at 9:00 and 15:00 h (p˂ 0.05) (Table 4). Additionally, the salinity caused by the addition of NaCl increased the Fm reflex values in the reduction of ‘closure’ of the PSII reaction centers (González et al., 2008), which in turn implies a greater activation of the reactions photochemicals (Jiménez-Suancha et al., 2015). This reinforces the hypothesis that the salinity damage presented in crops is the result of the use of NaCl rather than a balanced nutrient solution (SNS).
Table 4 Effect of the application of Si in two forms of salinity (SNS and NaCl) on the maximum fluorescence (Fm) in hydroponic tomato leaves.
Factor |
9:00 h |
11:00 h |
13:00 h |
15:00 h |
17:00 h |
Fm | |||||
Silicon (Si) |
|
|
|
|
|
0 |
1335.9 |
1155.3 |
1057.4 |
1010.1 |
1089 |
4 |
1298.7 |
1099.8 |
966.7 |
914.1 |
1037.9 |
8 |
1440.8 |
1172.8 |
956.2 |
1043.6 |
1067.5 |
Probability |
0.016 |
0.278 |
0.083 |
0.003 |
0.687 |
Salinity (S) |
|
|
|
|
|
SNS 4 dS m-1 |
1472.3 |
1257.6 |
1099.3 |
1122.5 |
1114.4 |
NaCl 4 dS m-1 |
1244.7 |
1027.7 |
1087.3 |
856 |
1015.2 |
Probability |
˂ 0.001 |
˂ 0.001 |
˂ 0.001 |
˂ 0.001 |
0.053 |
Si × S probability |
0.007 |
0.392 |
0.088 |
0.003 |
0.221 |
Stomatic conductance (gs)
Table 5 shows the Si × S interaction at gs (p ˂ 0.037) at the end of the day. Figure 3c indicates that when NaCl was applied without Si, the gs was increased while the salinity produced by the SNS kept low gs values even when Si was applied. On the other hand, Si significantly decreased GS at 9:00 h. While the salinity caused by the addition of NaCl, it increased at 9:00 and 17:00 in about 120 mmol m-2 s-1. Contrary to this study, Savvas et al. (2009) found that fertilization with Si promoted an increase in gs in zucchini plants (Cucurbita pepo L. cv. ‘Rival’) when they were subjected to a salinity of 6.2 dS m-1, with NaCl. On the other hand, Romero-Aranda et al. (2006) found no differences in the gs of tomato seedlings when they applied different combinations of NaCl and Si in Hoagland nutrient solutions to the Si application. However, when plants grown under salinity (NaCl) were treated with Si, the water content in them increased by 40%.
Table 5 Effect of the application of Si in two forms of salinity (SNS and NaCl) on stomatal conductance (gs) in hydroponic tomato leaves.
Factor |
9:00 h |
11:00 h |
13:00 h |
15:00 h |
17:00 h |
(mmol m-2 s-1) | |||||
Silicon (Si) |
|
|
|
|
|
0 |
810.2 |
915.2 |
953 |
873.4 |
796.8 |
4 |
794.7 |
912 |
967.9 |
948.2 |
703.4 |
8 |
640.5 |
833.3 |
991.6 |
887.1 |
732.4 |
Probability |
0.002 |
0.5 |
0.617 |
0.307 |
0.237 |
Salinity (S) |
|
|
|
|
|
SNS 4 dS m-1 |
679.9 |
828.5 |
966.7 |
864.3 |
681.1 |
NaCl 4 dS m-1 |
817 |
945.2 |
975 |
941.5 |
807.4 |
Probability |
0.002 |
0.081 |
0.798 |
0.076 |
0.01 |
Si × S probability |
0.712 |
0.539 |
0.793 |
0.07 |
0.037 |
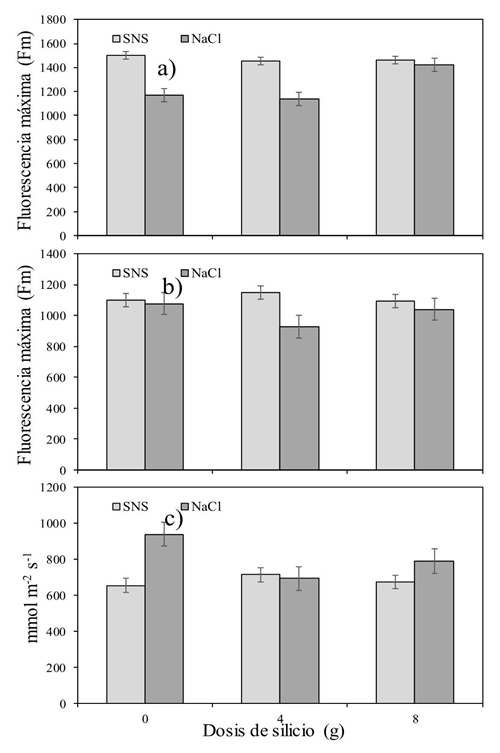
Figure 3 Silicon versus salinity (Si × S) interactive effect on the maximum chlorophyll fluorescence (Fm). a): 9:00 h; b) 15:00 h] and stomatal conductance; and c) 17:00 h] in the tomato crop.
The present investigation showed the response of the plant to two types of salinity and the application of Si. The use of SNS-induced salinity resulted in an increase in Fv/Fm expressed as light absorbed by PSII only early and late in the day. This behavior was the result of high values in Fo and low values in Fm caused by NaCl. Zribi et al. (2008) found similar results to those of this work. They mention that there is a relationship between the use of NaCl and the concentration of Na+ in the leaf tissue of tomato plants. The presence of Na+ in the leaf tissue has a negative impact on the amount of electron transport, as well as the photochemical efficiency of PSII.
The behavior of the gs had the same response to the type of salinity as the Fo. Romero-Aranda et al. (2006) presented similar values to those found in this study, but without deference for salinity or the addition of Si. In subsequent studies, the role of NaCl and the increase in gs should be studied in more detail, because the results in this study do not allow obtaining a clear role response of this type of salt. The same should happen with the foliar temperature response to this type of salinity.
Si failed to positively affect leaf temperature and most of the day negatively affected Fv/Fm. The interactive effect of Si and the type of salt made it clear that the dose of 8 g pot-1 day-1 impaired Fv/Fm instead of helping it (Figure 1). Something similar happened with the variable Fo. Which was increased by using the combination of Si at a dose of 8g and the salinity induced by NaCl. The opposite case when measuring the Fm (Figure 3a and 3b). More studies will be necessary but using lower doses than those proposed in this experiment, in addition to considering a longer period of time than the one studied. This could make it possible to find more conclusive results.
Conclusions
The maximum photochemical efficiency was higher in the SNS and was negatively affected with the increase in the dose of Si in the NaCl treatment.
The minimum fluorescence was not modified with the application of Si in the SNS, but in the treatment with NaCl it was increased with the higher dose of Si.
The maximum fluorescence was higher in the treatment with SNS, while in the NaCl treatment it was lower especially in doses 0 and 4 g Si pot-1 day-1.
At an early time, the stomatal conductance was lower as the dose of Si increased. In addition, early and late of the day, the plants with NaCl had the highest values than the SNS.
Literatura citada
Abbas, T.; Balal, R. M.; Shahid, M. A.; Pervez, M. A.; Ayyub, C. M. and Aqueel, M. A. 2015. Silicon-induced alleviation of NaCl toxicity in okra (Abelmoschus esculentus) is associated with enhanced photosynthesis, osmoprotectants and antioxidant metabolism. Acta Physiol Plant. 37(6):1-15. [ Links ]
Ahmed, C. B.; Rouina, S. B. B.; Sensoy, M.; Boukhris, F. B. and Abdallah. 2009. Changes in gas exchange, proline accumulation and antioxidative enzyme activities in three olive cultivars under contrasting water availability regimes. Environ. Exp. Bot. 67(2):345-352. [ Links ]
Ashraf, M. and Harris, P. J. C. 2013. Photosynthesis under stressful environments: an overview. Photosynthetica. 51(2):163-190. [ Links ]
Baker, N. R. 2008. Chlorophyll fluorescence: a probe of photosynthesis in vivo. Ann. Rev. Plant Biol. 59(1):89-113. Doi: 10.1146/annurev.arplant. [ Links ]
Ben-Asher, J.; Tsuyuki, I.; Bravdo, B. A. and Sagih, M. 2006. Irrigation of grapevines with saline water I. Leaf area index, stomatal conductance, transpiration and photosynthesis. Water Management. 83(1-2):13-21. [ Links ]
Brugnoli, E. and Lauteri, M. 1991. Effects of salinity on stomatal conductance, photosynthetic capacity, and carbon isotope discrimination of salt-tolerant (Gossypium hirsutum L.) and salt-sensitive (Phaseolus vulgaris L.) C3 non-halophytes. Plant Physiol. 95(2):628-635. [ Links ]
Cao, B.; Ma, Q.; Zhao, Q.; Wang, L. and Xu, Q. 2015. Effects of silicon on absorbed light allocation, antioxidant enzymes and ultrastructure of chloroplasts in tomato leaves under simulated drought stress. Scientia Hortic. 194(1):53-62. [ Links ]
Chen, W.; Yao, X.; Cai, K. and Chen, J. 2011. Silicon alleviates drought stress of rice plants by improving plant water status, photosynthesis and mineral nutrient absorption. Biol Trace Elem Res. 142(1):67-76. [ Links ]
Epstein, E. 1999. Silicon, annual review of plant physiology and plant molecular biology. 50(1):641-64. [ Links ]
Epstein, E. 2009. Silicon: its manifold role in plants. Ann. Appl. Biol. 155(2):155-160. [ Links ]
Gómez, H. T. y Sánchez, F. D. 2003. Soluciones nutritivas diluidas para la producción de jitomate a un racimo. Terra Latinoam. 21(1):57-63. [ Links ]
Gong, H.; Zhu, X.; Chen, K.; Wang, S. and Zhang, C. 2005. Silicon alleviates oxidative damage of wheat plants in pots under drought. Plant Sci. 169(2):313-321. [ Links ]
Gong, H. J.; Randall, D. P. and Flowers, T. J. 2006. Silicon deposition in the root reduce uptake in rice (Oryza sativa L.) seedling by reducing bypass flow. Plant. Cell Environ. 29(10):1970-1979. [ Links ]
González M. S.; Perales, H. V. y Salcedo, A. O. M. 2008. La fluorescencia de la clorofila a como una herramienta en la investigación de efectos tóxicos en el aparato fotosintético de plantas y algas. Rev. Edu. Bio. 27(4):119-129. [ Links ]
Gupta, B. and Huang, B. 2014. Mechanism of salinity tolerance in plants: physiological, biochemical, and molecular characterization. Inter. J. Genomics. 14(1):1-18. http://dx.doi.org/10.1155/2014/701596. [ Links ]
Hajiboland, R.; Aliasgharzadeh, A.; Laiegh, S. F. and Poschenrieder, C. 2010. Colonization with Arbuscular mycorrhizal fungi improve salinity tolerance of tomato (Solanum lycopersicum L.) plants. Plant and Soil. 331(1):313-327. [ Links ]
Hattori, T. S.; Inanaga, H.; Araki, P.; An, S.; Morita, M.; Luxova´, A. and Lux. 2005. Application of silicon enhanced drought tolerance in Sorghum bicolour. Physiology Plantarum. 123(4):459-466. [ Links ]
Horie, T.; Karahara, I. and Katsuhara, M. 2012. Salinity tolerance mechanisms in glycophytes: an overview with the central focus on rice plants. Rice Sci. 5(1):11-24. [ Links ]
Jiménez-Suancha, S. C.; Alvarado, S. O. H y Balaguera-López, H. E. 2015. Fluorescencia como indicador de estrés en Helianthus annuus L. Una revision. Rev. Colomb. Cienc. Hortíc. 9(1):149-160. [ Links ]
Kaur, S.; Kaur, N.; Siddique, K. H. M. and Nayyar, H. 2016. Beneficial elements for agricultural crops and their functional relevance in defence against stresses. Arch. Agron. Soil Sci. 62(7):905-920. [ Links ]
Kazunori, T. and Ma, J. F. 2003. Characterization of silicon uptake by rice roots. New Phytologist. 158(3):431-436. [ Links ]
Khan, W. U. D.; Aziz, T.; Hussain, I.; Ramzani, P. M. A. and Reichenauer, T. G. 2016. Silicon: a beneficial nutrient for maize crop to enhance photochemical efficiency of photosystem II under salt stress. Arch. Agron Soil Sci. 63(5):599-611. Doi.org/10.1080/03650340. 2016.1233322. [ Links ]
Li, H.; Zhu, Y.; Hu, Y.; Han, W. and Gong, H. 2015. Beneficial effects of silicon in alleviating salinity stress of tomato seedlings grown under sand culture. Acta Physiol Plant. 37(4):1-9. [ Links ]
Liang, Y.; Zhang, W.; Chen, Q.; Liu, Y. and Ding, R. 2006. Effects of exogenous silicon (Si) on H+ ATPase activity, phospholipids and fluidity of plasma membrane in leaves of salt-stressed barley (Hordeum vulgare L.). Environ. Exp. Bot. 57(3):212-219. [ Links ]
Liang, Y.; Sun, T.; Zhu, W. Y. G. and Christie, P. 2007. Mechanisms of silicon-mediated alleviation of abiotic stresses in higher plants: A review. Environmental Pollution. 147(2):422-428. [ Links ]
Ma, J. F. 2004. Role of silicon in enhancing the resistance of plants to biotic and abiotic stresses. Soil Sci. Plant Nutr. 50(1):11-18. [ Links ]
Ma, J. F. and Yamaji, N. 2008. Functions and transport of silicon in plants. Cell. Mol. Life Sci. 65(19):3049-3057. [ Links ]
Maghsoudi, K.; Emam, Y. and Ashraf, M. 2015. Influence of foliar application of silicon on chlorophyll fluorescence, photosynthetic pigments, and growth in water-stressed wheat cultivars differing in drought tolerance. Turk. J. Bot. 39(6):625-634. [ Links ]
Mehta, P.; V. Kraslavsky, S.; Bharti, S. I.; Allakhverdiev, A. and Jajoo. 2010. Analysis of salt-stress induced changes in photosystem II heterogeneity by prompt fluorescence and delayed fluorescence in wheat (Triticum vulgare) leaves. J. Photochem. Photobiol. 104(2):308-313. [ Links ]
Munns, R. and Tester, M. 2008. Mechanisms of salinity tolerance. Annual Review of Plant Biology. 59(1):651-681. [ Links ]
Naumann, J. C.; Bissett, S. N.; Young, D. R.; Edwards, J. and Anderson, J. E. 2010. Diurnal patterns of photosynthesis, chlorophyll fluorescence, and PRI to evaluate water stress in the invasive species, Elaeagnusum bellata. Trees J. 24(2):237-245. [ Links ]
Parvaiz, A. and Satyawati, S. 2008. Salt stress and phytobiochemical physiological response of salt-stressed maize (Zea mays L.). Inter. J. Plant Biol. 40(3):1041-1052. [ Links ]
Parveen, N. and Ashraf, M. 2010. Role of silicon in mitigating the adverse effects of salt stress on growth and photosynthetic attributes of two maize (Zea mays L.) cultivars grown hydroponically. Pak. J. Bot. 42(3):1675-1684. [ Links ]
Plaut, Z.; Edelstein, M. and Ben-Hur, M. 2013. Overcoming salinity barriers to crop production using traditional methods. Critical Reviews in Plant Sci. 32(4):250-291. [ Links ]
Rizwan, M.; Ali, S. M.; Ibrahim, M.; Farid, M.; Adrees, S.A.; Bharwana, M.; Zia-ur-Rehman, M. F. and Qayyum, F. Abbas. 2015. Mechanisms of silicon-mediated alleviation of drought and salt stress in plants: a review. Environ. Sci. Pollut Res. 22(20):15416-15431. [ Links ]
Romero-Aranda, M. R.; Jurado, O. and Cuartero, J. 2006. Silicon alleviates the deleterious salt effect on tomato plant growth by improving plant water status. J. Plant Physiol. 163(8):847-855. [ Links ]
Sánchez-Blanco, M. J.; Álvarez, S.; Ortuño, M. F. and Ruiz-Sánchez, M. C. 2014. Root system response to drought and salinity: root distribution and water transport. In: Root Engineering. Morte, A. and Varma, A. (Ed.). Soil Biology 40. Heidelberg: Springer. 325-352 pp. [ Links ]
Savvas, D.; Giotis, D.; Chatzieustratiou, E.; Bakea, M. and Patakioutas, G. 2009. Silicon supply in soilless cultivations of zucchini alleviates stress induced by salinity and powdery mildew infections. Environ. Exp. Bot. 65(1):11-17. [ Links ]
Shahenshah e Isoda, A. 2010. Effects of water stress on leaf temperature and chlorophyll fluorescence parameters in cotton and peanut. Plant Prod. Sci. 13(3):269-278. Doi: 10.1626/pps.13.269. [ Links ]
Shahid, M. A.; Pervez, M. A.; Balal, R.M.; Ahmad, R.; Ayyub, C. M.; Abbas, T. and Akhtar, N. 2011. Salt stress effects on some morphological and physiological characteristics of okra (Abelmoschus esculentus L.). Soil Environ. 30(1):66-73. [ Links ]
Shi, Y.; Zhang, Y.; Han, W.; Feng, R.; Hu, Y.; Guo, H. and Gong. 2016. Silicon enhances water stress tolerance by improving root hydraulic conductance in Solanum lycopersicum L. Frontiers in Plant Sci. 7(2):196-211. [ Links ]
Tang, X.; Mu, X.; Shao, H.; Wang, H. and Brestic, M. 2015. Global plant-responding mechanisms to salt stress: Physiological and molecular levels and implications in biotechnology. Critical Reviews in Biotechnol. 35(4):425-437. [ Links ]
Wang, S.; Liu, P.; Chen, D.; Yin, L.; Li, H. and Deng, X. 2015. Silicon enhanced salt tolerance by improving the root water uptake and decreasing the ion toxicity in cucumber. Frontiers in Plant Sci. 6:759-769. DOI=10.3389/fpls.2015.00759. [ Links ]
Yeo, A. R.; Flowers, S.A.; Rao, G.; Welfare, K.; Senanayake, N. and Flowers, T. J. 1999. Silicon reduces sodium uptake in rice (Oryza sativa L.) in saline conditions and this is accounted for by a reduction in the transpiration bypass flow. Plant Cell Environ. 22(5):559-565. [ Links ]
Zhu, Z.; Wei, G.; Li, J.; Qian, Q. and Yu, J. 2004. Silicon alleviates salt stress and increase antioxidant enzymes activity in leaves of salt stressed cucumber (Cucumis sativus L.). Plant Sci. 167(3):527-533. [ Links ]
Zobayed, S. M. A.; Afreen, F. and Kozai, T. 2005. Temperature stress can alter the photosynthetic efficiency and secondary metabolite concentrations in St. John’s Wort. Plant Physiol. Biochem. 43(10):977-984. [ Links ]
Zribi, L.; Fatma, G.; Fatma, R.; Salwa, R.; Hassan, N. and Néjib, R. M. 2008. Application of chlorophyll fluorescence for the diagnosis of salt stress in tomato ‘‘Solanum lycopersicum (variety Rio Grande)’’. Sic. Hortic. 120(3):367-372. [ Links ]
Received: January 01, 2020; Accepted: March 01, 2020
 Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
