











 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroducción
El salivazo, Aeneolamia albofasciata (Lallemand) (Hemiptera Cercopidae), es una de las plagas más dañinas de la caña de azúcar. Se presenta en extensas áreas del Golfo de México y zona costera del Océano Pacífico (Flores-Cáceres, 1994; Peck, 2001; Gómez, 2007; López-Collado et al., 2013, Alatorre-Rosas y Hernández-Rosas, 2015). Alatorre-Rosas et al. (2013); Alatorre-Rosas y Hernández-Rosas (2015) mencionan que A. albosfaciata presenta una distribución altitudinal que va desde 10 a 1 700 msnm. Los efectos generados por la alimentación de ninfas y adultos ocasionan estrés hídrico, retrasa el crecimiento de la planta, reduciendo la producción de biomasa (Dinardo-Miranda et al., 2000; Valerio et al., 2001; Alatorre-Rosas y Hernández-Rosas, 2015). A. albofasciata ocasiona pérdidas hasta de 9 t ha-1 de caña de azúcar (De la Cruz et al., 2005; Alatorre-Rosas y Hernández-Rosas, 2015).
Las ninfas aparecen pocos días después del inicio de las lluvias y se prolongan durante todo el periodo lluvia, apareciendo picos poblacionales de ninfas y adultos de manera escalonada y superpuestas. Las ninfas recién eclosionadas inician la producción de saliva, misma que las cubre totalmente, les confiere refugio, sirve como defensa contra sus enemigos naturales y al mismo tiempo de protección contra condiciones climáticas adversas (Marshall, 1966; Whittaker, 1970; Bodegas, 1973; Alatorre-Rosas y Hernández-Rosas, 2015).
Para el control de adultos se recomienda el uso de insecticidas en infestaciones severas y del hongo entomopatógeno Metarhizium anisopliae (Metschnikoff) Sorokin (Ascomycota: Hypocreales) (Torriello et al., 2008; Hernández-Rosas et al., 2013; Alatorre-Rosas y Hernández-Rosas, 2015). El uso de M. anisopliae como insecticida biológico se ha generalizado en diferentes áreas cañeras, para la regulación de adultos (Flores- Cáceres, 1994; Berlanga et al., 1997; Badilla, 2002; Carballo y Falguni, 2004; Castillo-Zeno 2006; Bustillo y Castro, 2011; Alatorre-Rosas y Hernández-Rosas, 2015). A la fecha, se cuenta con formulados en polvo humectable de inertes con esporas de M. anisopliae y aislamientos nativos de nematodos entomopatógenos para la regulación de poblaciones de ninfa o salivazo, debido que la saliva es una barrera efectiva contra enemigos naturales y otras estrategias de control (Alatorre-Rosas y Hernández-Rosas, 2015; Grifaldo-Alcántara et al., 2017).
Los nematodos entomopatógenos de las familias Steinernematidae y Heterorhabditidae han generado interés como estrategia de control biológico de ninfas de diferentes especies de salivazo, en este sentido las especiesSteinernema spp. y Heterorhabditis spp. contra el salivazo, Mahanarva fimbriolata han logrado mortalidades entre 96 y 100% bajo condiciones de laboratorio (Leite et al., 2002). Estudios de campo, demostraron que Heterorhabditis spp. en dosis de 5 x 1010 JI/h ocasionó mortalidades de 42.3% y con H. bacteriophora en dosis de 1.5 x 1011 JI/ha, la mortalidad sobre ninfas de A. varia fue 76% (Moreno et al., 2012).
Los factores abióticos pueden tener un efecto positivo o negativo en el desempeño, efectividad y nivel de éxito de los nematodos entomopatógenos en el control de insectos plaga. Es por ello que la aplicación de nematodos entomopatógenos puede resultar impredecible en condiciones naturales. En este sentido (Kaya, 1990; Kaya y Koppenhöfer, 1996; Smith, 1996) mencionan la importancia de tomar en cuenta los factores intrínsecos: comportamiento y fisiológicos, así como temperatura, humedad, textura de suelo y radiación (extrínsecos), además de factores bióticos como la competencia, que afectan su persistencia.
Por tal razón, resulta importante la realización de este estudio con algunos indicadores que permitan evaluar el tipo de suelo, la diversidad biológica y determinar el efecto sobre la permanencia de los nematodos entomopatógenos. El objetivo de esta investigación fue evaluar la patogenicidad de aislamientos nativos de Steinernema spp. (CPVC12; CPVC13) y Heterorhabditis sp. (CPVG13) y su persistencia en suelo procedentes de tres ingenios azucareros del estado de Veracruz.
Material y métodos
Susceptibilidad de ninfas de A. albofasciata hacia nematodos entomopatógenos ninfas de A. albofasciata
Las ninfas de tercer y cuarto instar de A. albofasciata fueron colectadas de campo con caña cultivada en Atoyaquillo del ingenio El Potrero, coordenadas 18° 59’ 38.74 latitud norte y 96° 46’ 50 longitud oeste. Las ninfas fueron colectadas con apoyo de espátulas de acero inoxidables, fueron separadas por género y colocadas en forma individual en placas de plástico con cavidades, que contenían en su interior papel toalla humedecido y estolones de pasto estrella (Cynodon sp.). Una prueba preliminar con estolones de pasto mostró que en la zona de los nudos se forman raicillas de diferente longitud, que permanecen activas durante varios días (10-12). Se consideró que si las ninfas podían adherirse a las raicillas y mantenerse al menos 4-5 días produciendo la excreción de saliva, constituiría un periodo favorable para los ensayos planteados en esta investigación.
Las ninfas colectadas fueron trasladadas al laboratorio y mantenidas en observación durante 24 h, fueron seleccionadas aquellas que sobrevivieron al manejo. Posteriormente, cada ninfa fue confinada individualmente en cajas Costar® (Corning Inc. NY USA) con 6 cavidades, cubiertas con papel filtro humedecido y un estolón de pasto (como alimento). Durante el experimento, las ninfas fueron mantenidas a temperatura ambiente. Las ninfas que sobrevivieron y continuaron produciendo saliva fueron usadas en el experimento, debido a su adaptación al espacio y cambio de alimento.
Nematodos entomopatógenos
Los aislamientos de nematodos utilizados para esta investigación fueron obtenidos de larvas de G. mellonella que fueron usadas como insecto cebo en muestras de suelo procedentes de tres ingenios azucareros del estado de Veracruz, México (Central Progreso, Constancia y La Gloria). Los experimentos fueron llevados a cabo con dos aislamientos de Steinernema sp. (CPVC12, CPVC13) y uno de Heterorhabditis sp. (CPVG13) (Cuadro 1). La determinación taxonómica morfológica y molecular de los aislamientos se encuentra en proceso y se reportará en un artículo posterior.
Cuadro 1 Nematodos utilizados para la prueba de patogenicidad.
| Tratamiento | UE (una placa) | Réplicas (placas) | Ninfas totales (réplica) | Repeticiones en tiempo | Total ninfas/UEs | Total de placas |
| 1. Steinernema (CPV12) | 1 | 4 | 24 | 2 | 48 | 8 |
| 2. Steinernema sp. (CPVC13) | 1 | 4 | 24 | 2 | 48 | 8 |
| 3. Heterorhabditis (CPVG13) | 1 | 4 | 24 | 2 | 48 | 8 |
| 4. Control (sin nematodos) | 1 | 4 | 24 | 2 | 48 | 8 |
| Totales | 16 | 96 | 192 | 32 |
Los nematodos fueron propagados sobre larvas de ultimo instar de G. mellonella (L.) y cosechados como juveniles infectivos (JI) mediante el método de trampas White (Kaya y Stock, 1997), se esterilizaron con cloruro de benzalconio al 0.01% y dos cambios de agua destilada estéril. La suspensión de juveniles infectivos (JI) se centrifugó a 500 rpm durante 5 min, con el fin de eliminar los nematodos muertos por decantación del sobrenadante. Los nematodos se conservaron a temperatura ambiente, en agua destilada, oxigenándolos con una bomba de pecera continuamente para disminuir la mortalidad. La concentración de JI en suspensión fue estimada usando alícuotas de 20 µL, utilizando el promedio de 7 conteos, ajustando la concentración requerida en la experimentación.
Procedimiento experimental
El proceso de inoculación de los diferentes aislamientos fue el mismo (Cuadro 1). Se utilizaron placas Costar® con 6 cavidades. Cada cavidad se cubrió con dos capas de papel filtro previamente esterilizado y humedecido. En cada cavidad se colocó un estolón (3.5 cm) de pasto estrella (Cynodon sp.) donde se indujo la formación de raicillas, sobre estos se colocó una ninfa de A. albofasciata. La inoculación se llevó a cabo aplicando una sola concentración de juveniles infectivos, 120 JI cm-2 de acuerdo a Alves et al. (2005). La aplicación se realizó sobre el papel filtro, permitiendo el desplazamiento de los JI y búsqueda del insecto hospedero.
Los tratamientos se incubaron a 25 ±2 °C y humedad relativa (74% min y 84% max) y fotoperiodo de 9 y 15 h luz-oscuridad. En el testigo, las ninfas fueron tratadas solamente con agua estéril. Las ninfas se revisaron cada 48 h y se determinó mortalidad por placa tomando en cuenta los síntomas (color rojo o beige, flacidez) de las ninfas. Cada ninfa muerta se retiró de la placa y se colocó en cámara húmeda, la cual consistió en una caja Petri con papel filtro humedecido, se determinó la emergencia de los JI por ninfa infectada. Los JI se colectaron durante un periodo de 4 a 5 días y se determinó el total de JI en 10 ml de la suspensión colectada por ninfa. Este valor se tomó como indicativo de la capacidad reproductiva de los nematodos. Cada tratamiento tuvo cuatro repeticiones (cuatro placas/24 ninfas) y todo el experimento se repitió en tres ocasiones diferentes. El experimento tuvo una duración de 7 días después de la aplicación de los juveniles infectivos. Toda la fase de laboratorio de realizó bajo condiciones asépticas.
Efecto del suelo con la persistencia de nematodos entomopatógenos
Se seleccionaron muestras de suelo de 8 sitios, dos pertenecientes al ingenio Constancia, dos a Central Motzorongo y cuatro a El Potrero. Las muestras de suelo fueron tomadas en un área de 30 x 30 cm de largo por ancho y de aproximadamente 10 cm de profundidad usando una pala. De cada sitio se obtuvieron trece muestras (un total de 5 kg) y fueron conservadas a 4 °C durante no más de dos semanas. Se prepararon muestras compuestas, combinando las 13 sub muestras de cada sitio por ingenio.
Para evitar el efecto de nematodos nativos en el experimento de persistencia, se llevó a cabo un muestreo en los suelos usando el método de insecto cebo con G. mellonella (Zimmerman, 1986). Cada muestra de suelo, fue humedecida con agua destilada estéril a capacidad de campo (60%). Se tomaron muestras de 500 g de cada localidad colocándolas en recipientes de plástico de 500 mL de capacidad (unidad experimental), siete larvas de G. mellonella del último instar fueron confinadas en forma individual en jaulas de malla metálica e introducidas en los recipientes con suelo y distribuidas al azar. Cada recipiente se cubrió con una tapa con perforaciones para permitir la aireación constante, los recipientes fueron invertidos (Miduturi y Moens, 1997) e incubados en una cámara de cría a 25 ±2 °C durante 7d, para confirmar la presencia de nematodos nativos. Este procedimiento se repitió dos veces.
Poblaciones microbianas cultivables
Dentro de la primera semana de colecta, las poblaciones naturales de bacterias, hongos y actinomicetos de cada muestra de suelo fueron estimadas. Muestras de 10 g de suelo de cada localidad, tamizadas en malla No. 10, se depositaron en un matraz Erlenmeyer de 250 mL conteniendo 90 mL de agua destilada estéril. Las muestras se agitaron durante 18 minutos en un agitador orbital (Thermolyne Cimarec® 3), para posteriormente preparar diluciones seriadas (Dhingra y Sinclair, 1985). Un factor final de dilución de 10-3 y 10-5 se consideró adecuado para el aislamiento de hongos, 10-4 y 10-6 para bacterias y actinomicetos 10-3 y 10-5.
Se realizó una siembra por extensión en placa en cajas Petri con agar nutritivo (NA) para bacterias, papa dextrosa agar (PDA) para hongos y medio agar Czapeck Dox con pH de 8.0 para actinomicetos con tres repeticiones por dilución. Las placas se incubaron a 27 °C/5d, 7d y 10d para las bacterias, hongos y actinomicetos, respectivamente. Después del periodo de incubación, el número de unidades formadoras de colonia (UFC) fue estimado seleccionando la dilución en cada medio que favorecía la enumeración adecuada (10-3 para bacterias y hongos y 10 -6 para actinomicetos). Las UFC/gramo de suelo seco (al aire) se calcularon al multiplicar el promedio aritmético del número de UFC por placa por el factor de dilución. El resultado se dividió entre el peso de suelo seco estimado. Las características físicas y químicas del suelo fueron llevadas a cabo por el Laboratorio de Fertilidad de Suelos del Programa de Edafología, Colegio de Postgraduados.
Procedimiento experimental
Para este experimento, se empleó suelo de los ingenios El Potrero, Central Motzorongo y Constancia, estos se seleccionaron porque no se registró la presencia de nematodos. Se emplearon dos aislamientos, CPVC12 (Steinernema spp.) y CPVG13 (Heterorhabditis sp.). El procedimiento fue el mismo para cada tipo de suelo y nematodo evaluado. Para cada combinación de suelo y nematodo, se prepararon 22 envases con 500 g de suelo cada uno, en el centro de 11 de los envases, se depositaron 5 000 juveniles infectivos, suspendidos en 2.5 mL de agua destilada estéril (ADE), en los 11 envases restantes únicamente se depositó 2.5 mL de ADE siendo estos el tratamiento testigo.
La supervivencia de los nematodos se cuantifico mediante la proporción de larvas de G. mellonella infectadas al estar en contacto con suelo inoculado por nematodos. Para esto, en dos envases, con y sin nematodos, se introdujeron en siete jaulas hechas de malla metálica en cada envase. Las jaulas de 1 cm de diámetro por 4 cm de longitud contenían una larva de G. mellonella. Siete días después, se repitió este procedimiento usando dos envases diferentes (con y sin nematodos) y así sucesivamente a los 14, 21, 28, 35, 42, 49, 56, 63 y 70d posteriores a la inoculación de los nematodos. El experimento se incubo a 25 °C en oscuridad total. Las larvas de G. mellonella se dejaron en contacto con el suelo durante 6d y después se retiraron del suelo y se cuantifico el número de larvas muertas.
Cada larva fue colocada en una cavidad de una placa de seis cavidades conteniendo papel filtro húmedo, e incubadas a las mismas condiciones experimentales. La presencia de juveniles infectivos (JI) fue determinada mediante observaciones entre los 10 a 15d. El experimento se realizó bajo un diseño completamente al azar, donde cada combinación de suelo/nematodo conto con dos replicas, y el experimento se repitió en dos ocasiones (cuatro repeticiones en total).
Los datos se analizaron usando regresión logística (asumiendo que los datos tuvieron una distribución binomial), donde cada número de larvas muertas fue una proporción del total de larvas evaluadas. Cuando fue necesario, la presencia de mayor dispersión de los datos bajo el supuesto de la distribución binomial se permitió, comparando la proporción de la desviación media de los tratamientos y la desviación media residual contra una distribución F, en lugar de comparar la desviación de los tratamientos con una distribución de χ2. Antes de comparar entre tratamientos, los resultados entre repeticiones en tiempo fueron comparados para poder ser combinados. Con los datos de las repeticiones combinadas, primero se comparó la supervivencia entre aislamientos de nematodos, entre los 4 tipos de suelo, entre los tiempos de evaluación y finalmente la interacción entre estos factores. El análisis se realizó en el programa GenStat v. 8 (Payne et al., 2005).
Resultados
Susceptibilidad de ninfas de A. albofasciata contra Heterorhabditis sp. y Steinernema spp.
De manera general se observaron diferencias en mortalidad causada por Steinernema spp. (CPVC12; CPVC13) y Heterorhabditis sp. (CPVG13). El porcentaje de mortalidad (62.5%) fue mayor para Heterorhabditis sp. a 25°C, comparado con Steinernema (CPVC12) (56.25%) y Steinernema sp. (CPVC13) (33.33%). A pesar de que se observó mortalidad en los tratamientos no fue posible realizar un análisis estadístico, ya que el tratamiento testigo mostro alta mortalidad por otras causas. Sin embargo, este experimento permitió demostrar que los aislamientos tanto de Steinernema como de Heterorhabditis tuvieron la capacidad de atravesar la saliva producida por las ninfas, penetrar la cavidad del insecto y causar infección (Figura 1b, 1c).
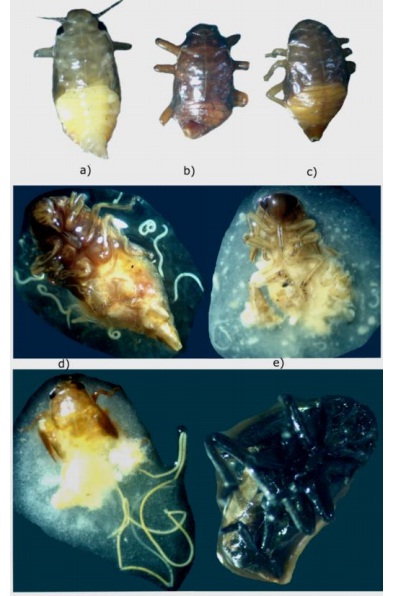
Figura 1 Ninfas de mosca pinta: a) ninfa sana; b) ninfa infectada por Heterorhabditis. (CPVG13); c) infectada por Steinernema (CPV12). Disecciones realizadas a ninfas parasitadas; d) nematodos adultos de (CPVG13) Heterorhbaditis; e) nematodos adultos de (CPV12) Steinernema; f) nematodos mermitidos; y g) ninfas con presencia de necrosis, posible presencia de bacterias.
Las disecciones de los cadáveres de ninfas infectadas permitieron verificar la capacidad reproductiva de los nematodos entomopatógenos (Figura 1d, 1e). Las ninfas expuestas a Heterorhabditis (CPVG13) y Steinernema sp. (CPVC12) mostraron diferencias en abundancia de juveniles infectivos. En el caso de Heterorhabditis la proporción más alta correspondió a 3 500 JI por ninfa (Cuadro 2).
Cuadro 2 Estimación del número de ninfas infectadas y JI encontrados en ninfas de A. albofaciata.
| Especie NEP/repetición | larvas infectadas | Población de JI/ninfa | |
| Mínima | Máxima | ||
| Steinernema R1 | 13 | 4 | 1 500 |
| Steinernema R2 | 14 | 7 | 1 500 |
| Heterorhabditis 1 | 12 | 1 000 | 3 500 |
| Heterorhabditis 2 | 18 | 10 | 3 500 |
| S. (Contancia) 1 | 6 | 9 | 1 000 |
| S. (Constancia) 2 | 10 | 4 | 1 000 |
| Control | 0 | - | - |
Poblaciones microbianas cultivables
La mayor población de bacterias y hongos se presentó en suelo de Constancia (3X109 UFC g-1 de suelo y 1.5 x 104 UFC g-1 de suelo para bacterias y hongos, respectivamente) (Figura 2a) mientras que en Central Motzorongo la proporción de bacterias (3 x 108 UFC) y hongos (4 x 104 UFC g-1 de suelo) fue considera baja (Figura 2b). La mayor población de actinomicetos se encontró en el sitio 2 del ingenio El Potrero (7 x 105 UFC g-1 de suelo) y la menor población se obtuvo en Constancia (8.6 x 104 UFC g-1 de suelo) y Motzorongo (1.7 x 195 UFC g-1 de suelo) (Figura 2c). Las características físicas y químicas del suelo (Cuadro 3).
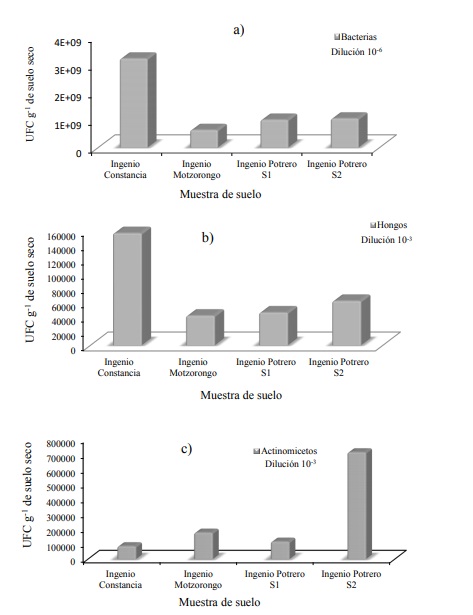
Figura 2 Unidades formadoras de colonias. a) bacterias; b) hongos; y c) actinomicetos, obtenidas de suelo de caña cultivada en el Ingenio Constancia, Central Motzorongo y El Potrero.
Cuadro 3 Características fisicoquímicas del suelo de tres ingenios azucareros.
| Identificación | pH | CE | MO (%) | N* | P | K | Textura Boyoucos |
| PotreroS1 | 4.8 | 0.32 | 1.7 | 0.09 | 41 | 0.4 | Arcilla |
| PotreroS2 | 5.1 | 0.28 | 5.4 | 0.27 | 64 | 0.7 | Arcilla |
| PotreroS3 | 6.4 | 0.26 | 3.3 | 0.16 | 12 | 0.3 | Franco arcilloso |
| PotreroS4 | 5.3 | 0.14 | 5.3 | 0.26 | 15 | 0.3 | Arcilla |
| ConstanciaS1 | 5.9 | 0.21 | 3.7 | 0.19 | 39 | 0.3 | Franco arcilloso |
| ConstanciaS2 | 6.5 | 0.16 | 5.3 | 0.26 | 31 | 0.4 | Arcilla |
| MotzorongoS1 | 5.2 | 0.21 | 2.2 | 0.11 | 23 | 0.6 | Franco arcilloso |
| MotzorongoS2 | 6 | 0.23 | 1.4 | 0.07 | 18 | 1 | Franco arcillo arenoso |
CE= conductibilidad eléctrica; MO= materia orgánica; N= nitrógeno; P= fósforo; K= potasio.
Efecto del suelo con la persistencia de nematodos entomopatógenos
No hubo diferencias significativas entre las cuatro repeticiones (
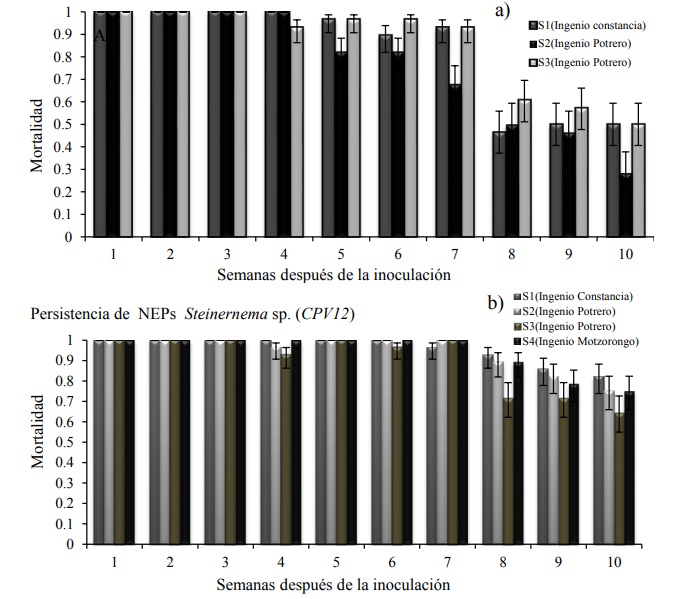
Figura 3 Proporciones de mortalidad a) heterorhabditis sp.; y b) steinernema sp. en larvas de G. mellonella. Las barras de error representan límites de confianza de 95%, transformada a partir de la escala logística. La persistencia de JI constituye el valor promedio de larvas muertas de G. mellonella por fecha de evaluación a 25 °C.
Discusión
Estos resultados muestran que los aislamientos de nematodos entomopatógenos CPVC13; CPVC12 (Steinernema) y CPVG13 (Heterorhabditis) pueden infectar ninfas de Aeneolamia albofasciata, siendo capaces de atravesar las barreras físicas del insecto, principalmente la masa espumosa de saliva que les sirve de protección. La capacidad de infección de los aislamientos de nematodos utilizados puede ser confirmada por la reproducción de los mismos en su hospedante, especialmente por el aislamiento CPVG13 de Heterorhabditis (Cuadro 2), así como por previos reportes de infección en otras especies de hemípteros bajo condiciones de laboratorio, invernadero y campo (Ferrer et al., 2004; Leite et al., 2005; Rosero- Guerrero, 2011; Morero et al., 2012). Se encontró una mayor infección ocasionada por Heterorhabditis comparada con Steinernema, lo cual podría ser atribuible a que los Heterorhaditidos tienen la capacidad de penetrar a través de la cutícula del hospedero con la ayuda de un pequeño diente que poseen (Bedding y Molyneux, 1982; Aguilera, 2001). La alta mortalidad de ninfas de A. albofasciata en el testigo probablemente haya sido ocasionada por patógenos traídos de campo, así como por la manipulación al momento de ser colectados.
Uno de los factores de mortalidad natural observados en esta investigación fue la presencia de nematodos mermitidos del género Hexamermis sp., fenómeno que ya ha sido reportado previamente en A. varia (Poinar y Linares, 1985) y Manhanarva fimbriolata (Bennett, 1984). Por esta razón, se considera importante que, en futuros experimentos, el material biológico proceda de colonias de insectos mantenidos en condiciones controladas, evitando con ello alta mortalidad durante el bioensayo y obtener resultados estadísticamente confiables.
En el experimento de persistencia de nematodos entomopatógenos, se observaron diferencias significativas entre tratamientos y cepas de nematodos, lo que significa que hubo una disminución de la población inicial de nematodos a través del tiempo. Tanto en Steinernema como en Heterorhabditis la persistencia de JI medida como la capacidad de matar insectos cebo a través del tiempo, mostró ligeras variaciones desde la cuarta semana, en el caso de Steinernema la mortalidad se mantuvo cercana al 100% hasta la séptima semana; sin embargo, en la octava semana, la proporción de mortalidad mostro variaciones 0.65 a 0.9 dependiendo del suelo donde se incubo (Figura 3B). En el caso de Heterorhabditis la persistencia de JI mostró variaciones más evidentes en las siguientes semanas. Al igual que Steinernema, a partir de la octava semana hubo una reducción en la persistencia de JI, pero en este caso la caída fue más drástica (0.25) (Figura 3B).
Heterorhabditis mostró problemas de persistencia en los diferentes tipos de suelo (Figura 3A), en suelo arcilloso, franco arcilloso y franco arenoso, con pH ácido a neutro (Cuadro 3) Heterorhabditis mostró una caída drástica en la curva de mortalidad, lo que significa que el suelo representa un factor importante para la persistencia de este nematodo entomopatógeno (Kung et al., 1990; Portillo-Aguilar et al., 1999).
De acuerdo con (Kaya, 1990; Kaya y Koppenhöfer, 1996; Epsky et al., 1998) la interacción entre microorganismos del suelo y nematodos entomopatógenos puede afectar la sobrevivencia de los JI, afectar el establecimiento y desarrollo del simbionte y afectar al insecto una vez invadido por el nematodo. Sin embargo, en este estudio se encontraron diferencias en la concentración de bacterias, hongos y actinomicetos presentes en el suelo, que pueden tener relación con la persistencia de los nematodos entomopatógenos. La interacción nematodo vs microbiota pudo afectar de manera diferente la cepa de nematodos, provocando mayor mortalidad en Heterorhabditis (CPVG13). Sin embargo, faltan estudios más profundos para corroborar el efecto del suelo-microbiota sobre la persistencia y actividad parasítica de los nematodos entomopatógenos.
Conclusiones
Los aislamientos de nematodos entomopatógenos del género Steinernema y Heterorhabditis tuvieron la capacidad de infectar a ninfas (salivazo) de Aeneolamia albofasciata, por su capacidad para atravesar la barrera de saliva espumosa que cubre a la ninfa. Además de mostrar efectividad biológica por su virulencia para desarrollarse y reproducirse en el hospedante. En especial, por el género Heterorabditis (CPVG13).
La persistencia de los aislamientos de Steinernema sp., como de Heterorhabditis sp., en cuanto a JI en ambos casos se mantuvo, en el caso de Steinernema a la septima semana la mortalidad casi fue al 100%, solo que a la octava semana hubo variaciones entre 0.65 a 0.9 dependiendo del tipo de suelo donde se mantuvo, en cambio Heterorhabditis la persistencia de JI fue semejante solo que destaco su caída repentina por la mortalidad que fue más pronunciada y esto se observó tanto en suelo arcilloso, franco arcilloso, y franco arenoso como en pH ácido a neutro.

