











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
En el suelo, la cantidad de P disponible para las plantas depende de la modificación del equilibrio dinámico que mantiene la disolución de los compuestos inorgánicos insolubles y de la descomposición de la materia orgánica (Navarro, 2003). Estos procesos los llevan a cabo microrganismos del suelo, a través de una serie de reacciones físicas-químicas y biológicas. Aproximadamente, 10% de la comunidad microbiana del suelo transforma activamente el P mediante los procesos de mineralización, solubilización, inmovilización y oxidación (Alexander, 1980; Coyne, 2000; Reyes, 2011).
Los microorganismos rizosféricos como los hongos micorrícicos arbusculares (HMA) y hongos solubilizadores de fósforo (HSF) son de particular importancia para las plantas, la fertilidad del suelo y los HSF promueven la disponibilidad del fósforo (Jeffries y Barea 2001), mientras que los HMA facilitan su transporte a la planta (Ferrol et al., 2002). La inoculación de hongos solubilizadores es una técnica prometedora, ya que se han reportado incrementos en el rendimiento de trigo (Whitelaw et al., 1997), cebolla (Vassilev et al., 1997) y soya (El-Azouni, 2008).
Adicionalmente, otros estudios muestran un mejor desarrollo y productividad de las plantas utilizando mezclas o consorcios de hongos solubilizadores de fósforo (Shin et al., 2006; Mittal et al., 2008). Estudios sobre HMA, han mostrado que estos hongos no solo actúan en el desarrollo y crecimiento de la planta, sino que contribuyen a la protección contra los patógenos (Bañuelos et al., 2014). Diversas investigaciones (Fernández et al., 2006; Mujica, 2012; Mujica y Batlle, 2013; Velázquez et al., 2017) han descrito que el sinergismo entre estos dos grupos de hongos (HSF y HMA), pueden repercutir en un mejor desarrollo de las plantas que cuando se inoculan solos.
El cultivo del jitomate en México es uno de los más importantes, durante 2015, el tomate fue el producto más importante en el valor de las exportaciones agropecuarias mexicanas, con una participación de 13%, al ubicarse en 1 665.8 millones de dólares y 1.43 millones de toneladas. El volumen exportado durante ese año fue equivalente al 53.3 por ciento de la producción nacional de esta hortaliza. La disponibilidad de nuevos tipos y variedades, nuevos métodos de cultivo y la creciente demanda de hortalizas han incentivado la producción mundial de tomate rojo.
El volumen cosechado a nivel mundial, el consumo total, así como el consumo promedio per cápita registran tendencia al alza durante la década reciente. China se mantiene como el principal productor y consumidor. Estados Unidos de América es el principal importador mundial, mientras que México es el principal proveedor externo (SAGARPA, 2018). Esta hortaliza demanda grandes cantidades de nitrógeno, fósforo y potasio; estos fertilizantes, al aplicarlos en dosis superiores a los requerimientos de los cultivos, conducen a un bajo aprovechamiento (Peña-Cabriales et al., 2001), contaminación de los suelos (Castellanos y Peña-Cabriales, 1990) y de los mantos acuíferos, así como la eutrofización de aguas superficiales (Nebel y Wright, 1996).
En este contexto, el uso de microorganismos debidamente seleccionados por su eficiencia y generados a partir de recursos locales, constituye una alternativa ecológicamente aceptable para reducir los insumos externos y mejorar la cantidad y calidad de los recursos internos. Este estudio surge por el interés de investigar a nivel de invernadero las interacciones de HSF y HMA nativos para el planteamiento de biofertilizantes; a partir, de consorcios fúngicos que promuevan la disponibilidad y transporte del fósforo insoluble hacia las plantas de jitomate.
Materiales y métodos
Las semillas de jitomate (Lycopersicon esculentum Mill.) previo a la germinación se desinfectaron con Cloralex® comercial al 50% (15 min). Las plántulas se trasladaron a macetas de 250 g con arena y perlita estéril 1:1 (v/v), a cada una se le agregó fosfato tricálcico como fuente insoluble de fósforo (0.125 g), el inoculo de HMA (10 g) y el inóculo de cada una de las cepas de HSF (1 x 108 UFC mL-1) tal y como lo propone Souchie et al. (2010). La aplicación de los inoculantes se realizó directamente al sustrato y sobre el sistema radicular de la planta.
Se estableció un diseño experimental de dos factores con cinco niveles, en total 10 tratamientos con tres repeticiones, esto es 30 unidades experimentales. Los tratamientos incluyeron: un consorcio de HMA, tres cepas de HSF (Aspergillus niger Tiegh, Penicillium brevicompactum Dierckx y Penicillium waksmanii K. W. Zaleski) y todas las posibles combinaciones incluyendo un testigo (sin hongos).
Las cepas de HSF empleadas: A. niger (An), P. brevicompactum (Pb) y P. waksmanii (Pw) pertenecen a la colección del Laboratorio de micromicetos del Instituto de Ecología, las cuales fueron aisladas de suelos de la región del centro del estado de Veracruz, México y se seleccionaron por su alta capacidad para solubilizar fosfato tricálcico. El consorcio de HMA pertenece a la colección del laboratorio de organismos benéficos de la Facultad de Ciencias Agrícolas de la Universidad Veracruzana. Las especies que conforman este consorcio fueron aisladas de suelos de la región de Xalapa y sus alrededores en el estado de Veracruz.
El inóculo HMA contenía 60 esporas g-1 de sustrato integrado con fragmentos de raíz con 79% de colonización, compuesto por las especies de Acaulospora morrowiae (Spain y N. C. Schenck), A. spinosa (C. Walker y Trappe), A. scrobiculata (Trappe), Gigaspora rosea (T. H. Nicolson y N. C. Schenck), Scutellospora pellucida (T. H. Nicolson y N. C. Schenck) C. Walker y F. E. Sanders, Glomus macrocarpum (Tul. y C. Tul.), Funneliformis mosseae (T. H. Nicolson y Gerd.) C. Walker y A. Schüßler, F. geosporum (T. H. Nicolson y Gerd.) C. Walker y A. Schüßler y Rhizophagus aggregatus (N. C. Schenck y G. S. Sm.) C. Walker.
El experimento se mantuvo durante cuatro meses (mayo-agosto de 2015) en un invernadero en el Instituto de Ecología, AC. El riego se realizó por capilaridad y se le adicionaron semanalmente 25 mL de solución nutritiva de Hewitt (Hewitt 1969) sin fósforo. Transcurrido ese tiempo, se procedió a cosechar las plantas, para medir la altura y longitud de raíz, porcentaje de colonización micorrícica y fósforo total de la parte área de las plantas.
Colonización micorrícica. El clareo y tinción de raíces, se realizó mediante la técnica de Phillips y Hayman (1970). Para ello, las raíces se lavaron con abundante agua, posteriormente se cortaron en pedazos pequeños y se colocaron en tubos con KOH al 10%, inmediatamente se sometieron a baño maría durante 15 min. Transcurrido ese tiempo se retiró el KOH y se lavaron con agua. Enseguida se les adicionó HCl al 10% y se mantuvieron en ese reactivo durante 10 min. Enseguida, se lavaron con agua y se les agregó azul de tripano al 0.05% y se dejaron a baño maría por 10 min.
Se eliminó el exceso de azul de tripano y se colocaron en tubos con ácido láctico hasta su observación. Para determinar el porcentaje de colonización de las raíces de las plantas, se utilizó la técnica descrita por Giovannetti y Mosse (1980), que consistió en colocar las raíces teñidas con azul de tripano en una caja Petri cuadriculada (1 cm x 1 cm) con ácido láctico y se observaron las estructuras fúngicas bajo el microscopio estereoscópico. El porcentaje de colonización micorrícica se calculó dividiendo el número total de interacciones de raíces micorrizadas, sobre el número total de intersecciones, multiplicando el resultado por 100.
Medición de fósforo total foliar. Con 0.25 g de material vegetal foliar seco, se llevó a calcinación en una mufla a 500 °C durante 2 h (Mckean, 1993). Las cenizas se disolvieron en agua destilada y la concentración de fósforo se determinó a través del método del azul de molibdato (Murphy y Riley 1962). Las muestras se midieron en un espectrofotómetro a 660 nm de absorbancia.
Variables de crecimiento de las plantas. La altura se midió a partir del cuello de la raíz hasta el meristemo apical y la longitud de la raíz desde el cuello de la raíz hasta el meristemo radicular.
Análisis estadísticos
Primeramente, se comprobó el supuesto de distribución normal y homogeneidad de varianza de los datos mediante pruebas de Kolmogorov-Smirnov y Bartlet. Para detectar diferencias en la micorrización entre los tratamientos, se utilizó un análisis de varianza de una vía y pruebas de medias de Tukey. Además, con el fin de detectar una interacción significativa entre el factor hongos solubilizadores de fósforo y el factor micorriza se realizaron análisis de varianza factorial. Cuando los datos de las interacciones fueron significativos, se realizaron pruebas de medias de Tukey. Todos los análisis se realizaron en el programa Statistica versión 10.0 para Windows.
Resultados y discusión
Los resultados de la colonización micorrícica en este estudio muestran una variación significativa entre los diferentes tratamientos evaluados (p= 0.000) (Figura 1). La colonización micorrícica en las plantas de jitomate inoculadas con los hongos micorrícicos y solubilizadores de fósforo estuvo en el rango de 48-66% de colonización, lo cual confirma la alta micotrofía de la planta, como se ha demostrado en otros estudios.
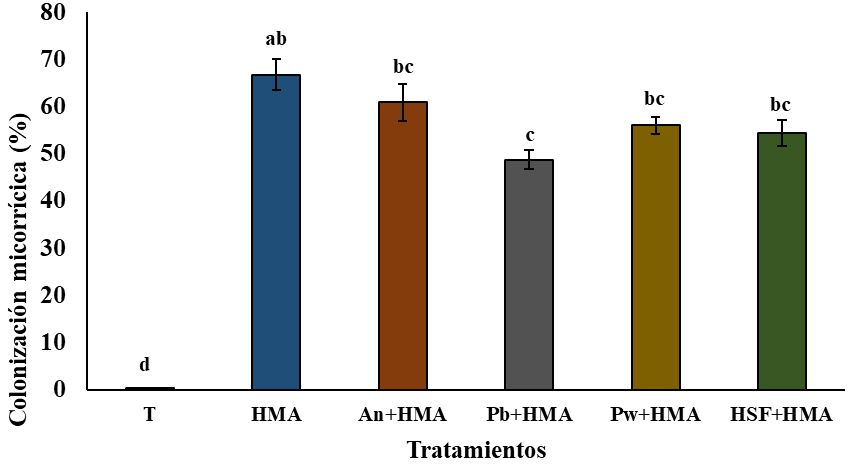
Figura 1 Porcentaje de colonización micorrícica de las plantas de jitomate después de cuatro meses de inoculados con los tratamientos. T= testigo; HMA= hongos micorrícicos arbusculares; An= Aspergillus niger; Pb= Penicillium brevicompactum; Pw= Penicillium waksmanii; y HSF= hongos solubilizadores de fósforo. n= 3; ±desviación estándar. Letras idénticas entre las columnas no existen diferencias significativas entre los tratamientos (Tukey LSD, p≤ 0.05).
En comparación con otras investigaciones en las que se evaluó la colonización micorrícica en plantas de jitomate inoculadas con HMA, en este trabajo se encontró mayor colonización a la reportada por Velázquez et al. (2005), quienes detectaron de 30-50% colonización micorrícica; Mujica y Fuentes (2012) de 40-47%; Mujica y Batlle (2013) 58% de colonización. La colonización micorrícica en este estudio fue analizada por la presencia de arbúsculos, vesículas y esporas; es importante señalar que se observó una alta esporulación dentro y fuera de las raíces (Figura 2).
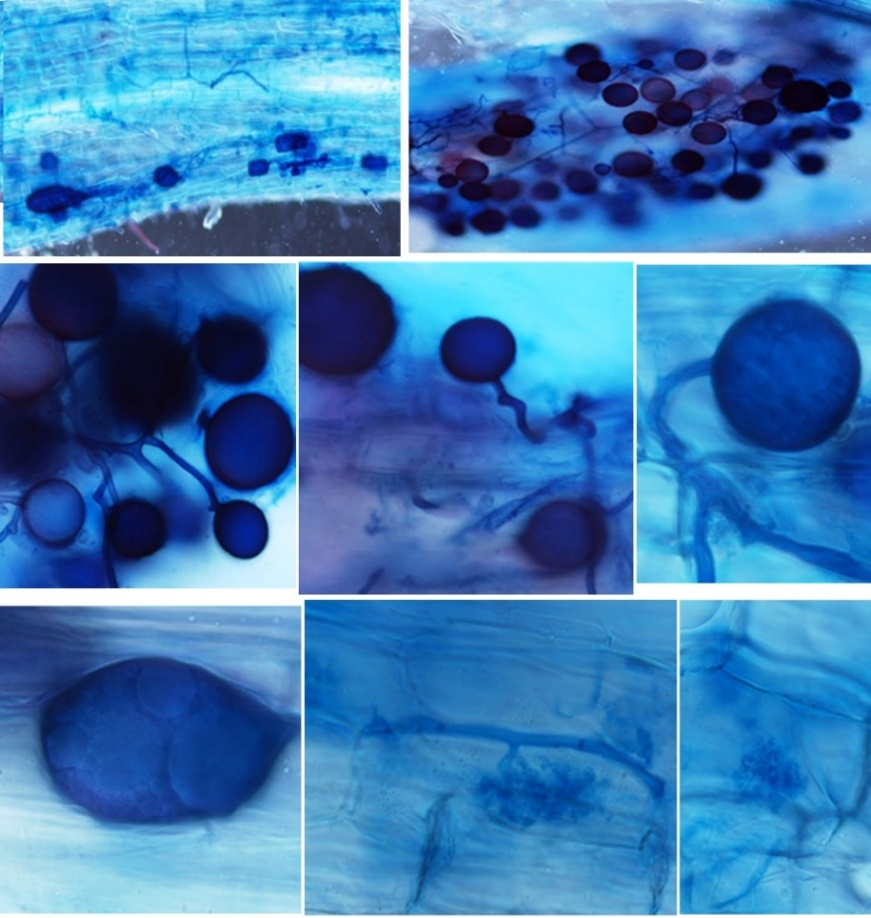
Figura 2 Raíces micorrizadas de jitomate a cuatro meses después de su inoculación. A) esporas germinadas dentro y fuera de las raíces; B) esporas; C) vesícula; y D) arbúsculos.
Las plantas de jitomate inoculadas con los tratamientos HMA + An fueron significativamente mayores que los tratamientos Pw (t= 0.006), Pb (t= 0.0002) y con HSF+HMA (t= 0.002) (Figura 1), sugiere un sinergismo entre el consorcio de HMA y la cepa solubilizadora de fósforo A. niger.
Existen estudios que confirman que la inoculación de hongos saprobios favorece la colonización de HMA en cultivos como chile con Claroideoglomus claroideum con Penicillium albidum y Penicillium frequentans (Castillo et al., 2013), en tomate con Glomus mosseae (Funneliformis mosseae) y A. niger (Velázquez et al., 2005), en trébol con Glomus clarum (Rhizophagus clarus) y Glomus geosporum (Funneliformis geosporum) con A. niger (Souchie et al., 2010); en bambú con Glomus spp. Aspergillus tubingensis (Babu y Reddy, 2011).
En este trabajo con jitomate, no se observó un incremento de la colonización micorrícica en presencia A. niger, P. brevicompactum y P. waksamnii de manera que se recomienda continuar evaluando la efectividad de otras cepas de HSF con el propósito de detectar alguna que favorezca una mayor colonización micorrícica.
Fósforo total foliar
Para el fósforo total foliar de las plantas de jitomate, se detectó una interacción significativa entre los factores HSF y HMA (p= 0.000). Todos los tratamientos presentaron una mayor concentración de fósforo total con respecto al testigo (t< 0.05). El tratamiento HSF fue significativamente mayor que el resto de los tratamientos (t< 0.05) (Figura 3). La concentración de fósforo total en la parte aérea en las plantas de jitomate osciló de 450 a 700 mg kg-1. Los tratamientos consorcios (HSF y HSF+ HMA) exhibieron los mayores incrementos (191%) de fósforo respecto al testigo.

Figura 3 Fósforo total foliar de la parte aérea de las plantas de jitomate después de cuatro meses de inoculadas con los tratamientos. T= testigo; HMA= hongos micorrícicos arbusculares; An= Aspergillus niger; Pb= Penicillium brevicompactum; Pw= Penicillium waksmanii; y HSF= hongos solubilizadores de fósforo. n=3; ±desviación estándar. Letras idénticas entre las columnas no existen diferencias significativas entre los tratamientos (Tukey LSD, p≤ 0.05).
Este incremento resulto superior a los reportados en otros estudios similares bajo condiciones de invernadero. Souchie et al. (2006) no detectaron incrementos del P foliar en Trifolium después de inocular G. mossae (Funneliformes mosseae) y Aspergillus niger. Por otra parte, Rodríguez-Cabral et al. (2012) encontraron 20% de incremento en esta variable en anacardo, también inoculado con estos hongos, Velázquez et al. (2005).
Reportan un incremento de 78% en el fósforo total foliar en tomate inoculado con G. mosseae (Funneliformes mosseae) y A. niger; Zhang et al. (2014) obtuvieron incrementos de 100% en una planta herbácea (Kosteletzkya virginica) inoculada con G. mosseae (F. mosseae) y Mortierella sp., Babu y Reddy (2011) detectaron 115% de incremento del fósforo foliar en plantas de bambú después de la co-inoculación de un consorcio de HMA y Aspergillus tubingensis, Zhang et al. (2014) por otro lado, detectaron un mayor incremento (300%) en fósforo foliar en plantas de ricino inoculadas con G. mosseae (F. mosseae) y Mortierella sp.
La combinación de la inoculación estos dos grupos de hongos (HMA y HSF) puede producir una mayor absorción de fósforo (Barea et al., 2008) e incrementar el desarrollo de las plantas principalmente por el efecto nutricional que esta combinación aporta, sin embargo existen otros mecanismos por los cuales los hongos pudieron afectar el desarrollo vegetal, tal es el caso de una mejora en la estructura del suelo (Khan, 1993), incremento en la humedad del suelo (Camargo-Ricalde, 2002), inducción de tolerancia al estrés ambiental (Cantrell y Linderman, 2001), mayor tasa fotosintética (Finlay, 1992), entre otros efectos distintos a los nutricionales reportados.
Variables de crecimiento de las plantas
Para la variable altura, se detectó una interacción significativa entre los factores HSF y HMA (p= 0.01) y un efecto significativo del factor HSF (p= 0.004). Sin embargo, el efecto del factor HMA no fue significativo (p= 0.35). La altura fue significativamente mayor en todos los tratamientos con hongos con respecto al testigo (Figura 4). El rango de valores promedios de las alturas (51-81cm) son superiores a los reportados por Velázquez et al., 2005 en plantas de jitomate (42.3 cm) con inoculaciones dobles de G mossae (Funneliformes mosseae) y una cepa de A. niger.
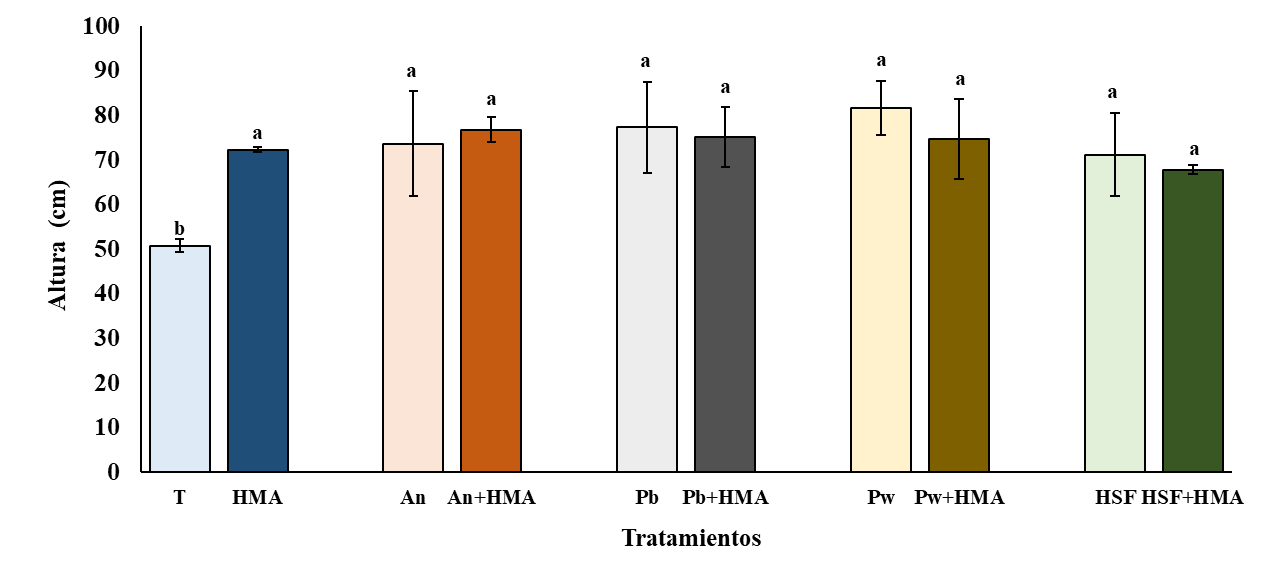
Figura 4 Altura de las plantas de jitomate después de cuatro meses de inoculadas con los tratamientos. T= testigo, HMA= hongos micorrícicos arbusculares, An= Aspergillus niger; Pb= Penicillium brevicompactum; Pw= Penicillium waksmanii; y HSF= hongos solubilizadores de fósforo. n= 3; ±, desviación estándar. Letras idénticas entre las columnas indican que no existen diferencias significativas entre los tratamientos (Tukey LSD, p≤ 0.05).
En otros estudios, también se ha observado que las inoculaciones dobles de HMA y HSF favorecen la altura de las plantas (Babu y Reddy, 2011; Castillo et al., 2013). Para la longitud de raíz de las plantas de jitomate, se detectó una interacción significativa entre los factores HSF y HMA (p= 0.03). No se observó un efecto significativo del factor HMA (p= 0.93); sin embargo, si lo hay con el factor HSF (p= 0.000). Las plantas con los tratamientos HMA y el testigo mostraron raíces significativamente más largas que con Pb +HMA, Pw +HMA y HSF (t< 0.05) (Figura 5).

Figura 5 Longitud de raíz de las plantas de jitomate después de 4 meses de inoculadas con los tratamientos. T= testigo; HMA= hongos micorrícicos arbusculares; An= Aspergillus niger; Pb= Penicillium brevicompactum; Pw= Penicillium waksmanii; y HSF= hongos solubilizadores de fósforo. n= 3; ±desviación estándar. Letras idénticas entre las columnas indican que no existen diferencias significativas entre los tratamientos (Tukey LSD, p≤ 0.05).
Los valores altos de longitud de raíces de las plantas testigos pudiera ser debido a las condiciones de poca disponibilidad de nutrientes en que se encontraban las plantas, las raíces se extendieron en busca de nutrientes (Barea et al., 2008). La longitud de raíz de las plantas inoculadas con el consorcio de HSF (mejor tratamiento en la disponibilidad de fósforo en la parte aérea) presentaron los valores menores, probablemente se debió a que la inoculación con el consorcio de HSF al proporcionar una mayor concentración de nutrientes a la planta, no fue necesario la elongación de la raíz ya que los nutrientes se encontraban de manera directa para las plantas (Barea et al., 2008).
Las cepas utilizadas son nativas de la región, el consorcio de HMA ya había sido probado y demostrado su eficiencia en otros cultivos (Ferrera et al., 2007; Sangabriel-Conde et al., 2010; Bañuelos et al., 2012; Zulueta- Rodríguez et al., 2013; Bañuelos et al., 2014). Por su parte, las cepas A. niger, P. brevicompactum y P. waksmanii también son nativas de la región centro de Veracruz, ya se había demostrado su capacidad como solubilizadoras de fosfato de calcio en condiciones in vitro (Perea, 2019).
Es importante, incidir en los productores sobre el uso de estos hongos nativos como una alternativa para la aplicación de los biofertilizantes, adoptando de esta manera nuevas estrategias que vayan en aumento en la concientización sobre el cuidado del ambiente, una reducción en los costos de producción, la conservación y recuperación de los suelos, así como mantener la biodiversidad (Fernández y Rodríguez, 2005).
Conclusiones
Las inoculaciones de los consorcios de HSF y HMA funcionaron de manera sinérgica, la relación HMA y HMA + An propició un alto porcentaje de micorrización, la interacción de HSF y HMA una mayor disponibilidad y transporte del fósforo en las plantas de jitomate y las alturas de las plantas se incrementó con todos tratamientos con hongos.

