










Serviços Personalizados
Journal
Artigo
 texto em
texto em  Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista mexicana de ciencias agrícolas
versão impressa ISSN 2007-0934
Rev. Mex. Cienc. Agríc vol.10 no.6 Texcoco Set. 2019 Epub 02-Out-2020
https://doi.org/10.29312/remexca.v10i6.1326
Articles
Antagonism of Trichoderma spp. vs fungi associated with wilting of chilli
1División de Procesos Naturales-Universidad Intercultural del Estado de Puebla. Calle principal a Lipuntahuaca s/n, Lipuntahuaca, Huehuetla, Puebla. CP. 73475. (andrahoy@gmail.com; hebert.jair@uiep.edu.mx).
2CONACYT-Instituto de Investigaciones Químico Biológicas-Universidad Michoacana de San Nicolás de Hidalgo. Av. Francisco J. Mújica s/n, Ciudad Universitaria, Morelia, Michoacán. CP. 58030.
3División de Estudios de Posgrado-Universidad Autónoma de Tamaulipas. Matamoros s/n, Zona Centro, Ciudad Victoria, Tamaulipas. CP. 87000. (eosorio@docentes.uat.edu.mx).
4Ingeniería en Biotecnología-Universidad Politécnica de Puebla. Tercer carril del ejido Serrano s/n, San Mateo Cuanalá, Juan C. Bonilla, Puebla. CP. 72640. (eduardo.molina@uppuebla.edu.mx).
5Universidad Politécnica Francisco I. Madero-Ingeniería en Agrotecnología. Tepatepec s/n, Francisco I. Madero, Hidalgo. CP. 42660. (nlanderova@conacyt.mx).
In Mexico, more than 100 varieties of chili (Capsicum annuum L.) are planted, it is a highly profitable crop and represents an economic activity of national importance. Currently, it faces serious phytosanitary problems due to the presence of diseases such as the wilting of chili, caused by a complex of soil pathogens. To control these diseases, toxic and residual fungicides are used, which pollute the environment and induce genetic resistance in phytopathogens. Therefore, it is necessary to look for control alternatives to solve this problem, therefore, the objective of this work was to evaluate the antagonistic effect of Trichoderma spp., against the causal agents of wilting in in vitro confrontations by means of dual cultures. The study was carried out in 2016, at the Intercultural University of the State of Puebla, Huehuetla, Puebla, Mexico. Three Trichoderma isolates from avocado tree rhizospheres, T. viride, T. harzianum and T. asperellum and three isolates of the phytopathogens P. capsici, F. oxysporum and R. solani obtained from the roots of plant plants were evaluated chili with wilting symptoms. The isolates were morphologically characterized for identification. The three antagonistic fungi presented a high percentage of inhibition, T. asperellum showed 88.25%, T. viride 87.22% and T. harzianum with 87.8%. Under the conditions of the study, T. asperellum, T. viride and T. harzianum, were efficient antagonistic microorganisms against the studied phytopathogens. It is suggested to carry out biocontrol evaluations with these isolates in the greenhouse and in the open field.
Keywords: Capsicun annum; Phytophthora capsici and Trichoderma spp
En México se siembran más de 100 variedades de chile (Capsicum annuum L.), es un cultivo altamente redituable y representa una actividad económica de importancia nacional. Actualmente, enfrenta graves problemas fitosanitarios por la presencia de enfermedades como la marchitez del chile, causada por un complejo de patógenos de suelo. Para el control de estas enfermedades, se utilizan fungicidas tóxicos y residuales, que contaminan el ambiente e inducen resistencia genética en los fitopatógenos. Por ello, es necesario buscar alternativas de control para solucionar este problema, por tanto, el objetivo de este trabajo fue evaluar el efecto antagónico de Trichoderma spp., contra los agentes causales de marchitez en confrontaciones in vitro mediante cultivos duales. El estudio se llevó acabo en 2016, en la Universidad Intercultural de Estado de Puebla, Huehuetla, Puebla, México. Se evaluaron tres aislamientos de Trichoderma provenientes de la rizósferas de árboles de aguacate, T. viride, T. harzianum y T. asperellum y tres aislamientos de los fitopatógenos P. capsici, F. oxysporum y R. solani obtenidos de las raíces de plantas de chile con síntomas de la marchitez. Los aislamientos se caracterizaron morfológicamente para su identificación. Los tres hongos antagonistas presentaron alto porcentaje de inhibición, T. asperellum mostró el 88.25%, T. viride 87.22% y T. harzianum con 87.8%. En las condiciones del estudio, T. asperellum, T. viride y T. harzianum, resultaron microorganismos antagónicos eficientes contra los fitopatógenos. Se sugiere realizar evaluaciones de biocontrol con estos aislamientos en invernadero y a campo abierto.
Palabras clave: Capsicun annum; Phytophthora capsici; Trichoderma spp
Introduction
The cultivation of chili (Capsicum annuum L.) in Mexico, faces serious phytosanitary problems that limit its production due to diseases such as wilting of chili, induced by a complex of soil phytopathogens, including Phytophthora capsici Leo (Hernández-Castillo et al., 2014), Rhizoctonia solani Kühn and Fusarium oxysporum (Vásquez et al., 2009). The main symptoms caused by this disease are the premature death of the plants, causing the obstruction and imbalance of the functioning of the vascular bundles, defoliation, color changes, foliage curl, damage to reproductive structures, advanced and irregular maturation, rot of the root, stem necrosis and occurrence of regrowths (Granke et al., 2012; Rivera et al., 2018).
To control the incidence and severity of these pathogens, highly toxic and residual fungicides are used that cause a negative environmental impact, contaminating the soil, air and aquifers, also poison the human being, create genetic resistance in the phytopathogens to the ingredients assets (Matar et al., 2009; Osorio et al., 2016; Hernández-Hernández et al., 2018) and increase production costs. Another consequence is the reduction of the planting area due to the infestation of the soil is the increase in the severity and incidence of the disease and reduction of the yields obtained by planted area (García, 2010).
The relevance of the genus Trichoderma lies in the efficiency it must fight diseases, the impact on root rotting has been reported in multiple studies, a problem that is found among diseases that cause significant economic losses in different agricultural crops. Although chemicals are still the main control tool for these diseases, biological agents are an effective way to provide faster and safer control (Verma et al., 2007).
The biological control mechanisms of Trichoderma spp. species have been investigated for more than 70 years (Martínez et al., 2013), its antagonistic capacity is widely documented as biocontrol agents of various pathogens that affect crops of agricultural and economic importance (Infante et al., 2009; Mayo et al., 2015). Trichoderma strains produce extracellular enzymes (Osorio-Hernández et al., 2016), compete with pathogenic fungi for space and nutrients by completely reducing or stopping the development of phytopathogens (Sánchez-García et al., 2017), promoting growth of plants and induce systemic resistance, mycoparasitism and antibiosis through direct restrictive action to phytopathogenic fungi.
These mechanisms are favored by the ability of Trichoderma isolates to colonize the rhizosphere of plants. (Leandro et al., 2007; Woo and Lorito, 2007; Guedez et al., 2012). The different species of Trichoderma are an alternative for biocontrol against various soil phytopathogens (Singh et al., 2011) including P. capsici, F. oxysporum and R. solani, in addition to stimulating the increase in root biomass and area foliar (Ezziyyani et al., 2005; Ezziyyani et al., 2007). Therefore, the objective of this research was to determine the antagonistic effect of Trichoderma spp., as a biocontrol method for the complex of phytopathogens that cause wilting of chili in in vitro evaluations using dual cultures.
Materials and methods
Study location
This work was carried out in the Biology Laboratory of the Intercultural University of the State of Puebla, Huehuetla, Puebla, Mexico. Its geographical coordinates are the parallels 20º 01’ 48” and 20º 09’ 12” of north latitude and the meridians 97º 35’ 00” and 97º 40’ 24” of west longitude.
Fungal material
Three species of Trichoderma were isolated from soil in potato dextrose-agar medium (PDA). 1 g of wet soil was dispersed in boxes with PDA medium and incubated at 25 ±2 °C in the dark for two days, then removed from the incubator and kept under a white light lamp for two days to induce sporderma sporulation sp., from the development of Trichoderma, a portion was taken to transfer to PDA culture medium and incubate at 25 ±2 °C, this was done until obtaining axenic and pure growths, free of other fungi or bacteria. After the last sporulation in this medium, they were preserved in sterile water and mineral oil.
The pathogens Rhizoctonia sp., Fusarium sp. and Phytophthora capsici were isolated from chili roots, the roots were washed with running water; subsequently, the tissue portion was deflated by immersion for 1 min in 1.5% solution of sodium hypochlorite and rinsed with sterile distilled water, the roots were drained on sterile sanitas, finally 1 cm fragments were seeded in PDA medium for isolation and purification of Rhizoctonia sp. and Fusarium sp.
In the case of the Phytophthora capsici oomycete, 1.5 cm long chili roots were seeded in agar-agar® medium with V8® juice (8 vegetable juice) and antibiotics were added (Pimaricin 10 µg L-1, Ampicillin 292 µg L-l, Rifampicin 10 µg L-1, Pentachloronitrobenzene 0.1 g L-1 and Himexazol 0.25 µg L-1), subsequently incubated at 28 ±2°C for three days (Andrade et al., 2012), previously purified pathogens from the chili root was kept under conservation in sterile water for the purpose of preservation (Molina-Gayosso et al., 2016). Prior to dual confrontation, microorganisms were activated in PDA culture medium plus lactic acid (Figure 1).
Morphological characterization
To study the morphological characteristics, both pathogen and Trichoderma isolates were grown in PDA medium and incubated at 24 ±2 °C for 12 h under white light (Rivera-Jiménez et al., 2018). In the identification of the genus Fusarium sp. the description of Burgess et al. (1994) and the identification of the species was made using the keys of Seifert (1996). Rhizoctonia species was identified based on Sneh et al. (1991) and that of the oomycete Phytophthora sp. It was characterized by the keys of Erwin and Ribeiro (1996); Gallegly and Hong (2008).
Finally, in the morphological characterization of the isolates of Trichoderma spp. the taxonomic keys of Barnett and Hunter (1972) were used. For the Trichoderma isolates, monoconidial cultures were used in combination with the wet chamber technique (Harris, 1986). A 5 mm diameter PDA disc was inoculated with the Trichoderma spore and placed between a slide and coverslip and this preparation on a sterile rod triangle inside a petri dish and incubated at 25 ±2 °C for 24 h.
The macro and microscopic characteristics to be considered were colony texture, presence of mycelium, concentric rings, colony staining on the reverse, shape and size of conidia and phyloid (Chaverri et al., 2015; Jang et al., 2018; Nawaz et al., 2018; Du Plessis et al., 2018), the structures of the Trichoderma spp. at 40X in an optical microscope (Zeiss Axiosiop plus).
Growth rate
In determining the growth rate of mycelium in antagonists and pathogens, grown in PDA, measurements were made every eight hours.
Trichoderma antagonistic activity on pathogens
The dual culture technique was used to determine the antagonistic activity of the different Trichoderma isolates on Rhizoctonia sp., Fusarium sp. and P. capsici (Sonnenbichler et al., 1983). A completely randomized design was used (with two factors: Trichoderma and pathogens), with eight repetitions.
The tests were carried out evaluating dual cultures of Trichoderma spp., Contrasted with three phytopathogens, three isolates of Trichoderma against Rhizoctonia sp., Fusarium sp. and P. capsici. In the test of dual cultures, petri dishes with antibiotic-free PDA were used, a 5 mm diameter disc with active mycelium of the phytopathogen (previously grown in PDA for five days) was placed on one end and an equidistant end was placed Mycelium disc (previously grown in PDA for five days) of 5 mm in diameter with Trichoderma spp., the boxes were incubated at 28 ± 2 °C and observed every 8 h to record the number of days at the first contact between the antagonist and the phytopathogen, the growth of both colonies (cm) was measured and the percentage of inhibition of radial mycelial growth was evaluated, based on the formula proposed by Ezziyyani et al. (2007), calculating as follows:
Where: PICR= percentage of radial growth inhibition; R1= radial growth (mm) of the pathogen without Trichoderma spp., R2= radial growth (mm) of the pathogen with Trichoderma spp.
Trichoderma mycoparasitism on pathogens
To know the presence of parasitism of the Trichoderma strains towards Rizoctonia sp., Fusarium sp. and P. capsici, the Riddel microculture technique described by Paul (1999) was used. This technique consists of placing a V-shaped glass rod inside a petri dish and on top of it, a sterile slide, on which a 10 mm diameter PDA disc was placed. The arrangement of each fungus was made in two cardinal points, after placing the mycelium of both fungi on the PDA, with a difference of 24 h, the pathogens were first placed with the help of a sterile needle, which have a slow growth compared with Trichoderma and a sterile coverslip was placed, it was incubated at 25 ±2 °C.
When the agar disk was covered with the growth of both fungi, the slide was removed from the petri dish to prepare the smears. The coverslip was placed on another clean slide and provided with a drop of dye known as cotton blue. On the other hand, the agar disc was removed from the original slide, a drop of the dye and a coverslip were placed. Once the excess dye was removed, the preparations were sealed and evaluated microscopically, that is, 10 samples were observed to determine whether or not there was parasitism by the Trichoderma sp. strains, 40X was observed in an optical microscope (Zeiss Axiosiop plus).
Statistic analysis
The PICR data, expressed as a percentage, were transformed with the angular arcsine √x+1 (Steel et al., 1986) and in the mycelial growth in the biocontrol test, a completely randomized design was used (three strains of Trichoderma sp and three phytopathogens) with eight repetitions. The data obtained for each of the trials were subjected to an analysis of variance and a comparison test of Tukey means (p≤ 0.05). These data were analyzed with the statistical package SAS version 9.0 for Windows (SAS, 2002).
Results and discussion
Morphological characterization
The pathogens causing the wilting of chili corresponded with what was reported by Rivera-Jimenez et al. (2018). Similarity was found with F. oxysporum, the growing colonies in PDA had abundant aerial mycelium and pale pink to deep purple to magenta with a cottony texture. The macro-conidia presented one to five septa, the conidia measured 23-54 × 3-4.5 μm, respectively. In the identification of Rhizoctonia solani, it corresponded with that reported by Sneh et al. (1991) and Lozano et al. (2015), R. solani was characterized by forming aerial mycelium, brown, in PDA culture medium formed mycelium hyaline.
The hyphae generally showed a right angle and after this a septum formed, the diameters of the hyphae presented 4.1 to 8.9 μm, which corresponds to Barnett and Hunter (1998). The isolation of Phytophthora capsici was characterized by forming ovoid, elongated, ellipsoid sporangia with one or two papillae, the size of the sporangia was 35.8 to 64.7 x 19.6 to 25.2 μm., finally the apical thickness was 1.85 to 4.3 μm (Erwin and Ribeiro, 1996; Gallegly and Hong, 2008).
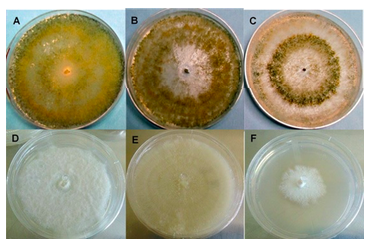
According to the taxonomic identification criteria of Barnett and Hunter (1972), Trichoderma species were compared with taxonomic keys and identified by their morphological characteristics. The species of T. harzianum presented light-to-dark green colonies with a dusty-cottony texture, with mycelium, conidophore and green-looking conidia (Figure 2, Table 1) show various branches in pyramidal form, in some cases the formation of two to three lateral branches and presents aerial mycelium. In T. asperellum and T. viride, an absent coloration can be seen on the back of the petri dish.
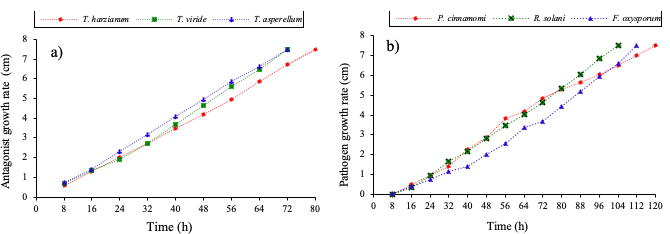
Figure 2 Growth rate, A) T. harzianum, T. viride and T. asperellum; and B) P. capsici, R. solani and F. oxysporum.
Table 1 Morphological characterization of Trichoderma spp.
| Trichoderma spp. | Colony color | Colony texture | Aerial mycelium | Rings | Color on the back | Shape of conidia | Size conid (μm) | Phyloid form | Size phialides (μm) |
| T. harzianum | Dark green to grayish color | Cottony | Abundant | No presence | Colorless | Subglobose | 3.8-4x 3.1-3.7 | Globose | 6.3-15.6x 2.7-3.4 |
| T. asperrellum | Light green to dark green | Dusty | No presence | 3 to 4 rings | Colorless | Ovoid to elisoipse | 4-4.4x 3-3.5 | Slightly globose | 2.3-2.8x 10.2-11.8 |
| T. viride | Dark green | Cottony | Abundant | No presence | Colorless | Ellipsoidal globose | 4-4.8x 3.5-4 | Lageniformes | 8-14 x2.4-3 |
The spores and phyloids, characteristic structures correspond to Trichoderma species, presented forms and sizes typical of the species of T. harzianum, T. asperellum and T. viride corresponding to what was reported by different authors (Barnett et al., 1972; de Hoog, 2000; Romero-Arenas et al., 2009; Chaverri et al., 2015; Jaklitsch et al., 2015).
Growth rate
The evaluation of the initial growth rate of the antagonists and pathogens turned out to be completely related at the beginning of mycelial growth at 8 h; however, the three species of Trichoderma have the ability to antagonize the completion of the filling of the petri dish in 80 h. These results indicate the importance of knowing the growth rate of pathogens, as well as the antagonists of T. harzianum, T. viride and T. asperellum.
The isolates of Trichoderma spp. showed a faster growth than the pathogens, this behavior is promising in the control of pathogens of the root, also allowed this trial allowed to consider it for the PICR tests and give Fusarium oxysporum, Rhizoctonia solani and Phytophthora capsici two days advantage.
Antagonistic activity of Trichoderma on pathogens
The antagonistic activity of T. viride against P. capsici showed a significant difference (p≤ 0.05), because T. viride presented 0.516 mm in comparison to P. capsici, which obtained 0.241 mm of growth in a petri dish with culture medium, is say T. viride is suitable for use as a biological control agent, as it is effective against P. capsici, since hyperparasitism was also observed by the pathogen (Figure 3). The foregoing is consistent with that reported by Bouziane et al. (2016), when evaluating T. viride against P. infestans in in vitro tests, they found that the beneficial fungus showed inhibitory capacity between 58 and 68%, similar results were presented when evaluating in plants.
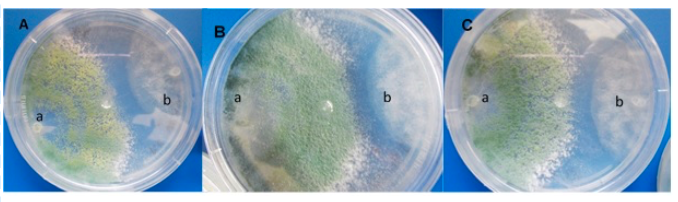
Figure 3 Dual growth, in vitro confrontations with Trichoderma against pathogens, A) Trichoderma viride vs. P. capsici; B) Trichoderma asperellum vs Fusarium oxysporum; and C) Trichoderma harzianum vs Rhizoctonia solani.
Similarly, Zegeye et al. (2011), found that T. viride showed a complete inhibition in the radial growth of P. infestans in in vitro tests. In addition, they mentioned that the foliar application of T. viride has a good potential to control P. infestans in greenhouse conditions. On the other hand, the evaluation of dual antagonism of T. viride against F. oxysporum showed growth of 0.656 mm of the antagonist and 0.221 mm by the pathogen, this evidences how T. viride is suitable for use in the control of F. oxysporum.
In the dual confrontation T. viride against R. solani obtained a growth of the 0.746 mm antagonist and the 0.112 mm pathogen, the results of growth by the antagonist were higher compared to the pathogen (Figure 3). In both cases similar results have been found where the two fungi are mycoparasited or destroyed by T. viride (Figure 3) since it has an anti-fungal effect with this pathogen (Jhon et al., 2010; Perveen, 2012; Sánchez-García et al., 2017).
It is important to mention that in the first 8 h the growth of the antagonists showed an accelerated growth that facilitates the encounter with the pathogen in less time, indicative that the antagonist grew or covered more surface in culture in vitro.
Similar results mentioned several authors where they demonstrated that Trichoderma spp. and T. viride are used for the biological control of R. solani (Sánchez-García et al., 2017). An antifungal effect was observed, in which it inhibits growth and causes lysis of this pathogen. In the confrontations of T. viride against the causative agents of the wilting of chili. The evaluation of the antagonism of T. harzianum against P. capsici, F. oxysporum and R. solani (Figure 4) showed greater growth by the antagonist compared to the pathogen, denoting hyperparasitism by completely invading the pathogen, a similar result was obtained by Osorio-Hernández et al. (2011) in which he observed that Tricoderma spp., is an effective antagonist for the control of P. capsici, inhibiting this pathogen up to 48%.
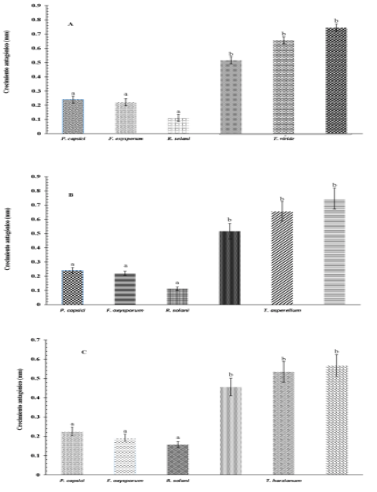
Figure 4 Mycelial growth (mm) in the biocontrol test of A) T. viride vs P. capsici, F. oxysporum and R. solani; B) T. asperellum. vs P. capsici, F. oxysporum and R. solani; and C) T. harzianum vs P. capsici, F. oxysporum and R. solani.
Also, in the confrontation of T. viride against F. oxysporum and R. solani, the same tendency was found. A significant value (p≤ 0.05) was found in the susceptibility assessment based on the PICR of the three pathogens responsible for the wilting of chili; however, the highest susceptibility was presented by Fusarium oxysporum with 92.68%, followed by Rizoctonia solani and P. capsici (Table 2), results that prove the effectiveness of the antagonists (Osorio et al., 2016) against pathogens that affect the root of the chilli from seedlings and plants in production (Romero et al., 2017).
Table 2 Evaluation of the susceptibility of pathogens to Trichoderma and percent inhibition of Trichoderma sp. against the pathogens causing the wilting of chili.
| Pathogens | Susceptibility of pathogens to Trichoderma PICR (%) | Trichoderma | Antagonistic activity Trichoderma PICR (%) |
| Fusarium oxysporum | 92.68 a | T. asperellum | 88.25 a |
| Rhizoctonia solani | 87.86 b | T. viride | 87.22 a |
| Phytophthora capsici | 83.33 c | T. harzianum | 87.8 a |
a, b, c= denotes the average treatments that are significantly the same or different according to the Tukey test in p< 0.05. PICR= percentage of radial growth inhibition.
In the evaluation of the percentage of radial growth inhibition (PICR) of the pathogens causing the wilting of the chili, there were no significant differences between the antagonists T. asperellum, T. viride and T. harzianum; therefore, it is suggested to use any of the three Trichoderma as a biological control agent; that is, they are effective in inhibiting more than 85% of the pathogens causing the wilting of chili (Table 2), the response of inhibition of the growth of phytopathogens is due to the synthesis of secondary metabolites and to the different mechanisms of action of mycoparasitism (Harman, 2006; Infante, 2009).
The similar inhibition behavior of the three treatments are favorable results found by those who report that above 80% inhibition is acceptable in the biocontrol of pathogens that are transmitted by the soil and by the efficacy of fighting root diseases (Hermosa et al., 2012; Sabbagh et al., 2017).
The results of mycoparasitism of the antagonists frequently had a more favorable growth, it should be mentioned that the first 8 h were significant (p≤ 0.05) showing that the antagonist had an accelerated growth that favors that the encounter can be had in a short time. Likewise, it was observed that not only did it present the inhibition of the growth of these pathogens, but it also presented a total invasion (hyperparasitism) of the antagonist towards the pathogen even sporulating on the pathogen. In a study with selections of 25 Trichoderma strains including T. harzianum, of which they presented inhibition in F. oxysporum on mycelium growth in 33 and 35%.
Similarly, Sinuco et al. (2017) indicate that volatile organic compounds of T. viride affected the growth halos of Fusarium spp. The growth of T. asperellum showed significant differences (p≤ 0.05) when confronted with the different pathogens causing the wilting of chili caused by the pathogens of P. capsici, F. oxysporum and R. solani (p≤ 0.001) since at to analyze the data, it was observed that the growth rate of the pathogen was reduced by almost half when confronted with T. asperellum.
In addition, this antagonist overgrown up to 100% on the pathogens, covering the box completely and thus causing hyperparasitism, thus showing a very efficient antagonistic ability to each of the pathogens.
These results coincide with other authors who mentioned that Trichoderma ssp. overgrowth Phytophthora causing hyperparasitism (Bae et al., 2016). Similarly, Trichoderma spp. it develops against R. solani, causing the inhibition of in vitro growth of R. solani from 58 to 86%. Trichoderma spp., has the ability to induce the expression of genes related to the defense of plants and produce a higher level of ergosterol, indicating its ability to grow at a higher rate in the soil, this explains its positive effects on the growth and defense of the plant in the presence of the pathogen (Mayo et al., 2015).
Trichoderma isolates confronted with P. capsici exerted a highly significant antagonistic effect (p≤ 0.001), causing inhibition in oomycete growth under dual conditions. T. harzianum, T. viride and T. asperellum, also had a similar behavior against F. Oxysporum and R. solani (Figure 4). The antagonists continued to grow up to six days later, showing hyperparasitism by invading or growing completely on the pathogen. In this regard, Segarra et al. (2013), mentioned that T. asperellum strain T34, is able to reduce P. capsici up to 71%, when applied to pepper plants at different stages of growth.
Mycoparasitism
In vitro tests the inhibition of the three species of Trichoderma was evaluated, the results showed the efficacy in competition for space and nutrients, in addition to appreciating the parasitic ability against Rhizoctonia solani, Fusarium oxysporum and P. capsici. In addition, was observed by light microscopy and corroborate in microcultures and double-confrontation cultures at the crossing point between the native isolates of Trichoderma spp. and pathogens causing the wilting of chili the different types of mycoparasitism (Figure 5).
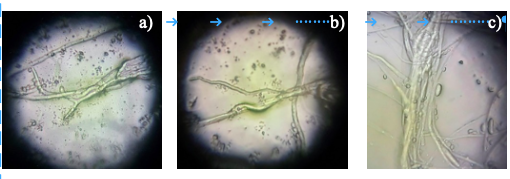
Figure 5 Mycoparasitism of Trichoderma sp. A) T. harzianum surrounding the mycelium of Rhizoctonia solani; B) T. viride and T. asperellum surrounding the mycelium of P. capsici; and C) massive winding of T. asperellum covering Fusarium oxysporum.
In some early investigations, myco-parasitism, antibiosis has been observed, in addition to Trichoderma having a growth behavior parallel to the pathogen until it is wound avoiding the different survival mechanisms of the pathogen (Chet et al., 1981; Harman, 2006; Infante, 2009).
Conclusions
T. asperellum, T. viride and T. harzianum are efficient antagonists of the phytopathogens that cause the wilting of chili. The tests with these antagonistic microorganisms have demonstrated the existence of biological alternatives for the control of phytopathogens of chili root in laboratory conditions, therefore, it is suggested to carry out biocontrol evaluations with these isolates and cultivation of chili in the greenhouse and in the open field. This will help to know the behavior in competitive conditions with a greater diversity of microorganisms and environmental factors different from those studied.
Literatura citada
Andrade, H. P.; De León, C.; Espíndola, B. M. C. , Alvarado, R.D.; López, J. A. y García, E. R. 2012. Selección de porta-injertos de aguacate para tolerancia-resistencia a Phytophthora cinamomo Rands. Usando temperaturas controladas. Spanish J. Rural Development. 3(4):23-30. [ Links ]
Bae, S. J.; Mohanta, T. K.; Chung, J. Y.; Ryu, M.; Park, G.; Shim, S.; Hong, S. B.; Seo, H.; Bae, D. W.; Bae, I., Kim, J. J. and Bae, H. 2016. Trichoderma metabolites as biological control agents against Phytophthora pathogens. Biological Control. 92:128-138. Doi:10.1016/ j.biocontrol.2015.10.005. [ Links ]
Barnett, H. and Hunter, B. 1972. Illustrated genera of imperfect fungi. EE. UU. Burgess Publ. Co., 241p. Doi:10.2307/3757954. [ Links ]
Barnett, H. L. and Hunter, B. B. 1998. Illustrated genera of imperfect fungi. 4a. Ed. American Phytophatology Society, MN. 217 p. [ Links ]
Bouziane, Z.; Dehimet, L. and Kacem, C. N. 2016. Inhibitory activity of Trichoderma viride against Phytophthora infestans that affects the Spunta potato (Solanum tuberosum L.) variety. Afr. J. Microbiol. Res. 29(10):1121-1127. [ Links ]
Burgess, L. W.; Summerell, B. A.; Bullock, S.; Gott, K. P. and Backhouse, D. 1994. Laboratory manual for Fusarium research. 3rd (Ed.). University of Sydney, Sydney, Australia. 132 p. [ Links ]
Chaverri, P.; Branco, R. F.; Jaklitsch, W.; Gazis, R.; Degenkolb, T. and Samuels, G. J. 2015. Systematics of the Trichoderma harzianum species complex and the re-identification of commercial biocontrol strains. Mycologia. 107(3):558-590. [ Links ]
Chet, I.; Harman, G. E. and Bake, R. 1981. Trichoderma hamatum: Its Hyphal Interactions with Rhizoctonia solani and Pythium spp. Microbial Ecol. 7(1):29-38. [ Links ]
de Hoog, G. S.; Guarro, J.; Gene, J. and Figueras, M. J. 2000. Atlas of clinical fungi, 2nd edition. entraalbureau voor Schimmelcultures, Utrecht. Mycological research. 106(4):507-511. Doi:10.1017/S0953756202226167. [ Links ]
Du Plessis, I. L.; Druzhinina, I. S.; Atanasova, L.; Yarden, O. and Jacobs, K. 2018. The diversity of Trichoderma species from soil in South Africa, with five new additions. Mycologia. 110(3):559-583. [ Links ]
Erwin, D. C. and Ribeiro, O. K. 1996. Phytophthora diseases worldwide. Department of Plant Pathology, University of California, Riverside, USA. American Phytopathol. Society. 562 p. [ Links ]
Ezziyyani, M.; Requena M. E.; Egea-Gilabert C. and Candela M. E. 2007. Biological Control of Phytophthora Root Rot of Pepper Using Trichoderma harzianum and Streptomyces rochei in Combination. Journal of Phytopathology. 155(6):342-349. Doi:10.1111/j.1439-0434.2007.01237.x. [ Links ]
Ezziyyani, M.; Requena, M. A. y Candela, M. A. 2005. Producción de proteínas-PR en la inducción de resistencia a P. capsici en plantas de pimiento. Anales de Biología. 27:143-154. [ Links ]
Gallegly, M. E. and Hong, M. C. 2008. Phytophthora: identifying species by morphology and DNA fingerprints. American Phytopathol. Society. St. Paul, M. N. 158 p. [ Links ]
García, E. R. 2010. Agroecología y enfermedades de la raíz en cultivos agrícolas. Colegio de Postgraduados, estado de México, México. 130 p. [ Links ]
Granke, L. L.; Quezada, O. L.; Lamour, K. and Hausbeck, M. K. 2012. Advances in research on Phytophthora capsici on vegetable crops in the United States. Plant disease. 11(96):1588-1600. [ Links ]
Guédez, C.; Cañizalez, L. M.; Castillo, C. y Olivar, R. 2012. Evaluación in vitro de aislamientos de Trichoderma harzianum para el control de Rhizoctonia solani, Sclerotium rolfsii y Fusarium oxysporum en plantas de tomate. Rev. de la Sociedad Venezolana de Microbiología. 3(32):44-49. [ Links ]
Harman, G. E. 2006. Overview of Mechanisms and uses of Trichoderma spp. Phytopathology. 96(2):190-194. Doi: 10.1094/PHYTO-96-0190. [ Links ]
Harris, I. J. 1986. Modified method for fungal slide culture. J. Clinical Microbiol. 24(3):460-461. [ Links ]
Hermosa, R.; Viterbo, A.; Chet, I. and Monte, E. 2012. Plant-beneficial effects of Trichoderma and of its genes. Microbiology. 158(1):17-25. Doi: 10.1099/mic.0.052274-0. [ Links ]
Hernández, C. F. D.; Lira, S. R. H.; Gallegos, M. G.; Hernández, S. M. and Solis, G. S. 2014. Biocontrol of pepper wilt with three Bacillus species and its effect on growth and yield. Phyton. 83(1):49-55. [ Links ]
Hernández, H. E. J.; Hernández, R. I.; Almaraz, S. J. J.; López, L. A.; Torres, A. M. y Morales, F. F. J. 2018. Caracterización in vitro de rizobacterias y su antagonismo con hongos causantes del damping off en chile. Rev. Mex. Cienc. Agríc. 9(3):525-537. [ Links ]
Infante, D.; Martínez, B.; González, N. y Reyes, Y. 2009. Mecanismos de acción de Trichoderma frente a hongos fitopatógenos. Revista de Protección Vegetal. 1(24):14-21. [ Links ]
Jaklitsch, W.M. and Voglmayr, H. 2015. Biodiversity of Trichoderma (Hypocreaceae) in Southern Europe and Macaronesia. Studies in Mycology. 80:1-87. Doi.org/10.1016/j.simyco. 2014.11.001. [ Links ]
Jang, S.; Kwon, S. L.; Lee, H.; Jang, Y.; Park, M. S.; Lim, Y. W.; Kim C. and Kim, J. J. 2018. New report of three unrecorded species in Trichoderma harzianum species complex in Korea. Micology. 46(3):177-184. [ Links ]
Jhon, R. P.; Tyagi, R. D.; Prévost, D.; Brar, S. K.; Pouleur, S. and Surampalli, R. Y. 2010. Mycoparasitic Trichoderma viride as a biocontrol agent against Fusarium oxysporum f. sp. adzuki and Pythium arrhenomanes and as a growth promoter of soybean. Crop Protection. 12(29):1452-1459. [ Links ]
Leandro, L. F. S.; Guzman, T.; Ferguson, L. M.; Fernandez G. E. and Louws F. J. 2007. Population dynamics of Trichoderma in fumigated and compostamended soil and on strawberry roots. Appl. Soil Ecol. 1(35):237-246. [ Links ]
Lozano, A. N.; Guzmán, P. R.; Zavaleta, M. E.; Aguilar, R. H.; Ayala, V. V. 2015. Etiology and evaluation of control alternatives for wilt in chile de árbol (Capsicum annuum L.) in La Vega, Metzitlán, Hidalgo, México. 33(1):31-53. [ Links ]
Martínez, B.; Infante, D. y Reyes, R. Y. 2013. Trichoderma spp. y su función en el control de plagas en los cultivos. Revista de Protección Vegetal. 1(28):1-11. [ Links ]
Matar, S. M.; El-Kazzaz, S. A.; Wagih, E. E.; El-Diwan, A. I.; Moustafa, H. E.; Abo-Zaid, G. A.; Abd-Elsalam, H. E. and Hafez, E. E. 2009. Antagonistic and inhibitory effect of Bacillus subtilis against certain plant pathogenic fungi, I. Biotechnology. 8(1):53-61. [ Links ]
Mayo, S.; Gutiérrez S.; Malmierca M. G.; Lorenzana A.; Campelo M. P.; Hermosa R. and Casquero P. A. 2015. Influence of Rhizoctonia solani and Trichoderma spp. In growth of bean (Phaseolus vulgaris L.) and in the induction of plant defense-related genes. Frontiers Plant Science. 6(685):1-11. Doi: 10.3389/fpls.2015.00685. [ Links ]
Molina, G. E.; Andrade, H. P.; García, E. R. y Sosa, H. C. M. 2016. Capacidad de sobrevivencia de tres especies de Phytophthora y dos de Pythium preservados en dos sustratos a corto y largo plazo. Rev. Mex. Cienc. Agríc. 7(7):1759-1764. [ Links ]
Nawaz, K.; Shahid, A. A.; Bengyell, L.; Subhani, M. N.; Alia, M.; Anwar, W.; Iftikhar, S. and Ali, S.W. 2018. Diversity of Trichoderma species in chili rhizosphere that promote vigor and antagonism against virulent Phytophthora capsici. Scientia Horticulturae. 239:242-252. Doi.org/10.1016/j.scienta.2018.05.048. [ Links ]
Osorio, H. E.; Hernández, C. F. D.; Gallegos, M. G.; Rodríguez, H. R. and Castillo, R. F. 2011. In-vitro behavior of Trichoderma spp. Against Phytophthora capsici Leonian. Afr. J. Agric. Res. 19(6):4594-4600. [ Links ]
Osorio, H. E.; Hernández, F. D. C.; Rodríguez, R. H.; Varela, S. D. F.; Estrada, B. D. y López, J. A. S. 2016. Actividad antagónica de Trichoderma spp. sobre Rhizoctonia solani in vitro. Investigación y Ciencia. 67(24):5-11. [ Links ]
Osorio, H. E.; Hernández, M. J.; Conde, M. V.; Michel, A. A. C.; López, S. J. A. and Torres, C. J. A. 2016. In vitro activities of Trichoderma species against Phytophthora parasitica and Fusarium oxysporum. Afr. J. Microbiol. Res. 15(10):521-527. [ Links ]
Paul, B. 1999. Pythium periplocum, an aggressive mycoparasite of Botrytis cinerea causing the gray mould disease of grape-vine. FEMS Microbiology Lett. 181(2):277-80. Doi:10.1111/j.1574-6968.1999.tb08855.x. [ Links ]
Perveen, K. and Bokhari, N. A. 2012. Antagonistic activity of Trichoderma harzianum and Trichoderma viride isolated from soil of date palm field against Fusarium oxysporum. African J. Microbiol. Res. 13(6):3348-3353. [ Links ]
Rivera-Jiménez, M. N.; Zavaleta-Mancera, H. A.; Rebollar-Alviter, A.; Aguilar-Rincón, V. H.; García de los Santos, G.; Vaquera-Huerta, H.; Silva-Rojas, H. V. 2018. Phylogenetics and histology provide insight into damping-off infections of ‘Poblano’ pepper seedlings caused by Fusarium wilt in greenhouses. Mycological Progress. 17(11):1237-1249. Doi.org/10.1007/s11557-018-1441-2. [ Links ]
Romero-Arenas, O.; Amaro, J. L.; Damián, M. A.; Valencia de Ita, M. A.; Rivera, A. y Huerta, M. 2017. Biopreparados de Trichoderma spp. para el control biológico de Phytophthora capsici en el cultivo de tomate de Puebla, México. Información Técnica Económica Agraria.113(4):313-324. Doi.org/10.12706/itea.2017.019. [ Links ]
Romero, A. O.; Huerta, L. M.; Damián, H. M. A.; Domínguez, H. F. y Arellano, V. D. A. 2009. Características de Trichoderma harzianum, como agente limitante en el cultivo de hongos comestibles. Rev. Colomb. Biotecnol. 11(2):143-151. [ Links ]
Sabbagh, S. K.; Roudini, M. and Panjehkeh, N. 2017. Systemic resistance induced by Trichoderma harzianum and Glomus mossea on cucumber damping-off disease caused by Phytophthora melonis. Arch Phytopathol. Plant Protec. 50(5-8):375-3788. [ Links ]
Sánchez, G. B. M.; Espinosa, H. E.; Villordo, P. E.; Rodríguez, G. R. y Mora, A. M. A. 2017. Identificación molecular y evaluación antagónica in vitro de cepas nativas de Trichoderma spp. sobre hongos fitopatógenos de raíz en frijol (Phaseolus vulgaris l.) cv. Montcalm. Agrociencia. 1(51):63-79. [ Links ]
SAS Institute Inc. 2002. SAS/STAT User guide. Release 9.0. North Carolina, USA. 1289. [ Links ]
Segarra, G.; Avilés, M.; Casanova, E.; Borrero, C. and Trillas, I. 2013. Effectiveness of biological control of Phytophthora capsici in pepper by Trichoderma asperellum strain T-34. Phytopathol. Mediterránea. 1(52):77-83. [ Links ]
Seifert, K. 1996. Fusarium interactive key. Agric. Agri-Food. Canada. 65 p. [ Links ]
Singh, B. N.; Singh, A.; Singh, S. P. and Singh, H. B. 2011. Trichoderma harzianum-mediated reprogramming of oxidative stress response in root apoplast of sunflower enhances defence against Rhizoctonia solani. European Journal Plant Pathology. 1(131):121-134. [ Links ]
Sinuco, D. C.; Pérez, A. C.; Moreno, N. 2017. Evaluación de la actividad fungicida e identificación de compuestos orgánicos volátiles liberados por Trichoderma viride. Rev. Colomb. Biotecnol. 1(19):63-70. [ Links ]
Sneh B.; Burpee L. and Ogoshi A. 1991. Identification of Rhizoctonia species. American Phytopathology Society, MN. 133 p. [ Links ]
Sonnenbichler, J.; Lamm, V.; Gieren, A.; Holdenrieder, O, and Lotter, H. 1983. A Cyclopentabenzopyranone produced by the fungus Heterobasidion annosum in dual cultures. Phyrochemistry. 22(6):489-1491. Doi.org/10.1016/S0031-9422(00)84043-1. [ Links ]
Steel, R. G. D. y Torrie, J. H. 1986. Bioestadística: principios y procedimientos. 2a (Ed.). México: McGraw-Hill. 228 p. [ Links ]
Vásquez, L. A.; Tlapal, B. B.; Yáñez, M. M.; Pérez, R. P. y Quintos, M. E. 2009. Etiología de la marchitez del ‘chile de agua’ (Capsicum annuum L.) en Oaxaca, México. Rev. Fitotec. Mex. 2(32):127-134. [ Links ]
Verma, M.; Brar, S. K.; Tyagi, R. D.; Sahai, V.; Prévost, D.; Valéro, J. R. and Surapalli, R. Y. 2007. Bench-scale fermentation of Trichoderma viride on wastewater sludge: rheology, lityc enzymes and biocontrol activity. Enzyme and Microbial Technology. 41(6):764-771. [ Links ]
Woo, S. L. and Lorito, M. 2007. Exploiting the interactions between fungal antagonists, pathogens and the plant for biocontrol. In: ovel biotechnologies for biocontrol agent enhancement and management. Vurro, M; Gressel, J. N. (Eds.). Springer, Dordrecht. 107-130 pp. [ Links ]
Zegeye, D. E.; Santhanam A.; Gorfu, D.; Tessera, M. and Kassa, B. 2011. Biocontrol activity of Trichoderma viride and Pseudomonas fluorescens against Phytophthora infestans under greenhouse conditions. Inter. J. Agric. Technol. 6(7):1589-1602. [ Links ]
Received: May 01, 2019; Accepted: August 01, 2019
 Este es un artículo publicado en acceso abierto bajo una licencia
Creative Commons
Este es un artículo publicado en acceso abierto bajo una licencia
Creative Commons
