










Serviços Personalizados
Journal
Artigo
 texto em
texto em  Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista mexicana de ciencias agrícolas
versão impressa ISSN 2007-0934
Rev. Mex. Cienc. Agríc vol.10 no.5 Texcoco Jun./Ago. 2019 Epub 03-Mar-2020
https://doi.org/10.29312/remexca.v10i5.1828
Articles
Anthocyanins and color in grain and cob of peruvian purple corn grown in Jalisco, Mexico
1 Departamento de Ingeniería Agroindustrial-Universidad Autónoma Chapingo. Carretera México-Texcoco, km 38.5, Chapingo, Estado de México, México. CP. 56230. (andaaghe-94@hotmail.com).
2 Campo Experimental Centro Altos de Jalisco-INIFAP. Av. Biodiversidad núm. 2470, Col. Las Cruces, Tepatitlán, Jalisco, México. CP. 47600. (ramirez.joseluis@inifap.gob.mx; bautista.edgardo@inifap.gob.mx; flores.hugo@inifap.gob.mx).
The grain and cob (Zea mays L.) of purple corn are a source of pigments, which can replace artificial colors. The objective was to determine the effect of the environment (temperature) of seed production on the content of anthocyanins in grain (CATg) and cob (CATo) and its correlation in a purple corn population of Peru. Seed production environments were Tepatitlan and La Huerta, Jalisco, Mexico. The seeds of the two environments were sown in the S-S 2015 cycle in Tepatitlan, Jalisco. They were pollinated from 100 to 130 plants. At harvest, the healthy ears with the intense purple grain color were analyzed by CATg and CATo by spectrophotometry. An analysis of variance and correlation of Person was made. The environment of seed production significantly affected (p≤ 0.05) the contents of anthocyanins in grain and cob. The seed obtained in Tepatitlan (average temperature 19.1 °C) presented a CATg of 496.3 mg equivalents of cyanidin 3-glucoside in 100 g of dry sample, which was 57.6% higher than that of the seed from La Huerta (mean temperature 25.4 °C). The cob presented between 48 and 70% more anthocyanins than the grain and similar chromatographic profile between grain and cob. The correlation between the grain color variables and the CATg was not significant (p≥ 0.05). However, the negative correlation of the chroma value with CATo was highly significant (r= -0.5484**) so it could be an estimator of the CAT in this structure.
Keywords: Zea mays L.; cob; grain; pigments; purple corn; temperature
El grano y olote de maíz (Zea mays L.) de color morado son fuente de pigmentos, que pueden sustituir a los colorantes artificiales. El objetivo fue determinar el efecto del ambiente (temperatura) de producción de semilla en el contenido de antocianinas en grano (CATg) y olote (CATo) y su correlación en una población de maíz morado de Perú. Los ambientes de producción de semilla fueron Tepatitlán y La Huerta, Jalisco, México. Las semillas de los dos ambientes se sembraron en el ciclo PV 2015 en Tepatitlán, Jalisco. Se polinizaron de 100 a 130 plantas. A la cosecha, las mazorcas sanas y con el color de grano morado intenso, fueron analizadas por CATg y CATo mediante espectrofotometría. Se hizo un análisis de varianza y correlación de Person. El ambiente de producción de semilla afectó de manera significativa (p≤ 0.05) los contenidos de antocianinas en grano y olote. La semilla obtenida en Tepatitlán (temperatura. media 19.1 °C) presentó un CATg de 496.3 mg equivalentes de cianidina 3-glucósido en 100 g de muestra seca, que fue 57.6% superior al de la semilla procedente de La Huerta (temp. media 25.4 °C). El olote presentó entre 48 y 70% más antocianinas que el grano y perfil cromatográfico similar entre grano y olote. La correlación entre las variables de color de grano y el CATg no fue significativa (p≥ 0.05). Pero, la correlación negativa del valor de croma con CATo fue altamente significativa (r= -0.5484**) por lo que podría ser estimador del CAT en esta estructura.
Palabras clave: Zea mays L.; grano; maíz morado; olote; pigmentos; temperatura
Introduction
Interest in food free of artificial colors has increased, possibly due to the criticism of the innocuousness of some of the synthetic dyes, particularly red allura (red 40), red 3 and red 4, which are related to hyperactivity in children (Arnold et al., 2012). Anthocyanins are phenolic compounds responsible for the colors in pink to purple, which have flowers and fruits. Anthocyanins can be obtained from the grain of some varieties of corn (Zea mays L.) in which they are found in high amounts, so they can be sources of natural pigments to replace some synthetic dyes used in food.
The content of total anthocyanins (CAT) in the grain of corn varies according to the color of the grain and the genotype, the deep purple grains have higher CAT than the blue, purple or red grains (Espinosa et al., 2009). In Mexican corns, 21 accessions of the Olotillo breed, with blue-purple grain, showed a variation in CAT between 276 and 904 mg equivalents of cyanidin 3-glucoside (ECG) kg-1 of dry sample (MS) (Salinas-Moreno et al., 2012a), while, in six accessions of the Elotes Conicos breed, with this same grain color (purple blue), the variation was 997-1332.2 mg ECG kg-1 MS (Salinas-Moreno et al., 2012b).
In grains of S2 lines derived from the purple corn population of San Juan Ixtenco, Tlaxcala, values were reported between 130 and 3090 mg ECG kg-1 of sample, doubling that obtained in blue-purple grain corn (Mendoza-Mendoza et al., 2017); however, the content was lower than quantified in purple corn cob from Peru that ranged between 2900 and 13330 ECG kg-1 of sample, which is valued as a possible commercial source of pigments (Yang and Zhai, 2010).
Purple corn is a common corn variety in Ecuador and Peru (Jing et al., 2007) used for the commercial extraction of anthocyanins from the grain and cob, for use in food (Yang and Zhai, 2010) Some varieties of this Corn have been developed to be used for this purpose in different regions of the world (Zhao et al., 2009; Lago et al., 2014). For this reason, although in Mexico there is a great diversity of native corn with anthocyanins in grain and cob, it is preferable to use an exotic material as a donor to accelerate the process of obtaining varieties with high CAT.
The synthesis and accumulation of anthocyanins are processes governed by the genetic information of the material and the interaction with its environment (Gazula et al., 2005). Of the environmental factors, the ones with the greatest impact on the process of anthocyanin biosynthesis are temperature and luminosity (Jaakola, 2013). In purple corn, high temperature reduces the accumulation of anthocyanins, while its synthesis increases at low temperatures (Jing et al., 2007).
In the genetic improvement of corn, two cycles per year are relevant to shorten the time to obtain improved varieties (Ramírez et al., 2015). However, the effect of any increase and corns selection of purple in the autumn-winter agricultural cycle where the temperature rises in April and May is unknown, which may affect the synthesis and accumulation of anthocyanins.
The objectives of the study were: a) to determine the effect of the production environment of the seed on the content of anthocyanins in grain (CATg) and cob (CATo) of a population of purple corn of Peru, under the hypothesis that the effect that has the environment of seed production on the CATg and CATo variables are not inherited to its subsidiary; and b) analyze the relation of the color variables in grain and cob, with the content of anthocyanins, to assess if it is possible to make selection by color of grain for anthocyanin content.
Materials and methods
Increase in plant material
It is started with a sample of 1 kg of the population of purple corn of Peruvian origin ‘Kculli’, provided by the company Altecsa, SA de CV, located in Huamantla, Tlaxcala.
This sample was multiplied, through fraternal crosses in the spring-summer cycle (SS) 2014, under temporary conditions in the Experimental field Center-Altos de Jalisco (CECAJ) of INIFAP, located in the municipality of Tepatitlan de Morelos, Jalisco at 1 900 m of altitude, in the geographic coordinates 20° 49’ 01’’ north latitude and 102° 43’ 59’’ west longitude. A sample of the seed that was obtained in the SS 2014 cycle was multiplied by fraternal crosses in the autumn-winter (AW) cycle 2014-2015, under irrigation conditions, in the Experimental Site Costa de Jalisco, located in the municipality of La Huerta, Jalisco at 300 m altitude at the geographic coordinates 19° 29’ 00’’ north latitude and 104° 39’ 00’’ west longitude. As for the harvest, purple corn of Peruvian origin was obtained from two environments.
The seeds from the two environments were sown in the agricultural cycle SS 2015 in the CECAJ in plots of six rows of 4 x 0.8 m. The multiplication was carried out through fraternal crosses with pollination between 100 and 130 plants. During the harvest, 26 ears of corn were selected for grain health and characteristic purple color in grains and grains of the seed from La Huerta, Jalisco (284#), while 17 seeds were obtained from the seed from Tepatitlán, Jalisco (283#). Subsequent analyzes were performed on grains and cobs of the selected ears.
The maximum and minimum temperatures that were presented from the sowing to the Kculli harvest were obtained from the climatic stations of the experimental fields where the seed increase was made.
Color of grain and ear of selected ears
The color (L*, brightness, a* and b*) was determined in three different points of the middle part of each ear with a Hunter Lab MiniScan XE Plus® colorimeter (Model 45/O-L) in CIELab scale. The parameters a* and b* were used to calculate the hue angle (Hue°) and the color saturation index (chroma) (Jha, 2010).
Preparation of the sample for analysis
The middle section of each ear was shelled the grains by ear (between 100 and 150), and the corresponding ear portion were placed in paper envelopes. Said sample was used for the physical analyzes of grain and CAT. This last determination was made; from the flour obtained from a sample of 25 grains, to which the germ was removed manually with the help of a scalpel, since purple grain corn like ‘Kculli’ lacks anthocyanins in its germ and when it is eliminated Interference of the oil in the CAT variable is avoided. The non-germ grains were milled in a cyclonic type mill (UDY Corporacion, Cyclone sample mill USA) with 0.5 mm mesh. The portion of cob of the middle part of the ear was crushed with a marro and subsequently ground in a Retsch mill (ZM200, 1215030814P, Germany), with a mesh of 0.75 mm.
Physical analyzes performed on grain samples
The weight of 100 seeds (PCS) was quantified in a semi-analytical balance (Sartorius BL610), with two repetitions per ear. For pericarp thickness (EP), 10 healthy grains were randomly taken and soaked in water for 20 min at room temperature. With the help of a scalpel the pedicel and the crown were cut, making lateral cuts to the grain to separate the pericarp.
Five layers of pericarp were placed opposite the germ and the reading was taken in mm in the central part of the group of layers, with a digital micrometer (Digimatic Mitutoyo 293). The result was divided by five to obtain the EP of a layer. The methodologies of physical characterization of grain were applied as described by Salinas and Vázquez (2006).
The type of endosperm was evaluated in the same 10 grains to which the pericarp was removed, which were cut in the longitudinal plane, to obtain two halves, which were classified visually based on the proportion of the floury endosperm, according to what was proposed by Bedolla and Rooney (1982), where: 1= 100% floury; 2= 75%; 3= 50%; 4= 25%; and 5= 100% crystalline.
Quantification of total anthocyanin content in grain (CATg) and cob (CATo)
In both cases (CATg and CATo) the methodology described by Moreno et al. (2005). A standard curve of cyanidin 3-glucoside (Extrasinthase, France) was prepared to express the results in mg equivalents of cyanidin 3-glucoside (ECG) in 100 g of dry sample (MS).
Profile of anthocyanins in grain and cob corn purple by high performance liquid chromatography (HPLC)
The analysis was carried out with the methodology of Fossen et al. (2001) and the modifications of Moreno et al. (2005). Extracts of grain and cob samples were selected with anthocyanin contrasting contents from each environment. A couple of samples for each seed source. The identity of the peaks was made by comparing the retention times and UV-visible spectrum of commercial standards of cyanidin 3-glucoside (Cy 3-glu), pelargonidin 3-glucoside (Pg 3-glu) and peonidin 3-glucoside (Pn 3-glu) (Polyphenols, Nw) and considering previous works in the analysis of the content of anthocyanins in purple corn (Moreno et al., 2005; Cuevas-Montilla et al., 2011).
The identification of acylated type anthocyanins was achieved by analyzing the sample after alkaline hydrolysis (de Pascual-Teresa et al., 2002). With this hydrolysis the acylated anthocyanins are eliminated and do not appear in the chromatogram.
Statistical analysis of the data
The data were analyzed under a completely randomized design, in which the populations obtained from seeds from La Huerta and Tepatitlan, Jalisco were considered as treatments. As repetitions, the number of ears selected within each population was considered, which were 26 and 17 ears of corn. All the variables were analyzed in duplicate. With the data of EP, PCS, CATg, CATo and the color variables of grain and cob, an analysis of variance was performed by means of the GLM procedure of the SAS package, version 9.0. (SAS Institute, 2002). Multiple means comparison tests were also obtained (Tukey, p≤ 0.05). With the variables of color in grain and cob, CATg and CATo, a Pearson correlation analysis was performed using the CORR procedure of the SAS package, version 9.0 (SAS Institute, 2002).
Results and discussion
Grain color and cob
The hue of the grain varied between 9.53 to 34.15° (283#) and 15.38 to 39.97° in the 284#, the values of L* were between 13.2 to 18.56% (283#) and 13.19 to 15.47 (284#), the chroma ranged from 0.94 to 4.7 (283#) and 1.19 to 2.95 (284#). There was no separation in the distribution of the variable Hue° for grain, by origin of seed (Figures 1A and 1B), although the grains with the highest values (close to 40°) were presented within 284# and corresponds to an orange red tone; those with the lowest values (close to 10°) were the grains of 283# and correspond to a purple red tone. The proportion of grains with contrasting tones was low and in general the grain presented a dark red-purple color.
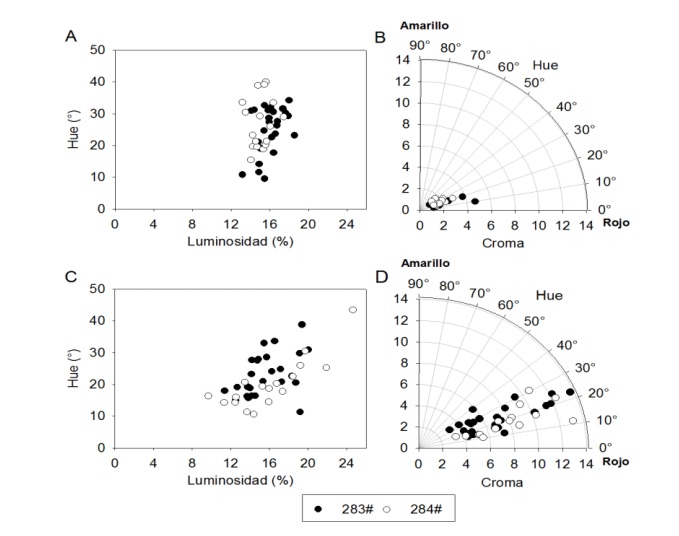
Figure 1 . Distribution in the hue-luminosity plane for grain (A) and cob (C), in the hue-chroma plane for grain (B) and cob (D) of the samples from two environments, evaluated in Tepatitlan, Jalisco México.
In cob, Hue° values varied between 11.34 and 38.77° (283#) and 10.64 to 43.47° (284#). For L* the variation was between 11.38 to 20.06% (283#) and 9.71 to 24.66% (284#), while chroma ranged from 3.13 to 13.7 (28#) and 3.3 to 14.98 (284#) (Figures 1A and 1C). The main color differences between grain and cob were presented in L* and chroma, with higher values in cobs than in grain. Both grain and cob have the same color tone, but the cob is darker and more intense.
Lower chroma values indicate darker shades and are associated with higher anthocyanin content (Salinas-Moreno et al., 2012a). Chroma values observed in the grain of 283# and 284# were lower than those reported by Mendoza-Mendoza et al. (2017) for grain of S2 lines of the purple corn population of San Juan Ixtenco, Tlaxcala, probably due to genetic differences that exist between both populations or due to the effects of the formation of lines and their selection.
Physical variables of the grain and content of anthocyanins in grain and cob
The grains of the ears of 283# and 284# had a completely mealy TE, characteristic common in corn with high content of anthocyanins in the grain (Salinas-Moreno et al., 2012a).
In corn destined for the extraction of pigments, grains with a vitreous endosperm ≥50% are desired to facilitate the mechanical removal of peripheral layers rich in pigments (Salinas-Moreno et al., 2005). Therefore, in a pigmented corn breeding program, crossings with glassy grain materials must be made, which decrease the endosperm mealy portion.
The average PCS value was 38.2 g (283#) and 39.6 g (284#) (Table 1), weights higher than the average values reported for the racial groups of Mexico that are between 18.7 and 33 g (Figueroa-Cárdenas et al., 2013). The desirable in corn for the extraction of pigments is low PCS, which is associated with small and medium grain size, which allows greater contact surface per unit weight and higher pigment content ( Salinas-Moreno et al., 2005).
Table 1 Comparison of means for weight of one hundred seeds (PCS), thickness of pericarp (EP) and content of total anthocyanins in grain (CATg) and cob (CATo) of Peruvian purple corn from two environments and evaluated in Tepatitlán, Jalisco Mexico.
|
Plot |
Variable |
PCS (g) |
EP (m) |
CAT†g |
CATo |
|
283# (n=26) |
Average |
38.2 a |
125.3 b |
496.3 a |
967.9 a |
|
Minimum value |
19.4 |
84.2 |
185.8 |
97.5 |
|
|
Maximum value |
57.5 |
255.7 |
1047.8 |
2147.2 |
|
|
284# (n=17) |
Average |
39.6 a |
134.7 a |
201.3 b |
670 b |
|
Minimum value |
26.7 |
96.9 |
36.9 |
21.9 |
|
|
Maximum value |
55.2 |
186.3 |
816.8 |
2187.4 |
|
|
1.36 |
5.65 |
7.08 |
2.58 |
†= the CAT values are expressed in mg ECG 100 g-1 dry sample; n= number of ears of corn selected; DMS= significant minimum difference. Values in the same column followed by the same letter are not statistically different (Tukey, p≤ 0.05).
The EP varied between 84.2 and 255.7 μm in the 283# population, and from 96.9 to 186.3 μm in the 284#. The average values for each seed origin were located below that reported by Cassani et al. (2017) in the pericarp of the grain of the purple corn variety ‘Nero Spinoso’ cultivated in Italy that was 173 ±12 μm, but are superior to those of Espinosa et al. (2009) of 109 and 107 μm in samples of red corn with pericarp pigment.
The CATg and CATo values were statistically different (p≤ 0.05). The grain of the population 283# showed a CATg 57.6% higher than that of the population 284# (Table 1). In cob, this difference was 30% in favor of 283 #. The content of anthocyanins in the cob was greater than in the grain, a trend similar to that reported by Yang and Zhai (2010). On average, the 283# cob presented 48.7% more anthocyanins than the grain, while in 284#, this difference in favor of the cob was 70%.
The values obtained are greater than those of Yang and Zhai (2010), for a hybrid of purple corn grown in China, which were 55.8±1.5 and 92.3 ±2.1 mg 100 g-1 BS, for grain and cob, respectively.
Of the four variables in Table 1, CATg and CATo had a wider distribution with respect to their mean, which reveals a high variability among the ears analyzed. The above is due to the fact that the initial population is a segregant obtained from free pollination. Greater variability was observed in CAT than in physical characteristics of the grain, which is attributed to the number of genes that regulate each characteristic and its interaction with the evaluation and seed collection environment.
It was expected that the subsidiaries obtained in Tepatitlan, Jalisco from seeds from two environments (La Huerta and Tepatitlan, Jalisco) would have similar contents of anthocyanins in grain and cobs, since these are the same population and a characteristic of maternal effect (Ron-Parra et al., 2016), with a different locality of increase, however, this did not happen. One possible explanation is the effect of the diurnal (~ 30 °C) and nocturnal temperature (~ 20 °C) prevailing in Huerta, Jalisco (Figure 2), during the formation and development of the grain that modified the enzymatic battery responsible for the synthesis and accumulation of anthocyanins, whose activity was not recovered when cultivating the seed in Tepatitlan, which has more favorable temperature conditions than in La Huerta. The duration of the crop cycle in La Huerta was around four months, while in Tepatitlan it was almost six months (Figure 2), which is associated with the prevailing temperatures in each site, significantly affecting the synthesis of the anthocyanins.
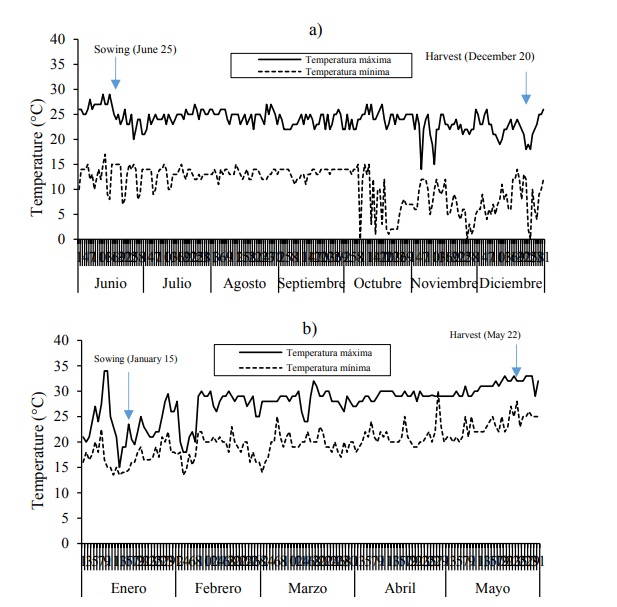
Figure 2 . Maximum and minimum temperatures from sowing to the harvest of Kculli in Tepatitlan, Jalisco during SS 2014 (a) and in Huerta, Jalisco, during the AW 2014-2015 (b).
About 10 enzymes participate in the synthesis of anthocyanins, of which the initial one is phenylalanine ammonium lyase. According to Chon et al. (2012) whose activity is greater under low temperatures both day and night (20/13 °C).
The temperature and light (intensity, quality and duration) are relevant for the synthesis and accumulation of anthocyanins, since they affect the transcription factors related to these processes (Jaakola, 2013). The genes associated with the regulation of anthocyanin synthesis as a response to temperature are independent of those that regulate the expression of light (Azuma et al., 2012). In the cultivation of grapes, greater accumulation of anthocyanins has been observed under conditions of high luminosity and low temperature, than with high luminosity and high temperature.
The results obtained for CATg and CATo in the population of Peruvian purple corn in the two environments of multiplication of the seed indicate that there is a ‘residual’ effect of the high temperatures that results in a low synthesis and accumulation of anthocyanins in the grain and cob; however, it is necessary to carry out further research to determine the suitability of using the AW cycle, within a genetic improvement program of pigmented corn.
Although, the multiplication of the corn seed in the two considered sites was carried out in different humidity regimes: irrigation in La Huerta and temporary in Tepatitlan, reports in vegetable cultivation indicate that the impact of humidity in the accumulation of anthocyanins is lower than that obtained by differences in temperature and luminosity (Biesiada and Tomczak, 2012).
Distribution of anthocyanin content in grains and grains of the ears analyzed
19.2% of the grains of the ears of the plot 283# were located in the lowest interval (0-250 mg ECG in 100 g MS), while, 38.5% presented values in the immediate superior interval (250-500 mg ECG in 100 g MS), in the intervals 501-750 and 751-1000 mg ECG in 100 g MS the same proportion of samples was located (19.2%) and a very low proportion (3.8%) was in the highest interval of anthocyanin content (>1 000 mg ECG in 100 g MS) (Figure 3A).
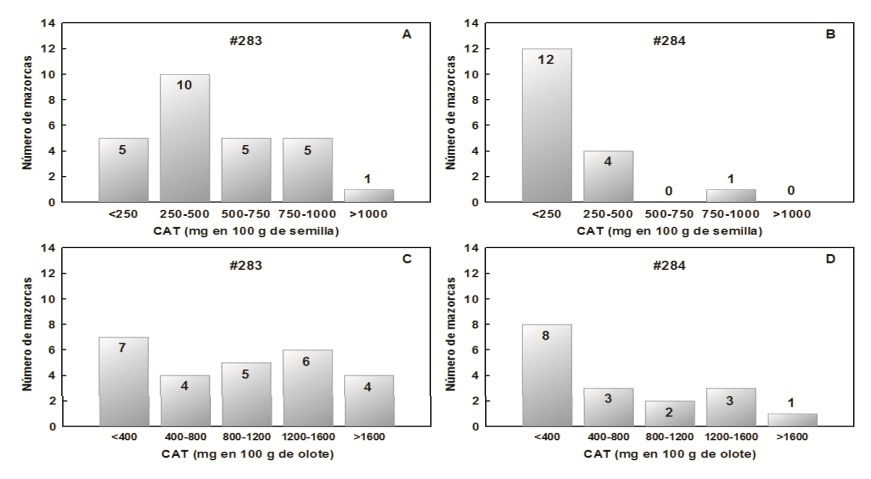
Figure 3 . Distribution by intervals of the content of anthocyanins (CAT, mg in 100 g of dry sample) in grain (A and B) and cob (C and D) in the seed origins of Tepatitlan (283#) and La Huerta (284#).
Regarding the population 284#, 70.5% of the ears were placed in the low range (0 - 250) and 23.5% in the immediate upper interval and only one ear was in the range of 750 to 1000 mg ECG in 100 g MS (Figure 3C). The above is attributed to the fact that in La Huerta, Jalisco the average minimum and maximum temperature was 15 and 30 °C respectively, so the seed did not accumulate anthocyanins in its vacuoles, a condition that it inherited from its subsidiary harvested in Tepatitlan, Jalisco in SS-2015.
In the case of the cob, 283# showed a uniform distribution of the samples between the different anthocyanin content ranges (Figure 3C). However, a greater proportion of samples in the lower range of anthocyanin content was observed in the row on plot 284# (Figure 3D). What is probably due to the temperatures of the seed increased environments.
Chromatographic profile of anthocyanins in grain and cob
Both in grain and in cob, 13 types of anthocyanins were separated (Figure 4 A and B), of which peaks 1, 4, and 5 correspond to simple glycosylated anthocyanins, the other peaks belong to anthocyanins of the acylated type. The difference in the profile of anthocyanins between grain and cob was in the relative percentage of the two main anthocyanins (peaks 1 and 6) corresponding to cyanidin 3-glucoside and cyanidin-3- (6” malonyl glucoside). In grain, the relative proportions of area of these two anthocyanins were similar, while in cob dominated by cyanidin-3- (6” malonyl glucoside). The results obtained in the profiles of anthocyanins for grain coincide with those reported by Cuevas-Montilla et al. (2011) for purple corn grain of Andean variety Kculli variety, but differ with that reported by Yang and Zhai (2010) who reported three and seven anthocyanins in the grain and cob extracts respectively, in a hybrid of purple corn grown in China . The differences can be attributed to the genetic structure of the cultivar, production site and applied analysis protocol.

Identity of the peaks: 1) cyanidin 3-glucoside; 2) cyanidin 3-malonylglucoside*; 3) cyanidin 3-malonylglucoside*; 4) Pelargonidin 3-glucoside; 5) peonidine 3-glucoside; 6) cyanidin-3- (6”malonylglucoside); 7 and 8) acylated anthocyanins; 9) coeluyen pelargonidina-3-(6”- malonylglucoside and peonidina-3-(6”-malonylglucoside); 10) cyanidin-3- dimalonylglucoside; 11) acylated anthocyanin; 12) pelargonidin 3-dimalonylglucoside; and 13) peonidine 3-dimalonylglucoside. *= are considered isomers.
Figure 4 . Chromatographic profiles of grain (A) and ear (B) of purple corn populations.
Correlation of color with anthocyanin content
Of the variables of color in grain, none presented significant correlation with the CAT in this structure. The brightness of the cob (Lo) presented a negative correlation with the CATo in this structure (r= -0.3389**); also, the chroma variable showed negative correlation, and highly significant, with the CATo (r= -0.5484**) (data not shown). These results indicate that the values of brightness and chroma in the cob are lower, the higher will be its CAT.
The correlation between grain and corn CAT, although significant, was low (r= 0.2211*), so the selection of genotypes with high CAT in grain does not ensure that the cob also has a high CAT and vice versa.
Conclusions
The environment where the seed was increased had a significant effect on the content of anthocyanins in grain and cob of the population of purple corn analyzed, with a higher content in the grain increased in Tepatitlan, Jalisco. The grain of the purple corn population Kculli has a higher content of anthocyanins than the grain, with a similar chromatographic profile between both structures, but with differences in the proportion of the two predominant anthocyanins. It is not possible to use any of the variables of color in grain as a criterion to select samples of purple corn for content of anthocyanins since there is no significant correlation between the variables. For cob, the significant negative correlations of luminosity and chroma with their anthocyanin content make possible the selection for anthocyanin content through any of these variables.
Literatura citada
Arnold, L. E.; Lofthouse N. and E. Hurt. 2012. Artificial food colors and attention-deficit/hyperactivity symptoms: Conclusions to die for. Neurotherapeutics. 9(3):599-609. doi: 10.1007/s13311-012-0133-x . [ Links ]
Azuma, A.; Yakushiji H.; Koshita Y. and Kobayashi. S. 2012. Flavonoid biosynthesis-related genes in grape skin are differentially regulated by temperature and light conditions. Planta 236(4):1067-1080. doi: s00425-012-1650-x. [ Links ]
Bedolla, S. and Rooney, L. W. 1982. Cooking maize for masa production. Cereals Food World 27(5):219-221. [ Links ]
Biesiada, A. and Tomczak, A. 2012. Biotic and abiotic factors affecting the content of the chosen antioxidant compounds in vegetables. Vegetable and Crops Research Bulletin. 76(1):55-78. doi: 10.2478/v10032-012-0004-3. [ Links ]
Cassani, E.; Puglisi, D.; Cantaluppi, E.; Landoni, M.; Giupponi, L.; Giorgi, A. and Puli, A. 2017. Genetic studies regarding the control of seed pigmentation of an ancient European pointed maize (Zea mays L.) rich in phlobaphenes: the ‘Nero Spinoso’ from the Camonica valley. Genetic Res. Crop Evol. 64(4):761-773. doi: 10.1007/s10722-016-0399-7. [ Links ]
Chon, S. U.; Boo, H. O.; Heo, B. G. and Gorinstein, S. 2012. Anthocyanin content and the activities of polyphenol oxidase, peroxidase and phenylalanine ammonia-lyase in lettuce cultivars. Inter. J. Food Sci. Nutr. 63(1):45-48. doi: 10.3109/09637486.2011.595704. [ Links ]
Cuevas, M. E.; Hillebrand, S.; Antezana, A. and Winterhalter, P. 2011. Soluble and bound phenolic compounds in different bolivian purple corn (Zea mays L.) cultivars. J. Agric. Food Chem. 59(13):7068-7074. doi: 10.1021/jf201061x. [ Links ]
Pascual-Teresa, S.; Santos-Buelga, C. and Rivas-Gonzalo, J. C. 2002. LC-MS analysis of anthocyanins from purple corn cob. J. Sci. Food Agric. 82(9):1003-1006. doi: 10.1002/jsfa.1143. [ Links ]
Espinosa, T. E.; Mendoza, C. M. del C.; Castillo, G. F.; Ortiz, C. J.; Delgado, A. A. y Carrillo, S. A. 2009. Acumulación de antocianinas en pericarpio y aleurona del grano y sus efectos genéticos en poblaciones criollas de maíz pigmentado. Rev. Fitotec. Mex. 32(4):303-309. [ Links ]
Figueroa-Cárdenas, J. D.; Narváez-Gonzáles, D. E.; Mauricio-Sánchez, A.; Suketoshi, T.; Gaytán-Martínez, M.; Véles-Medina, J. J.; Rincón-Sánchez, F. y Aragón-Cuevas, F. 2013. Propiedades físicas del grano y calidad de los grupos raciales de maíces nativos (criollos) de México. Rev. Fitotec. Mex. 36 (3-A):305-314. [ Links ]
Fossen, T.; Slimestad R. and Andersen O. M. 2001. Anthocyanins from maize (Zea mays L.) and reed canarygrass (Phalaris arundinacea). J. Agric. Food Chem. 49(5):2318-2321. doi: 10.1021/jf001399d. [ Links ]
Gazula, A.; Kleinhenz, M. D.; Streeter, J. G. M. D. and Miller, A. R. 2005. Temperature and cultivar effects on anthocyanin and chlorophyll b concentrations in three related Lollo Rosso lettuce cultivars. HortSci. 40(6):1731-1733. doi:10.21273/HOTSCI40.6.1731. [ Links ]
Jaakola, L. 2013. New insights into the regulation of anthocyanin biosynthesis in fruits. Trends Plant Science 18(9):477-483. doi: 10.1016/j.tplants.2013.06.003. [ Links ]
Jha, S. N. 2010. Colour measurements and modeling. In: Nondestructive evaluation of food quality. Theory and practice. Jha, S. N. (Ed.). Springer-Verlag Berlin Heidelberg. Berlin Alemania 17-40 pp. doi: 10.1007/978-3-642-15796-7_2, 288p. [ Links ]
Jing, P.; Noriega, V.; Schwartz, S. J. and Giusti, M. M. 2007. Effects of growing conditions on purple corncob (Zea mays L.) anthocyanins. J. Agric. Food Chem. 55(21):8625-8629. Doi:10.1021/jf070755q. [ Links ]
Lago, C.; Cassani, E.; Zanzi, C.; Landoni, M.; Trovato, R. and Pilu, R. 2014. Development and study of a maize cultivar rich in anthocyanins: coloured polenta, a new functional food. Plant Breed. 133:201-217. doi: 10.1111/pbr.12153. [ Links ]
Mendoza-Mendoza, C. G.; Mendoza-Castillo, M. C.; Delgado-Alvarado, A.; Castillo-González, F.; Kato-Yamakake, A. y Cruz-Izquierdo, S. 2017. Antocianinas totales y parámetros de color en líneas de maíz morado. Rev. Fitotec. Mex. 40(4):471-479 [ Links ]
Moreno, Y. S.; G. Salas, S. G.; Rubio, H. D. and Ramos, L. N. 2005. Characterization of anthocyanin extracts from maize kernels. J. Chromatographic Sci. 43:483-487. doi: 10.1093/chromsci/43.9.483. [ Links ]
Ramírez-Díaz, J. L.; Ledesma-Miramontes, A.; Vidal-Martínez, V. A.; Gómez-Montiel, N. O.; Ruiz-Corral, J. A.; Velázquez-Cardelas, G. A.; Ron-Parra, J.; Salinas-Moreno, Y. y Nájera-Calvo, L. A. 2015. Selección de maíces nativos como donadores de características agronómicas útiles en híbridos comerciales. Rev. Fitotec. Mex. 38(2):119-131. [ Links ]
Ron-Parra, J.; Rivera, M. M. M.; López, J. J.; Cordero, A. A. J. M.; Larios, L. D. L. C. and González, J. J. S. 2016. Maternal genetic inheritance of red pericarp in the grain of maize. Maydica 61(2016): M21. [ Links ]
Salinas, M. Y. y Vázquez, C. G. 2006. Metodologías de análisis de la calidad nixtamalera-tortillera en maíz. CIRCE-CEVAMEX-INIFAP. Folleto técnico núm. 23. 91 p. [ Links ]
Salinas-Moreno, Y.; Rubio-Hernández, D. y Díaz-Velázquez, A. 2005. Extracción y uso de pigmentos del grano de maíz (Zea mays L.). Arch. Latinoam. Nutr. 55(3):293-298. [ Links ]
Salinas-Moreno, Y.; Cruz-Chávez, F. J.; Díaz-Ortíz, S. A. y Castillo-González, F. 2012a. Granos de maíces pigmentados de Chiapas, características físicas, contenido de antocianinas y valor nutracéutico. Rev. Fitotec. Mex. 35(1):33-41. [ Links ]
Salinas-Moreno, Y.; Pérez-Alonso, J. J.; Vázquez-Carrillo, G.; Aragón-Cuevas, F. y Velázquez-Cardelas, G. A. 2012b. Antocianinas y actividad antioxidante en maíces (Zea mays L.) de las razas Chalqueño, Elotes cónicos y Bolita. Agrociencia. 46(7):693-706. [ Links ]
SAS. 2002. SAS/STAT User’s Guide: Statistics. SAS Institute, Inc. Cary, NC, USA. 5136 p. [ Links ]
Yang, Z. and Zhai, W. 2010. Identification and antioxidant activity of anthocyanins extracted from the seed and cob of purple corn (Zea mays L.). Innovative Food Sci. Emerging Technol. 11:169-176. doi: 10.1016/j.ifset.2009.08.012. [ Links ]
Zhao X.; Zhang, C.; Guigas, C.; Corrales, Ma. Y.; Tauscher, B. and Hu, X. 2009. Composition, antimicrobial activity, and antiproliferative capacity of anthocyanin extracts of purple maize from China. Eur. Food Res. Technol. 228(5):759-765. doi: 10.1007/s00217-008-0987-7. [ Links ]
Received: May 01, 2019; Accepted: August 01, 2019
 Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
