











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
El cacao Theobroma cacao L. (Malvales: Malvaceae), es un cultivo de gran importancia económica, social y ambiental, además de ser una especie primordial en el sistema agroforestal campesino (Jaimes y Aránzazu, 2010).
En México la superficie cosechada del cultivo de cacao para 2017 correspondió a 59 837.8 ha, con una producción de 27 287.25 t, y un valor de la producción nacional de $1 074 303.38. La producción del cacao está concentrada en los estados de Tabasco, Chiapas y Guerrero con 17 430.21, 9 611.63 y 245.41 t, respectivamente (SAGARPA-SIAP, 2018). En el estado de Chiapas, el cacao se cultiva en cuatro regiones agronómicas: Soconusco, Norte, Centro y Selva Palenque, ocupando el sexto lugar en superficie cultivada, detrás del maíz, café, frijol, palma de aceite y mango. De las variedades mexicanas de cacao, en la región Soconusco se cultiva el criollo, por su agradable sabor y aroma, aunque con mayor susceptibilidad a las enfermedades con respecto a otras variedades importantes como el trinitario y el forastero que cuentan una mejora genética resultado del entrecruzamiento con el criollo (González y Amaya, 2005).
Diversos factores, como la escasa tecnología para el manejo del cultivo, el uso de materiales de baja calidad agronómica y la frecuencia de enfermedades por hongos, insectos, aves y roedores, han afectado la calidad y producción del cultivo, generando una crisis de desabasto, que obliga al país a la importación de semilla (González y Amaya 2005; Hernández et al., 2015). De todos estos factores, las enfermedades son las que provocan pérdidas considerables de 40-100% de la producción (Jaimes y Aránzazu, 2010; Hernández et al., 2015).
En el agroecosistema cacao, la enfermedad con mayor distribución es la mancha negra causada por Phytophthora palmivora Butl. /Phytophthora capsici Leonian (Peronosporales: Pythiaceae), la escoba de bruja Moniliophthora perniciosa (Sthael) y la moniliasis del cacao Moniliophthora roreri (Cif.) H. C. Evans, Stalpers, Samson y Benny (Agaricales: Marasmiaceae), siendo estas dos últimas las más destructivas de dicho cultivo, causando pérdidas que oscilan de 50 a 80% de la producción (Aime y Phillips-Mora, 2005). En Villa de Comaltitlán Chiapas, se observaron frutos de cacao con características típicas de pudrición, con esporulación en forma de polvo blanco sobre el tejido, manchas de color café/chocolate, aceitosas/brillosas y con forma de giba. El objetivo fue identificar hongos asociados a frutos de cacao y tejido vegetal con síntomas pudrición.
Materiales y métodos
Recolección de frutos de cacao
Se recolectaron 100 frutos (mazorcas) de 15-20 cm de longitud del cultivo de cacao, con signos típicos de pudrición, en cinco puntos de una parcela no comercial en el municipio de Villa de Comaltitlán, Chiapas (15° 13’ 00’’ latitud norte 92° 34’ 00’’ longitud oeste). El material biológico se colocó en bolsas de polietileno transparente estériles con cierre hermético y colocadas a temperatura ambiente (25 °C) en hieleras de poliestireno expandido y trasladado al laboratorio de Fitopatología del Departamento de Parasitología de la Universidad Autónoma Agraria Antonio Narro (UAAAN) Saltillo, Coahuila.
Aislamiento y purificación del hongo
El aislamiento del hongo para la obtención de colonias puras, se llevó a cabo por dos vías, la primera consisto en tomar esporas directamente del tejido dañado de la mazorca, utilizando una aguja de disección estéril y sembradas en medio de cultivo papa dextrosa agar (PDA) (Bioxon®) y la segunda vía consistió en seccionar tejido vegetal enfermo con un bisturí, el cual se desinfectó con un triple lavado con hipoclorito al 3%, agua destilada estéril (dH2O), alcohol al 70% y dH20 y sembrado en medio de cultivo PDA. En ambas vías de aislamiento, se incubarón a 27 ±2 °C en una cámara de crecimiento (Binder®).
Identificación morfológica, morfométrica y molecular
La identificación morfológica se realizó mediante montas de estructuras del micelio y sus respectivas conidias y esporas en porta y cubreobjetos, con una solución de lactofenol y azul de algodón (para teñir estructuras hialinas). Con ayuda de un microscopio compuesto se observaron las estructuras a 40 y 100x y se identificaron apoyó de las claves taxonómicas para géneros de hongos imperfectos de Barnett y Hunter (1998)).
La evaluación morfométrica se realizó a partir de montas en porta y cubreobjetos de las estructuras del micelio (conidios, hifas, esporas, septos y ramificaciones) en microscopio digital con cámara integrada (AM4023x) y con apoyo del software de medición DinoCapture 2 para la obtención de las dimensiones de las estructuras evaluadas.
La identificación molecular se realizó para el hongo Nodulosporium sp., por la poca información en cultivos de importancia y se realizó mediante la extracción y evaluación de ADN del hongo, siguiendo el método de CTAB (Almeyda et al., 2001) y posteriormente el método de PCR, utilizando los iniciadores universales ITS4 (5’-TCC TCC GCT TAT TGA TAT GC-3’) e ITS5 (5’-GGA AGT AAA AGT CGT AAC AAG G-3’) (White et al., 1990). La amplificación y visualización del producto del PCR se realizó con base en el protocolo de Ahrens y Seemüller (1992)), con modificaciones en las reacciones de PCR y cuyo volumen fue de 25 μL [agua ultrapura estéril: 11.9 μL, Buffer 10X (2.5 μL) dNTPs a 10 mM (0.4 μL), iniciadores 1TS4 e ITS5 a 5 mM (3 μL de cada uno), ADN polimerasa (Green Taq DNA Polimerase, GenScript®) a 1U (0.2 μL) y muestra problema de ADN a 96.7 ng (4 μL)].
Las condiciones de la reacción de PCR fueron: 1 ciclo de desnaturalización inicial a 95 ºC por 3 min, 35 ciclos de desnaturalización a 95 ºC por 10 s, 35 ciclos de alineamiento a 57 ºC por 30 s, 35 ciclos de extensión a 72 ºC por 45 s y 1 ciclo de extensión final a 72 ºC por 5 min. El producto amplificado se visualizó en un gel de agarosa al 1% mediante electroforesis. El producto de PCR se purificó con el kit de bandas In vitro gen (QuickClean II Gel Extraction Kit [100rxns], GenScript®) y el producto purificado se envió para su secuenciación en dos direcciones (5’ a 3’ y 3’ a 5’) al Laboratorio Nacional de Biotecnología Agrícola, Médica y Ambiental (Lanbama) del Instituto Potosino de Investigación Científica y Tecnológica AC (Ipicyt), el resultado obtenido se comparó con las secuencias en la base de datos del banco de genes del National Center for Biotechnology Information (NCBI, 2018).
Suspensión de esporas
La fuente de inóculo se obtuvo a partir de esporas del medio PDA, obtenidas con una varilla de vidrio estéril y se colocaron en un tubo de ensayo de 10 mL diluidos en agua destilada estéril. El conteo se realizó en cámara de Newbauer para determinar la concentración de esporas en la suspensión (1x108 esporas mL-1).
Pruebas de patogenicidad
Se recolectaron frutos de 15-20 cm sanos de plantas sin presencia de algún tipo de daño. La inoculación en los frutos se realizó por punción, con aguja de disección colonizada con esporas del hongo Nodulosporium cultivado en medio PDA, colocando el inóculo en la parte distal (punta) del fruto. En esta prueba se utilizaron 10 frutos y se realizaron cuatro réplicas, colocadas sobre papel estraza estéril durante el estudio.
La inoculación en follaje se realizó con la técnica de hoja desprendida (Pettitt et al., 2011) sobre una charola plástica de 25 cm, con una suspensión de 1x108 esporas mL-1, colocando esta suspensión sobre la superficie adaxial (haz) en un total de 10 hojas, replicado tres veces. En ambas pruebas se consideró un testigo absoluto en el que solo se aplicó agua destilada estéril siguiendo la misma metodología. Las pruebas se desarrollaron bajo condiciones controladas a una temperatura de 25 ±2 °C, humedad relativa de 70 ±5% y 12:12 luz: obscuridad. Además, la prueba con follaje se hidrató con una pequeña torunda de algodón saturada de agua estéril y reponiendo esta humedad cuando fue necesario. Cuando aparecieron síntomas en el tejido inoculado, se re-aisló el patógeno en medio de cultivo PDA y se identificó nuevamente.
Resultados y discusión
Identificación morfológica, morfométrica y molecular
En medio de cultivo PDA se aislaron diversos hongos de esporas del tejido dañado de la mazorca y tejido vegetal enfermo con síntomas de pudrición en cacao. Cada hongo fue purificado e identificado, encontrando Aspergillus sp. Micheli (Trichocomaceae), Penicillium sp. Link (Trichocomaceae), Rhizopus sp. Ehrenb (Mucoraceae), Trichoderma sp. Persoon (Hypocreaceae) con una incidencia de 85% y Nodulosporium sp. Preuss (Xylariaceae) con 15% de incidencia en los aislados. Se destaca la ausencia del hongo M. roreri, el cual es considerado como el agente causal de la pudrición de mazorca o moniliasis en diversas regiones productoras de cacao en México (Phillips-Mora et al., 2006; Phillips-Mora et al., 2007).
En este estudio se registra por primera ocasión la presencia del hongo Nodulosporium (= Nodulisporium) en frutos del cultivo de cacao en Chiapas, México. En México, el género Nodulosporium se ha encontrado en mezquite (Prosopis laevigata L.) (Fabaceae) en el estado de Puebla (De la Torre-Almaráz et al., 2009), en el cáliz de Flor de Jamaica Hibiscus sabdariffa L. (Malvaceae) en el estado de Guerrero (Ruiz-Ramírez et al., 2015) y en cacahuananche Gliricidia sepium (Jacq.) Kunth ex. Walp. (Fabaceae) en el estado de Morelos (Sánchez- Fernández, 2016). En el cultivo de cacao, el género Nodulosporium ha sido mencionado por Márquez-Dávila et al. (2013)), aislado de hojas y tallos de cacao nativo en cuencas del alto amazonas de Perú.
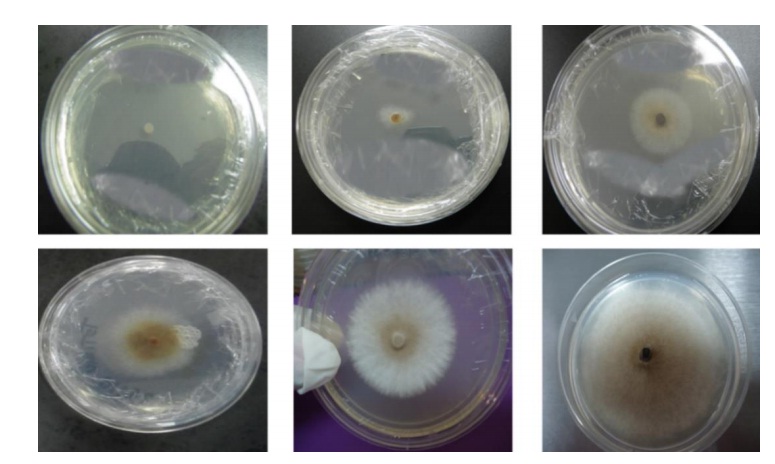
Nodulosporium sp., presentó un llenado de la caja de Petri (9 cm diámetro) en 7 días, desarrollando al principio una colonia micelial de color blanquecino y de aspecto algodonoso, 5 días después de la siembra se presentó la maduración de conidios, tornándose de color beige (similar a lo observado en el tejido enfermo), posteriormente al séptimo día, las colonias maduras de conidios adquirieron una coloración café-obscura (Figura 1).
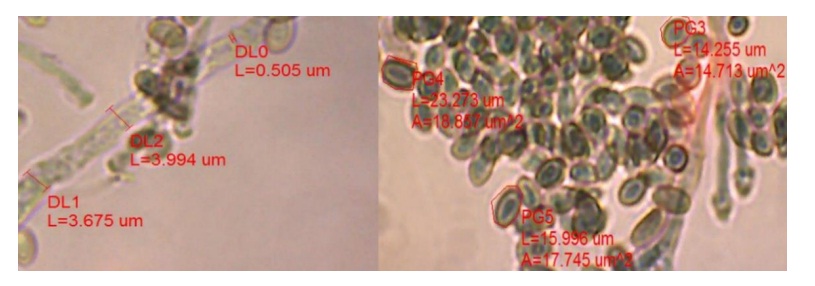
Nodulosporium sp., presenta hifas poco septadas de color café claro, que se tiñen de color azul claro al centro (efecto del lactofenol y azul de algodón), conidioesporas en el conidióforo de forma sarcinada, hialinas alimonadas, que tomaron un color verde claro (efecto del lactofenol y azul de algodón). La pared o membrana de la espora ligeramente amarilla (Figura 2).
Además de presentar un olor característico a musgo.
La observación al microscopio digital permitió ver las estructuras del micelio, con un tamaño (L= longitud) de conidios de 117.189-117.589 µm, grosor de hifas de 3.675-3.994 µm, grosor de septos de 0.505 µm, largo de ramificaciones de 34.797-75.848 µm, grosor de ramificaciones de 3.194-3.761 µm, esporas de 13.415-23.273 µm en forma poligonal (la periferia) y un área de 12.816-21.666 µm2 (A= área) y un grosor de la membrana de la espora de 1.197-1.249 µm (Figura 2 y 3).

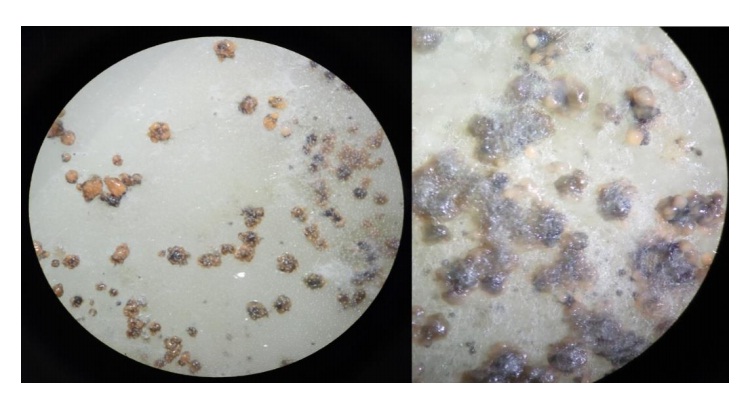
De las pruebas de patogenicidad en follaje se logró recuperar el hongo, el cual fue aislado del tejido y puesto en medio de cultivo PDA, de este solo se obtuvo micelio y conglomerados de hifas (picnidios), con una sustancia gelatinosa (mucosidad) de color naranja-claro sobre y en la periferia del conglomerado (Figura 4).
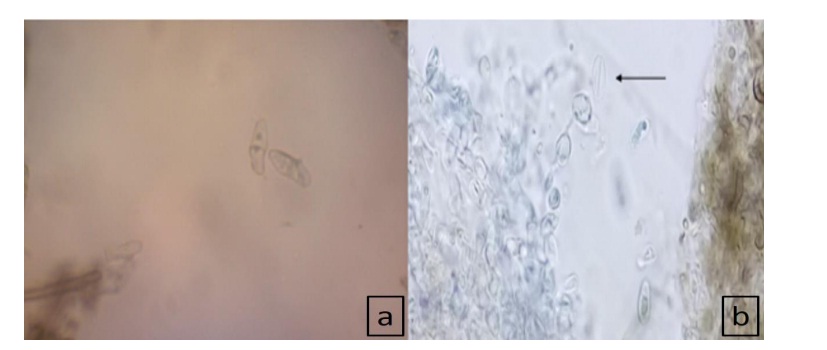
Estas estructuras fueron montados en porta y cubreobjetos con solución de lactofenol y azul de algodón, donde se observó la formación de estromas con esporas, las cuales son de color hialino (nunca se tornaron obscuras, incluso ante obscuridad durante un periodo de 24 días), unicelulares, elipsoides-inequilaterales, con extremos ligeramente más redondeados, con abertura germinal alargada (11.5 µm) y con presencia de periespora dehiscente (largo:13.188-14.526 µm; ancho 4.342-5.427 µm) hialina (Figura 5a, 5b), estructuras características que con base en las claves de Barnett y Hunter (1998)), se identifica como el género Hypoxylon Bull. (Xylariaceae) teleomorfo de Nodulosporium, por lo cual ambos están relacionados; en este sentido Ju y Rogers (1996)), reconocen a Nodulosporium como anamorfo o fase sexual de varias especies del género Hypoxylon.
Se obtuvo el producto de PCR del ADN del micelio de Nodulosporium y de picnidios de Hypoxylon; ambos con un producto de 600 pb con los oligonucleótidos ITS4-ITS5. Se obtuvieron 2 secuencias para Nodulosporium y 3 de Hypoxylon, para su comparación en la base de datos del banco de genes del algoritmo BLAST (NCBI, 2018). Se confirmó la identidad del hongo aislado de frutos de cacao con pudrición y el recuperado en la prueba de patogenicidad en follaje obtenidas del municipio de Villa de Comaltitlán en el estado de Chiapas, revelando a Hypoxylon en las 5 secuencias homólogas en dos aislados en el GenBank, procedentes de Estados Unidos de América (número de acceso: KF496192.1) con una identidad de 100% (Cuadro 1).
Cuadro 1 Comparación de secuencias en el banco de genes del NCBI.
| Identificación morfológica | Primer’s de secuencia | Total score | Valor E | Identidad (%) | Núm. acceso | Identificación molecular |
| Hypoxylon sp. | ITS4- ITS5 | 501 | 4e-138 | 100 | KF496192.1 | Hypoxylon sp. |
| Nodulosporium sp. | ITS4- ITS5 | 501 | 4e-138 | 100 | KF496192.1 | Hypoxylon sp. |
| Nodulosporium sp. | ITS4- ITS5 | 501 | 4e-138 | 100 | KF496192.1 | Hypoxylon sp. |
| Hypoxylon sp. | ITS4- ITS5 | 501 | 4e-138 | 100 | KF496192.1 | Hypoxylon sp. |
| Hypoxylon sp. | ITS4- ITS5 | 501 | 4e-138 | 100 | KF496192.1 | Hypoxylon sp. |
Pruebas de patogenicidad
La prueba de patogenicidad con frutos de cacao durante un periodo de 15 días de evaluación esta mostró síntomas leves de crecimiento del hongo inoculado; sin embargo, los frutos de esta prueba provenientes de campo se encontraban naturalmente enfermas con pudrición, incluso se observó esta condición en los frutos que se utilizaron como testigo inoculado solo con agua destilada estéril. La infección natural bajo las condiciones de laboratorio presentó un crecimiento rápido, lo que indica que el interior del fruto ya se encontraba invadido antes de la inoculación. Desde el cuarto día de inoculación, se observaron las primeras manchas asimétricas de color café oscuras y brillantes, posteriormente al octavo día el fruto se encontraba completamente cubierto por la mancha con presencia de mezcla de micelios de colores blanquecinos, naranjas, verdes y café, observando pudrición en alta severidad.
La prueba de patogenicidad con follaje mostró sensibilidad a las 24 h de la inoculación, observándose clorosis sobre la hoja la cual exhibió a partir del punto de inoculación un proceso de deshidratación (desecamiento sin perder el color verde totalmente) y posteriormente a las 48 h presentó crecimiento de micelio blanquecino, quedando de esta manera hasta el término de la prueba (7 días en total) (Figura 6).

Figura 6 a y b) hoja de cacao sin presencia de daño por Nodulosporium sp. (testigo); c y d) presencia de daño por Nodulosporium a los 48 h en follaje de cacao.
Esta lesión se observó solo en 30% de las muestras inoculadas. En el testigo inoculado con agua destilada estéril no se observó ningún síntoma o lesión alguna de la presencia del patógeno. En las muestras sintomáticas fue re-aislado el hongo, encontrando únicamente al hongo inoculado en forma de picnidios y micelio con conidios, correspondiendo al teleomorfo de Nodulosporium, identificado como Hypoxylon. Este resultado indica que Nodulosporium es levemente patogénico en follaje de cacao, causando clorosis y deshidratación de la hoja.
Los géneros Nodulosporium e Hypoxylon se han reportado como fitopatógenos, ocasionando muerte descendente, pudrición del tronco negro y resinosis de ramas de mezquite P. laevigata (De la Torre-Almaráz et al., 2003; De la Torre-Almaráz et al., 2009). Nodulosporium además se ha encontrado provocando pudrición basal del estípite en palma africana de aceite Elaeis guineensis Jacq. (Arecaceae) (Mestizo et al., 2012) y pudrición basal en el cáliz de Flor de Jamaica H. sabdariffa (Ruiz-Ramírez et al., 2015).
El género Nodulosporium se ha asociado como endófito a diversas plantas; Collado et al. (2001)) lo reportan en corteza del tronco, hojas y ramas sanas y secas de Quercus ilex L. (Fagaceae). Salgado y Cepero (2005)) lo encontraron en hojas de Rosa hybrida L. (Rosaceae), Márquez-Dávila et al. (2013)) aislado a partir de hojas y tallos de cacao T. cacao, Lizarazo-Medina et al. (2014)) en hojas de dos especies de orquídeas Cattleya percivaliana y Cattleya trianaei (Orchidaceae) y se aisló también de cacahuananche (G. sepium) con potencial como antagonista contra hongos y oomicetos (Sánchez-Fernández, 2016).
Las especies del género Hypoxylon se han reportado como endófitos en tejidos de plantas leñosas o herbáceas (Petrini y Petrini, 1985), así como saprofitos o fitopatógenos débiles en madera de árboles de zonas templadas o tropicales (Miller, 1961). Para México, este género es el segundo con mayor número de especies en la Familia Xylariaceae (San Martín, 1992) con 41 especies identificadas, 14 registradas para el estado de Chiapas en siete municipios, encontradas sobre madera, principalmente en bosque tropical alto perennifolio, bosque tropical mediano subcaducifolio, bosque tropical bajo caducifolio, bosques de pino encino y bosque mesófilo de montaña (San Martín, 1999).
Reconocer la diversidad de patógenos asociados al cultivo de cacao es de suma importancia, debido a que pueden representar una amenaza para el cultivo en un futuro.
Conclusión
Se identifica al hongo Nodulosporium sp., aislado de tejido vegetal enfermo con síntomas de pudrición en mazorca de Theobroma cacao para el estado de Chiapas y se confirma su teleomorfo identificado como Hypoxylon. Asimismo, su patogenicidad sobre follaje de cacao, causa clorosis y deshidratación de la hoja.

