










Servicios Personalizados
Revista
Articulo
 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de ciencias agrícolas
versión impresa ISSN 2007-0934
Rev. Mex. Cienc. Agríc vol.10 no.3 Texcoco abr./may. 2019 Epub 30-Mar-2020
https://doi.org/10.29312/remexca.v10i3.1540
Articles
Variability in soluble carbohydrates of the stem and its contribution to grain yield in wheat
1Facultad de Ciencias Agrícolas-Universidad Autónoma del Estado de México-Campus Universitario ‘El Cerrillo’. Carretera Toluca-Ixtlahuaca km 15, Entronque El Cerrillo, Toluca, Estado de México. AP. 435. CP. 50200. Tel. y Fax. 01(722) 2965518. (aqua-alpharadio@yahoo.com.mx; adominguezl@uaemex.mx; cgmartinezr@uaemex.mx; ejmoralesr@uaemex.mx).
In wheat, the capacity to store soluble carbohydrates (CHS) in the stem and to remobilize them towards the grain, constitutes a desirable characteristic to be incorporated in the germplasm directed to regions where terminal drought frequently occurs. The objective of the work was to evaluate the variability in the accumulation and remobilization of water-soluble carbohydrates (CHS) and their contribution to grain yield when the post-synthesis source is altered by defoliation and water stress. The experiments were carried out in the summer-autumn 2013 cycle (temporal, exp. 1) and winter-spring 2013-2014 (irrigation, exp. 2) in Toluca, Mexico, at a density of 350 seeds m-2, in the exp 1 the defoliation was performed 7 days after anthesis and in the exp. 2 was not watered after anthesis. In the exp. 1 60 wheat elite lines and two source reduction treatments (defoliated and defoliated, called environments onwards) were studied under a split plot design with 2 replications. The exp. 2 consisted of 20 genotypes selected from exp. 1 and subjected to two water regimes (irrigation and post-flowering drought) in a randomized complete block design with 3 replications where each water regime was considered an environment. Great variability was observed for grain yield (RG) and the associated physiological characteristics. The RG was reduced by an average of 14% due to defoliation and post-anthesis drought. The greater contribution of CHS was in line with a greater efficiency of remobilization. There are promising genotypes for use in breeding programs with a view to incorporating physiological characteristics to increase the potential yield under conditions of biotic or abiotic stress in post-anthesis.
Keywords: Triticum aestivum L.; defoliation; soluble carbohydrates; water stress
En trigo la capacidad para almacenar carbohidratos solubles (CHS) en el tallo y remobilizarlos hacia el grano, constituye una característica deseable a incorporar en el germoplasma dirigido a regiones donde ocurre con frecuencia sequía terminal. El objetivo del trabajo fue evaluar la variabilidad en la acumulación y remobilización de los carbohidratos solubles en agua (CHS) y su contribución al rendimiento de grano cuando la fuente postantesis es alterada por defoliación y estrés hídrico. Los experimentos se llevaron a cabo en el ciclo verano-otoño 2013 (temporal, exp. 1) e invierno-primavera 2013-2014 (riego, exp. 2) en Toluca, México, a una densidad de 350 semillas m-2, en el exp. 1 la defoliación se realizó 7 días después de antesis y en el exp. 2 no se regó después de antesis. En el exp. 1 se estudiaron 60 líneas élite de trigo y dos tratamientos de reducción de fuente (defoliado y sin defoliar, llamados ambientes en adelante), bajo un diseño de parcelas divididas con 2 repeticiones. El exp. 2 consistió en 20 genotipos seleccionados del exp. 1 y sometidos a dos regímenes hídricos (riego y sequía postfloración) en un diseño de bloques completos al azar con 3 repeticiones en donde cada régimen hídrico se consideró un ambiente. Se observó gran variabilidad para rendimiento de grano (RG) y las características fisiológicas asociadas. El RG fue reducido en promedio 14% por efecto de la defoliación y sequía pos-antesis. La mayor contribución de CHS estuvo en línea con una mayor eficiencia de remobilización. Existen genotipos promisorios para usarse en programas de mejoramiento genético con miras a incorporar características fisiológicas para incrementar el rendimiento potencial bajo condiciones de estreses bióticos o abióticos en pos-antesis.
Palabras clave: Triticum aestivum L.; carbohidratos solubles; defoliación; estrés hídrico
Introduction
One of the great challenges facing national and international wheat breeding programs is the search for physiological attributes that allow the crop to cushion the effects of biotic and abiotic stresses. It is now known that water-soluble carbohydrates (CHS) stored in the stem before anthesis contribute to maintain the rate of grain filling when photosynthesis declines due to some stress (Dreccer et al., 2009). In Mexico, the wheat producing areas (Bajío and Northwest) could in the near future be limited by water during the filling of grain, mainly because of the cost that this represents (Ledesma-Ramírez et al., 2012), while in the valleys high fungal diseases defer wheat yields (Hortelano et al., 2013).
In this sense, having efficient germplasm in storing and remobilizing CHS from the stem to the grain in the face of any stress could be important in this region. The filling of grain in wheat depends on the amount of assimilates stored in the stem, as well as the photosynthesis of the leaves and spikes; however, CHS helps maintain the rate of grain filling when photosynthesis decreases due to biotic and abiotic stress (Blum, 1998; Ehdaie et al., 2006). The accumulation, translocation and distribution of CHS in wheat play an important role in grain yield (Zhang et al., 2013).
The higher concentration of CHS in stems is associated with increases in yield and grain weight under stress and non-stress conditions (Shearman et al., 2005; Rebetzke et al., 2008). The contribution of CHS to yield varies according to growth conditions and can range from 10 to 20% without stress (Shearman et al., 2005) to 60% under severe stress (Blum, 1998; van Herwaarden et al., 1998a, b). Recent studies mention that there is genetic variation for the accumulation of CHS in wheat (van Herwaarden and Richards, 2002; Ruuska et al., 2006); however, the genotypic variability to store, remobilize and its contribution to grain yield in elite wheat lines is not yet very clear.
The remobilization of CHS from the vegetative tissues (stems and pods) is induced by early senescence, a situation that occurs when the soil loses moisture rapidly during grain filling (Yang et al., 2000), reducing photosynthesis, shortening the period of filling and final grain weight (Tahir and Nakata, 2005; Zhang et al., 2012). This capacity associated with senescence is also influenced by the environment, genotype and CHS demand for developing grains (Davison and Chevalier, 1992).
Among the associated physiological characteristics of the plant for drought tolerance, the CHS stored in leaves or stems have been considered of great importance because they not only act as a regulator of osmotic adjustment, but are also a carbon source to maintain grain growth when photosynthesis is inhibited by water stress in post-anthesis (Van Herwaarden et al., 2006; Yang et al., 2007). On the other hand, the remobilization of CHS from the stems to the spike changes depending on the genotype and environmental conditions. Mariotti et al. (2003) found that the contribution of CHS in pre-anthesis was 43 to 54% in durum wheat, whereas in barley it ranges from 4 to 24% (Przulj and Momcilovic, 2001).
Ehdaie et al. (2008) found a group of 11 different genotypes of wheat that the current assimilates and the reserves in the stems contributed between 19.1 to 53.6% in irrigation, while in drought the range was from 36.6 to 65.4%. The variability in the accumulated reserves and those that are moved to the grain depends on the genotype, magnitude of the stress and even on the method used to measure the CHS (Ehdaie et al., 2006).
In recent years, genetic improvement has used selection towards a greater accumulation of CHS in the stem as an important tool to improve adaptation to unfavorable environments (van Herwaarden and Richards, 2002; Ehdaie et al., 2006, 2008). It has been mentioned that there is genetic variation for accumulation of CHS in the stem in anthesis and that genetic improvement to increase the amount of CHS may be possible because it is a highly heritable character (Ruuska et al., 2006), but controlled by a complex polygenic regulation (Rebetzke et al., 2008).
Recent studies with modern and old wheat cultivars indicate that the genetic improvement in this species has increased the storage capacity of CHS in the stem of the new varieties (Foulkes et al., 2007). This requires the search of the relationships between the CHS with agronomic characters that allow the breeder to identify progenitors or advanced lines with high contents or with high efficiency of CHS remobilization. Recently the world wheat consortium grouped a set of elite lines in an international trial called CIMCOG (CIMMYT Mexico Core Germplasm Panel) (Foulkes et al., 2011), with genotypes adapted to different regions of the world.
The characterization of said germplasm in response to different types of stress could help to identify cultivars with high capacity to store and remobilize CHS when there are biotic and abiotic stresses post-anthesis.
The objective of the work was to evaluate the variability in the accumulation and remobilization of water-soluble carbohydrates and their contribution to grain yield when the post-anthesis source is altered by defoliation and water stress.
Materials and methods
Two experiments were carried out in the Faculty of Agricultural Sciences of the Autonomous University of the State of Mexico (UAEM), Toluca, Mexico at 19° 15’ 33’’ North latitude, 99° 39’ 38’’ West longitude and 2 640 masl. The climate of this locality is classified as a sub-humid semicold with summer rains, with an annual precipitation range of 800 to 1 300 mm and a temperature of 8 to 14 °C (INEGI, 2009). The experiments were established in a Vertisol-type soil with a clay-loam texture (USDA, 2000).
The genotypes used were 60 wheat lines with good agronomic adaptation that are part of the elite CIMCOG material. The complete collection of the genotypes of the CIMCOG group is potentially useful in breeding programs aimed at further raising the yield potential. For this reason, they are the main germplasm studied up to now by the world wheat consortium (Reynolds et al., 2011).
The exp. 1 was carried out in the summer-autumn (S-A) cycle of 2013 (temporary), which consisted of the factorial combination of the 60 wheat genotypes and two source reduction treatments (D= defoliate and SD= no defoliate), these last named environments. The treatments were distributed in an arrangement of divided plots with 2 replications, where the large plot corresponded to the genotypes and the subplot to the environments.
Seeding was done manually at a population density of 350 seeds per m2 in plots of 2 rows of 3 m long and 0.2 m separation. Seven days after anthesis (Zadoks 60) (Zadoks et al., 1974), the 1 m long plants of the 2 rows of each plot were defoliated manually eliminating all the green leaves.
The exp. 2 was conducted in winter-spring (W-S) 2013-2014, which consisted of the factorial combination of 20 wheat lines (selected from exp.1 for showing different capacity to store and remobilize CHS, but similar cycle before anthesis) and two treatments of water availability (R= irrigation throughout the cycle and S= irrigation until anthesis and post-anthesis drought). In each situation (R and S), the genotypes were distributed in a randomized complete block design with 3 replications. The availability of water was considered as an environment. The cultivars were sown manually at a density of 350 seeds per m2 in plots of 6 furrows 3 m long and 0.2 m from separation.
The water was controlled by drip irrigation. The experiments were kept free of weeds, pests and diseases. The fertilization consisted of the formula 200-60-30, fractionating the nitrogen in the sowing, terminal spikelet (encañe) and expanded flag leaf.
Grain yield and its components
In physiological maturity (Z89) (Zadoks et al., 1974) the 0.4 m2 plants were extracted in each plot. The dry weight of leaves, stems and ears was recorded after drying the samples in a forced air oven for 72 h at 70 °C, until reaching constant weight. The grain yield per m2 (RG) was determined by weighing the grain obtained in the sample and the individual grain weight (PIG) when counting and weighing 1 000 grains.
Laboratory analysis
The CHS content was determined in anthesis (CHSAnt) and physiological maturity (CHSMf), randomly selecting a subsample of 20 fertile main stems (without spike or leaf blade) from the biomass sample of each plot in each treatment. The CHS were extracted and measured using the anthrone method of Yemm and Willis (1954) described by Galicia et al. (2009). The data was obtained as a percentage of CHS in dry matter. These were used to calculate the CHS content per stem (g stem-1) or per unit area (g m-2), according to the procedure described by Pask et al. (2013).
The amount of CHS mobilized from the stem to the spike, the remobilization efficiency and the contribution to the grain yield were calculated using the following formulas (Shakiba et al., 1996): 1. Total amount of CHS remobilized or translocated (CHSTransl)= maximum quantity of CHS-quantity of CHS at maturity; 2. Total remobilization efficiency ERCHS= total amount of CHS remobilized x 100/maximum amount of CHS; 3. Contribution to grain yield (CRG)= total amount of CHS remobilized x 100/grain yield; and 4. Water soluble carbohydrates present in the grain (CHSG).
Statistical analysis
Analysis of variance was carried out according to the models mentioned above (Littell et al., 1996) to evaluate the effect of treatments (environments) on each of the variables studied. When analysis of variance revealed significant differences, the mean values for each treatment were compared using Tukey's 5% honest minimum difference (DMSH) (Palaniswamy and Palaniswamy, 2006) using SAS software (SAS, 2002).
Results and discussion
Effect of treatments on the variables studied
In the analysis of variance, important effects of the treatments were observed on the variables evaluated in both experiments (Table 1 and 2). It can be seen that in the exp. 1 The effect of the genotypes was highly significant (p< 0.01) for all the variables analyzed in the same way for environments with the exception of CHSAnt and the genotype-environment interaction (GxA) was not significant for the PIG (Table 1). The coefficients of variation ranged from 0.9 to 19.3% corresponding to soluble carbohydrates stored in the stem in anthesis and soluble carbohydrates stored at physiological maturity (CHSMf) (Table 1).
Table 1 Values of F and its statistical significance for the variables evaluated in experiments 1 and 2.
| FV | gl |
(g m-2) |
(mg) |
(g m-2) |
(g m-2) |
(g m-2) |
(%) |
(%) |
| Exp. 1 | ||||||||
| Repetition | 1 | 8* | 0.02ns | 0.6ns | 15.1* | 1.8ns | 6.6* | 6.1* |
| Genotypes (G) | 59 | 8** 62 | 5.2** 49 | 20.3** 95 | 8.1** 39 | 22.2** 83 | 14.8** 59 | 26.4** 80 |
| Error to ( CM) | 59 | 1969.7 | 12.6 | 1698 | 508.9 | 1660.1 | 89.8 | 106.6 |
| Environments (A) | 1 | 336.3** 19 | 78.6** 14 | 4ns | 305.1** 20 | 298.3** 4 | 194.5** 11 | 519.7** 8 |
| G x A | 59 | 1.7* 6 | 1.4ns | 4** | 7.8** 31 | 8** 7 | 6.5** 22 | 7.1** 7 |
| Error b (CM) | 60 | 874.7 | 14.7 | 6.6 | 413.9 | 411.2 | 76.2 | 34.4 |
| CV (%) | 6.7 | 8.6 | 0.9 | 19.3 | 11.7 | 15 | 14 | |
| Exp. 2 | ||||||||
| Repetition/A | 4 | 1.3ns | 0.1ns | 0.7ns | 0.7ns | 0.5ns | 0.5ns | 0.1ns |
| Environments (A) | 1 | 52.3* 20 | 448.5** 18 | 446.4** 35 | 1330.7** 54 | 1281.2** 58 | 1841.8** 71 | 4536.6** 62 |
| Genotypes (G) | 19 | 8.2** 48 | 7.8** 48 | 21.5** 44 | 22** 24 | 15.6** 23 | 8.4** 12 | 11.5** 20 |
| G x A | 19 | 0.9ns | 1.3ns | 5.9** 12 | 14.4** 16 | 8** 12 | 8.5** 12 | 6.3** 11 |
| Error (CM) | 76 | 1634.8 | 6.8 | 1420.5 | 128.2 | 1415.9 | 33.7 | 140.6 |
| CV (%) | 10 | 6.8 | 13.3 | 19.9 | 16.7 | 7.6 | 20 |
*= significant (p< 0.05), **= highly significant (p< 0.01) and ns= (p> 0.05) not significant. CM= mean square of the error; FV= source of variation; gl= degrees of freedom; RG= grain yield; PIG= individual grain weight; CHSAnt= water soluble carbohydrates at the time of anthesis; CHSMf = water soluble carbohydrates at physiological maturity; CHSTransl= soluble translocated carbohydrates (difference between CHSAnt and CHSMf); ERCHS= total remobilization efficiency; CRG= contribution to grain yield. Bold numbers indicate the contribution of the factor to the sum of squares total in (%). The bold number indicates the contribution of the factor to the sum of squares total in (%).
Table 2 Mean values for variables evaluated in each environment (D= defoliate, SD= no defoliate, R= irrigation, S= drought). Average as genotypes average.
| Ambient |
RG (g m-2) |
PIG (mg) |
CHSAnt (g m-2) |
CHSMf (g m-2) |
CHSTrasl (g m-2) |
ERCHS (%) |
CRG (%) |
CHSG (%) |
| SD | 475.6 a | 46.3 a | 278.5 a | 127.9 a | 150.6 b | 50.2 b | 33 b | 1 a |
| D | 405.6 b | 41.9 b | 277.9 a | 82 b | 195.9 a | 65.9 a | 50.2 a | 0.9 b |
| R | 431.4 a | 39.7 a | 219.6 b | 88.1 a | 131.5 b | 58.9 b | 31 b | 0.8 b |
| S | 371.8 b | 36 b | 343.3 a | 25.1 b | 318.2 a | 92.2 a | 87.2 a | 0.9 a |
Means with the same letter within each experiment do not differ significantly with 5% DMSH. RG= grain yield; PIG= individual grain weight; CHSAnt= water soluble carbohydrates at the time of anthesis; CHSMf= water soluble carbohydrates at physiological maturity; CHSTransl= total amount of soluble remobilized carbohydrates (difference between CHSAnt and CHSMf); ERCHS= total remobilization efficiency; CRG= contribution to grain yield; CHSG= water soluble carbohydrates present in the grain.
The variability observed among genotypes in terms of RG represented 62 and 48% in experiment 1 and 2, respectively of the total variation, while for the PIG it represented 50% in both experiments. For the accumulation of carbohydrates soluble in the stem in anthesis (CHSAnt) the genotypic effect observed 95% of the total variability, which coincides with evidence in the literature that mention that there is genetic variability for the accumulation of CHS in the wheat stem (Ruuska et al., 2006; Dreccer et al., 2009).
On the other hand, the variability between genotypes in terms of CHS remobilization and their contribution to grain yield represented 83 and 80% of the total variability, which indicates that the genetic improvement in wheat towards a greater contribution to the RG of the reserves stored could be possible to stabilize the grain yield in environments where stress due to biotic and abiotic factors is frequent (Ehdaie et al., 2008).
General response of environments
With the exception of CHSAnt, defoliation affected significantly (p< 0.05) all the variables studied. The RG, PIG, CHSMf and CHSG decreased on average 14, 9, 35 and 10%, respectively with respect to the non-defoliated treatment, while, CHSTransl, ERCHS and CRG increased significantly by 30, 31 and 52% respectively (Table 2) with defoliation. Drought on average decreased GR, PIG and CHSMf by 14, 9 and 71%, respectively with respect to irrigation, while in the variables CHSAnt, CHStransl, ERCHS, CRG, CHSG, significant increases were observed (56, 142, 56, 181 and 13%, respectively) with respect to the irrigated treatment (Table 2).
In both experiments there were significant increases in the values of CHTransl, ERCHS and CRG (30, 31 and 52%, respectively in defoliation, 142, 56 and 181%, respectively in drought). The contribution of stem reserves to grain yield increased, when the genotypes were subjected to different environments (defoliation and drought), with the contribution and remobilization of the assimilated during the filling of larger grain under drought conditions. Similar results report that the relative contribution of stem reserves to grain yield varies widely depending on environmental conditions and genotypes and their values range between 6 and 100% (Borrell et al., 1993).
Evidence in the literature mentions that the reduction in assimilation under post-anthesis humidity stress will induce a greater mobilization of stem reserves that will be used by the grain (Yang et al., 2000). Also, several studies have shown that water stress during grain filling promotes senescence of plants and increases the remobilization of stored carbon reserves in pre-anthesis towards the grain (Ahmadi et al., 2009). The major contribution to grain yield under stress conditions (defoliation and post-anthesis drought) was in line with a greater efficiency of remobilization of the CHS from the stem.
Grain yield was marginally reduced (14% on average) both by defoliation and by drought, which can be attributed to the fact that the extent of retranslocation of assimilates was much higher (59%), under that condition, acting as an element of important compensation to ensure grain filling (Singh and Singh, 2002; Ehdaie et al., 2008).
General effect of genotypes
A wide variability was observed among the 60 genotypes evaluated, in the variables RG, PIG, CHS in anthesis, translocated CHS and contribution of CHS to the grain yield (Figure 1 and 2). The RG ranged from 302 to 585 g m-2, the PIG was between 31 and 57 mg grain-1 (Figure 1).

Figure 1 Grain yield (a) and grain weight; and (b) for 60 wheat lines grown in Toluca, Mexico. Average data of environments (defoliated and unfrozen).

Figure 2 Carbohydrates soluble in anthesis (a) translocated soluble carbohydrates; (b) contribution to grain yield; and (c) for 60 wheat lines. Average data of environments (defoliated and defoliate).
While the CHs in anthesis were in the range of 125.6 to 545.1 g m-2, the CHS translocated between 11.2 to 464.9 g m-2 and the contribution of CHS to the yield ranged between 3 and 155.5% (Figure 2). A greater amount of CHS remobilized to the grains was in line with a greater contribution of these to the RG. In this sense, genotypes 8, 12, 13, 17, 19, 26 and 40, which translocated between 76 and 469.4 g m-2, in percentage terms also contributed more to the grain yield (76.4 to 155.5%) (Figure 2).
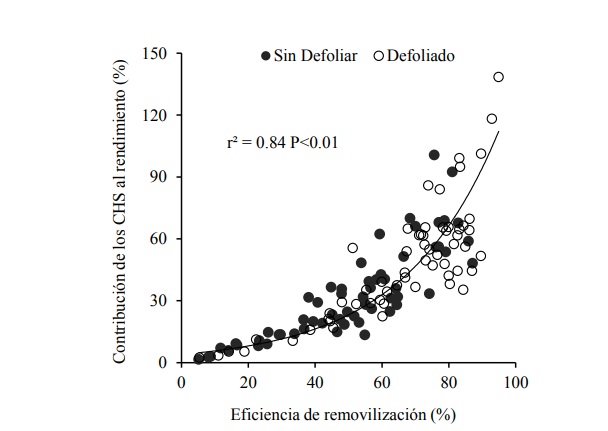
The great variability observed for grain yield, CHSAnt accumulation and contribution to grain yield shows that within elite wheat germplasm (CIMCOG) there is potential to select progenitors for breeding programs with a view to incorporating promising physiological characteristics (CHS) (Rebetzke et al., 2008) that allow to increase the yield under conditions of biotic stress (foliar diseases) (Serrago et al., 2011) or abiotic (drought) (Foulkes et al., 2007). A greater contribution of CHS to the grain yield was related to a greater efficiency of remobilization (Figure 3). In the defoliated plants there was a greater contribution of the CHS to the grain yield (range between 2.4 and 138.4%) with respect to the plants without defoliate (between 5.2 and 100.6%). In the same way, the efficiency of remobilization presented higher average values in defoliated plants (range between 5.4 to 94. 9%) in relation to non-defoliated plants (5.2 to 87.1%) (Figure 3).
Effect of water stress on CHS and grain yield
Water stress post-anthesis affected the RG, the PIG and the amount of CHS translocated from the stem to the grain in the different cultivars (Figure 4). The average RG in drought was 371.8 g m-2 and under irrigation 431.5 g m-2, which represents a 14% reduction with respect to the irrigation treatment. In this sense, the translocation of CHS under drought conditions was higher (318.2 g m-2) with respect to irrigation (131.5 g m-2), representing an increase of 141% with respect to the irrigation condition. Similar results of CHS remobilization were found by Zhang et al. (2012) when water availability represented only 50% of a favorable condition.

Figure 4 Relationship between grain yield (a) individual grain weight; and (b) soluble carbohydrates translocated from the stem to the spike, in 20 wheat lines grown in two environments. Vertical and horizontal lines within the figures represent the average of each environment.
In regions where the availability of water for grain filling is limiting, CHS play an important role in the definition of RG (Ehdaie et al., 2008), therefore, the identification of genotypes with the ability to accumulate and remobilize CHS from the stem is a priority in breeding programs (Dreccer et al., 2009). Based on the great variability observed in both environments, it was possible to identify genotypes (12 and 16) that did not show significant changes in the RG and in the PIG in both environments, mainly due to their ability to remobilize assimilated from the stem to the spike (Figure 4a, 4b).
The average PIG was marginally affected (9.5%) by the post-anthesis water deficit, while the CHS translocation under this condition represented 59% with respect to the control treatment (Figure 4b). The contribution of the CHS to the yield of grain in drought was higher (range between 31.0 to 155.6%) with respect to irrigation, thus also greater efficiency of remobilization was observed (82.9 to 95.7%) with respect to the cultivars grown under irrigated conditions, with values of 10.8 to 71.7% for the contribution to yield and from 23.3 to 80% in efficiency of remobilization (Figure 5a).

Figure 5 Relationship between CHS contribution to yield and efficiency of CHS (a) CHS remobilization in physiological maturity and stem dry weight (b), in 20 wheat lines grown under irrigated and drought conditions.
In drought it could be seen that for similar values of remobilization efficiency there was a wide variability in the contribution of the CHS to grain yield (Figure 5a). A higher CHS content per stem at physiological maturity was associated with a higher dry weight of them at that stage (Figure 5b). The content of carbohydrates stored in the stem for the genotypes grown under irrigation conditions was 0.27 to 1.23 g stem-1, while in the drought situation it was 0.06 to 0.28 g stem-1 (Figure 5b).
Conclusions
There was variability between the genotypes studied for grain yield and for accumulation and remobilization of CHS towards the grain. The amount of CHS remobilized, efficiency of remobilization and contribution to yield increased significantly (30, 31 and 52% in defoliation and 142, 56 and 181% in drought, respectively). There are genotypes (8, 12, 13, 17, 19, 26 and 40) that could have potential for use in breeding programs with a view to incorporating promising characteristics. The contribution of CHS to grain yield related to greater remobilization efficiency.
Acknowledgments
The author Cristóbal Valdés-Valdés, thanks the National Council of Science and Technology of Mexico (CONACYT) for the scholarship that allowed him to carry out his PhD studies in Agricultural Sciences and Natural Resources. The present work was part of the project ‘trial of adaptation of diverse wheat to contrasting environments in Mexico’ with code 3350/2012E.
REFERENCES
Ahmadi, A.; Joudi, M. and Janmohammadi, M. 2009. Late defoliation and wheat yield: little evidence of post-anthesis source limitation. Field Crops Res. 113(1):90-93. [ Links ]
Blum, A. 1998. Improving wheat grain filling under stress by stem reserve mobilisation. Euphytica. 100(1-3):77-83. [ Links ]
Borrell, A.; Incoll, L. D. and Dalling, M. J. 1993. The influence of the Rht1 and Rht2 alleles on the deposition and use of stem reserve in wheat. Ann. Bot. 71(4):317-326. [ Links ]
Davidson, D. J. and Chevalier, P. M. 1992. Storage and remobilization of water-soluble carbohydrates in stems of spring wheat. Crop Sci. 32(1):186-190. [ Links ]
Dreccer, M. F.; van Herwaarden, A. F. and Chapman, S. C. 2009. Grain number and grain weight in wheat lines contrasting for stem water soluble carbohydrate concentration. Field Crops Res. 112(1):43-54. [ Links ]
Ehdaie, B.; Alloush, G. A. and Waines, J. G. 2008. Genotypic variation in linear rate of grain growth and contribution of stem reserves to grain yield in wheat. Field Crops Res. 106(1):34-43. [ Links ]
Ehdaie, B.; Alloush, G. A.; Madore, M. A. and Waines, J. G. 2006. Genotypic variation for stem reserves and mobilization in wheat. II. Postanthesis changes in internode water-soluble carbohydrates. Crop Sci. 46(5):2093-2103. [ Links ]
Foulkes, M. J.; Slafer, G. A.; Davies, W. J.; Berry, P. M.; Sylvester-Bradley, R.; Martre, P.; Calderini, D. F.; Griffiths, S. and Reynolds, M. P. 2011. Raising yield potential of wheat. III. Optimizing partitioning to grain while maintaining lodging resistance. J. Exp. Bot. 62(2):469-486. [ Links ]
Foulkes, M. J.; Sylvester-Bradley, R.; Weightman, R. and Snape, J. W. 2007. Identifying physiological traits associated with improved drought resistance in winter wheat. Field Crops Res. 103(1):11-24. [ Links ]
Galicia, L.; Nurit, E.; Rosales, A. and Palacios-Rojas, N. 2009. Laboratory protocols. Soluble sugars determination using anthrone reagent. Centro Internacional de Maíz y Trigo (CIMMYT). El Batán, Estado de México. 22-25 pp. [ Links ]
Hortelano, S. R. R.; Villaseñor, M. H. E.; Martínez, C. E.; Rodríguez, G. M. F.; Espitia, R. E. y Mariscal, A. L. A. 2013. Estabilidad de variedades de trigo recomendadas para siembras de temporal en los Valles Altos de la Mesa Central. Rev. Mex. Cienc. Agríc. 4(5):713-725. [ Links ]
INEGI. 2009. Instituto Nacional de Estadística Geografía e Informática. Producción Agrícola Estatal. México. http://www.inegi.org.mx/est/contenidos/proyectos/agro/ca2007 /resultados-agricola/default.aspx. [ Links ]
Kobata, T.; Palta, J. A. and Turner, N. C. 1992. Rate of development of post-anthesis water deficits and grain filling of spring wheat. Crop Sci. 32(5):1238-1242. [ Links ]
Ledesma-Ramírez, L.; Solís-Moya, E.; Suaste-Franco, M. P.; Rodríguez-Caracheo, J. F. y de la Cruz-Gonzalez, M. L. 2012. Análisis gge biplot del rendimiento de trigo (Triticum spp.) con riego normal y restringido en el Bajío, México. Agrociencia. 46(1):119-131. [ Links ]
Littell, R. C.; Milliken, G. A.; Stroup, W. W. and Wolfinger, R. D. 1996. SAS® system for mixed models. Cary, NC. SAS Institute Inc. 633 p. [ Links ]
Mariotti, M.; Arduini, I. and Lulli, L. 2003. Traslocazione della biomassa nel frumento duro durante il grain-filling. In: Proceedings of the XXXV Congress of the Italian Society of Agronomy, Napoli. Mori, M. and Fagnano, M. (Eds.) 16-18 September. Imago Media, Caserta, Italy. 339-340 pp. [ Links ]
Palaniswamy, U. R. and Palaniswamy, K. M. 2006. Handbook of statistics for teaching and research in plant and crop science. The Harworth Press, Inc., New York. 624 p. [ Links ]
Pask, A. J. D.; Pietragalla, J.; Mullan, D. M.; Chavez-Dulanto, P. N. and Reynolds, M. P. 2013. Physiological breeding II. A field guide to wheat phenotyping. CIMMYT. Mexico, DF. 133 p. [ Links ]
Przulj, N. and Momcilovic, V. 2001. Genetic variation for dry matter and nitrogen accumulation and translocation in two-rowed spring barley I. Dry matter translocation. Eur. J. Agron. 15(4):241-254. [ Links ]
Rebetzke, G. J.; Van Herwaarden, A. F.; Jenkins, C.; Weiss, M.; Lewis, D.; Ruuska, S.; Tabe, L.; Fettell, N. A. and Richards, R. A. 2008. Quantitative trait loci for water-soluble carbohydrates and associations with agronomic traits in wheat. Austr. J. Agric. Res. 59(10):891-905. [ Links ]
Reynolds, M. P.; Bonnett, D.; Chapman, S. C.; Furbank, R. T.; Mans, Y.; Mather, D. E. and Parry, M. A. 2011. Raising yield potential of wheat. I. Overview of a consortium approach and breeding strategies. J. Exp. Bot. 62(2):439-452. [ Links ]
Ruuska, S. A.; Rebetzke, G. J.; Van Herwaarden, A. F.; Richards, R. A.; Fettell, N. A.; Tabe, L. and Jenkins, C. L. D. 2006. Genotypic variation in water-soluble carbohydrate accumulation in wheat. Functional Plant Biol. 33(9):799-809. [ Links ]
Sas Institute. 2002. SAS/STAT 9.0 User’s guide. SAS Inst., Cary, NC. 633 p. [ Links ]
Serrago, R. A.; Carretero, R.; Bancalb, M. O. and Miralles, D. J. 2011. Grain weight response to foliar diseases control in wheat (Triticum aestivum L.). Field Crops Res. 120(3):352-359. [ Links ]
Shakiba, M. R.; Ehdaie, B.; Madore, M. A. and Waines, J. G. 1996. Contribution of internode reserves to grain yield in a tall and semidwarf spring wheat. J. Genet. Breed. 50(1):91-100. [ Links ]
Shearman, V. J.; Sylvester-Bradley, R.; Scott, R. K. and Foulkes, M. J. 2005. Physiological processes associated with wheat yield progress in the UK. Crop Sci. 45(1):175-185. [ Links ]
Singh, D. and Singh, D. 2002. Effect of leaf blade and awn on grain yield on rainfed wheat (Triticum aestivum L.) at different stages of spike development. Ind. J. Agric. Sci. 72(7):468-471. [ Links ]
Tahir, I. S. A. and Nakata, N. 2005. Remobilization of nitrogen and carbohydrate from stems of bread wheat in response to heat stress during grain filling. J. Agron. Crop Sci. 191(2):106-115. [ Links ]
USDA. 2000. Natural Resources Conservation Service. World Soil Resources Natural Resources Conservation Service (http://www.nhq.usda.gov/WSR/). United States Department of Agriculture. Washington, DC. [ Links ]
Van Herwaarden, A. F. and Richards, R. A. 2002. Water soluble carbohydrate accumulation in the stems is related to breeding progress in Australian wheats. Plant breeding for the 11th millennium. In: McComb, J. A. (Ed.). Proceedings of the 12th Plant Breeding Conference, Perth, 15-20 September. 878-882 pp. [ Links ]
Van Herwaarden, A. F.; Angus, J. F.; Richards, R. A. and Farquhar, G. D. 1998b. Haying-off, the negative grain yield response to nitrogen fertiliser. II. Carbohydrate and protein dynamics. Aust. J. Agric. Res. 49(7):1083-1093. [ Links ]
Van Herwaarden, A. F.; Richards, R. A.; Farquhar, G. D. and Angus, J. F. 1998a. Haying-off, the negative grain yield response to nitrogen fertilizer. I. Biomass, grain yield and water use. Aust. J. Agric. Res. 49(7):1067-1081. [ Links ]
Van Herwaarden, A.; Richards, R. and Angus, J. 2006. Water-soluble carbohydrates and yield in wheat. The Australian Society of Agronomy. Proceedings of 13th Agronomy Conference (http://www.regional.org.au/au/asa/2003/c/6/vanherwaarden.htm). [ Links ]
Yang, D. L.; Jing, R. L.; Chang, X. P. and Li, W. 2007. Identification of quantitative trait loci and environmental interactions for accumulation and remobilization of water-soluble carbohydrates in wheat (Triticum aestivum L.) stems. Genetics. 176(1):571-584. [ Links ]
Yang, J. C.; Zhang, J. H.; Huang, Z. L.; Zhu, Q. S. and Wang, L. 2000. Remobilization of carbon reserves is improved by controlled soil-drying during grain filling of wheat. Crop Sci. 40(6):1645-1655. [ Links ]
Yemm, E. W. and Willis, A. J. 1954. The estimation of carbohydrates in plant extracts by anthrone. Biochem. J. 57(3):508-514. [ Links ]
Zadoks, J. C.; Chang, T. T. and Konzak, C. F. 1974. A decimal code for the growth stages of cereals. Weed Res. 14(6):415-421. [ Links ]
Zhang, H.; Turner, N. C. and Poole, M. L. 2012. Increasing the harvest index of wheat in the high rainfall zones of southern Australia. Field Crops Res. 129(2):111-123. [ Links ]
Zhang, Y. P.; Zhang, Y. H.; Xue, Q. W. and Wang, Z. M. 2013. Remobilization of water-soluble carbohydrates in non-leaf organs and contribution to grain yield in winter wheat under reduced irrigation. International J. Plant Production. 7(1):97-116. [ Links ]
Received: February 01, 2019; Accepted: May 01, 2019
 Este es un artículo publicado en acceso abierto bajo una licencia
Creative Commons
Este es un artículo publicado en acceso abierto bajo una licencia
Creative Commons
