











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
La fusariosis de la espiga, también conocida como roña de la espiga o Fusarium head blight (FHB, por sus siglas en inglés), es una enfermedad importante en los cereales de grano pequeño como cebada (Hordeun vulgare L.). Esta enfermedad, ocasionada por algunas especies de Fusarium entre las que destaca F. graminearum (Schwabe) (Salas et al., 1999), ataca principalmente en la etapa de floración a grano lechoso afectando el rendimiento y la calidad sanitaria del grano. Ireta y Gilchrist (1994) reportaron pérdidas de 17% en rendimiento en trigo en la zona productora de Valles Altos de México; sin embargo, se han registrado pérdidas de hasta 50% en otras partes del mundo (Windels, 2000). El consumo de granos o productos de cebada maltera contaminados afecta la salud de las personas y de los animales debido a la producción de compuestos tóxicos como el Deoxinivalenol (DON), del grupo de los tricotecenos por Fusarium (Bezerra et al., 2014).
Por las implicaciones a la salud humana y animal de la toxina DON, también conocida como vomitoxina por ocasionar vómito en las personas y rechazo del alimento en animales (Zain, 2011), la industria alimentaria ha implementado límites de tolerancia a la concentración de DON en el grano contaminado. Al respecto, la agencia de drogas y alimentación norteamericana (FDA, por sus siglas en inglés) establece los siguientes límites en granos y subproductos de granos de trigo: 1 ppm para productos de trigo (harina, salvado y germen) destinados para consumo humano, 5 ppm para productos de grano para alimentos para cerdos, de 10 ppm para granos y subproductos de grano para la elaboración de alimentos para ganado bovino y pollos y de 30 ppm en granos para destilería (FDA, 2010).
Sin embargo, la industria cervecera estableció un límite de 0.5 ppm para la producción de malta (Windels, 2000). Lo anterior indica que la fusariosis es una enfermedad que limita el rendimiento, afecta la calidad sanitaria del grano y repercute negativamente en los sectores agropecuario, alimenticio e industrial. Por otra parte, Malihipour y Gilbert (2012); De la Torre-Hernández et al. (2014) reportan que la toxina DON es un factor de virulencia y la asocian con la capacidad de las especies de Fusarium que la producen de causar enfermedad en su hospedante. De Wolf et al. (2003); Bondalapati et al. (2012) indican que condiciones de humedad relativa alrededor de 90 % y temperatura de 15 a 30 °C durante el periodo de antesis son favorables para el desarrollo de la fusariosis y la acumulación de DON.
El Programa de Cebada del Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP) del Campo Experimental Valle de México (CEVAMEX) tiene trabajos de cruzas para el mejoramiento de cebada maltera contando actualmente con 131 líneas avanzadas con tolerancia a roya amarilla (Puccinia striiformis f. sp. hordei West), a roya de la hoja (P. hordei Otth), mancha reticulada (Drechslera teres Sacc), tizón foliar (Bipolaris sorokiniana) y escaldadura de la hoja (Rhyncosporium secalis (Oud) J. J. Davis), enfermedades consideradas como las más frecuentes en las zonas productoras de los Valles Altos de México (Gilchrist-Saavedra, 2000; Zamora et al., 2008).
Es importante mencionar que algunas líneas avanzadas de cebada tienen como progenitores a Gobernadora, Chevron y CI14064, las cuales se utilizan como fuentes de resistencia por desarrollar bajos niveles de fusariosis de la espiga (Zhu et al., 1999; McCallum et al., 2004; Marchand et al., 2008). Considerando que entre las medidas recomendadas de control de la fusariosis de la espiga se encuentra el uso de variedades resistentes para disminuir la incidencia de Fusarium en el grano (Horevaj et al., 2011; McMullen et al., 2012), los objetivos de este trabajo fueron: evaluar la resistencia a la fusariosis de las 131 líneas avanzadas de cebada maltera del Programa de Cebada del INIFAP-CEVAMEX en condiciones de invernadero; y, cuantificar los niveles de producción de toxina DON en semilla de cebada maltera cosechada de espigas inoculadas con F. graminearum.
La información generada en este trabajo es relevante y puede utilizarse como punto de partida en un programa de cruzamientos de cebada maltera en el que incluya, en el conjunto de enfermedades, a la fusariosis de la espiga y a la acumulación de toxina DON en grano.
Materiales y métodos
Este estudio se llevó a cabo durante el ciclo invierno-primavera de los años 2015, 2016 y 2017, en el Colegio de Postgraduados, Campus Montecillo, Estado de México. El campus está a una altitud de 2240 msnm, presenta clima templado subhúmedo, con precipitación media anual de 700 mm, con régimen de lluvias en verano. Las plantas se establecieron en un invernadero de 437.5 m2 construido con plástico transparente y piso de tierra sin condiciones ambientales controladas. Durante el periodo de inoculación de las plantas, en el invernadero se registró una temperatura promedio de 20 °C (con mínimo de 9 °C y máximo de 35 °C); HR de 46.6 % (con mínimo de 26.5 % y máximo 68.9 %), con un sensor automático HOBO H8 (Onset Computer corporation, USA).
Germoplasma y establecimiento de plantas
La semilla de 131 líneas avanzadas de cebada maltera, organizadas en sublotes: 03 (30 líneas), 02 (30 líneas), 01 (25 líneas) y Elite (16 líneas), de seis hileras de grano; y el sublote 02A (30 líneas), de dos hileras, fue proporcionada por el Programa de Cebada del INIFAP-CEVAMEX. Se destaca que la selección de las líneas fue a partir del sublote 03 seguido del 02A, 02, 01 y finalizando con Elite. Es decir, los sublotes 03 y Elite representan los sublotes de menor y mayor ciclo de selección, respectivamente. Como testigos se incluyó semilla de las variedades: Seebe, CDC Copeland, Xena, AC Ranger, Kasota y AC Lacombe, proporcionada por el Centro Internacional de Mejoramiento de Maíz y Trigo (CIMMYT), El Batán, así como de las variedades: Gobernadora, Adabella, Esmeralda y Shenmai 1, proporcionada por el INIFAP-CEVAMEX (Cuadro 1).
Cuadro 1 Relación de variedades de cebada utilizadas como testigo y características de número de hileras de grano por espiga y resistencia a fusariosis de la espiga, según la literatura .
Variedad |
Núm. de hileras de grano por espiga |
Resistencia a la enfermedad |
Resistente (R) ó Moderadamente resistente (MR) | ||
Gobernadora |
2 |
Variedad R, con el tiempo se ha comportado como MR (McCallum et al., 2004; Marchand et al., 2008). |
Seebe |
2 |
Registrada como MR, en ocasiones se comporta como R (Helm et al., 1996; He et al., 2015). |
Moderadamente resistente (MR) ó moderadamente susceptible (MS) | ||
Adabella |
6 |
Variedad MR con tolerancia (Zamora et al., 2008). |
Shenmai 1 |
2 |
Nombre utilizado en China para referirse a una población segregante de Gobernadora (F4) (Vivar et al., 1997). |
CDC Copeland |
2 |
Se comporta como MR y MS (He et al., 2015). |
Xena |
2 |
Se comporta como MR y MS (He et al., 2015). |
MS ó Susceptible (S) | ||
Esmeralda |
6 |
Variedad R pero con el tiempo se ha comportado como S (Zamora et al., 1997). |
AC Lacombe |
6 |
Variedad considerada entre MS y S (Kibite, 1993; He et al., 2015). |
AC Ranger |
6 |
Variedad considerada entre MS y S (Therrien, 2002). |
Kasota |
6 |
Variedad S (Helm et al., 1996). |
La semilla se sembró por duplicado en bolsas de plástico (25 x 25 cm) con sustrato peat moss (Promix®) esterilizado con vapor. Por cada línea y variedad testigo se depositaron 10 semillas por bolsa. Al inicio del cultivo se dieron tres riegos por gravedad con agua de pozo por semana; después de la floración, las plantas se regaron diariamente por aspersión. Para la fertilización de las plantas se utilizó triple 16 (16-16-16), 18-46-00 y urea (Yara®) aplicando la fórmula 100-46-30. El fósforo y el potasio se aplicaron en su totalidad a los ocho días después de la siembra; el nitrógeno solo el 50% y el restante, a los 60 días después de la siembra. Durante el ciclo de 2017 se colocó una malla sombra de 75% para generar condiciones frescas en el invernadero.
Cepas y preparación de inóculo
En el Cuadro 2 se presenta la relación de cepas de F. graminearum empleadas para las inoculaciones en invernadero durante los tres años de estudio. Estas cepas son monospóricas aisladas de granos infectados con F. graminearum. En 2015 se utilizó la cepa CIMFU1189 aislada de granos de trigo colectados en Toluca, Edo. de México; mientras que en 2016, la mezcla de las cepas CIMFU1308, CIMFU1310, CIMFU1312 y CIMFU1313 aisladas de granos de trigo. Todas estas cepas fueron proporcionadas por el Laboratorio de Fitopatología del CIMMYT y se utilizaron por su alta capacidad para producir DON (Cuadro 2).
Cuadro 2 Relación de cepas de F. graminearum empleadas para las inoculaciones en invernadero.
Año |
Fuente: clave cepa |
Origen Huésped |
Producción toxina DON (ppm kg-1) |
Inoculación† de espigas |
2015 |
CIMMYT: CIMFU1189 |
Trigo |
14.5 |
Por aspersión |
2016 |
CIMMYT: CIMFU1313, CIMFU1310, CIMFU1308, CIMFU1312 |
Trigo |
16.1, 17, 18.1 y 18.8, respectivamente (con promedio de 17.5) |
Por punción |
2017 |
Este estudio: SL10R10, CU6R30, SL3R24, SL3R23 y CU1R30 |
Cebada |
3.4, 3.6, 5, 9.2 y 17, respectivamente (con promedio de 7.6) |
Por aspersión |
†= en 2015 con 100 000 conidios mL-1; en 2016 y 2017 con 70 000 conidios mL-1.
En tanto que, en 2017 se aplicó la mezcla de las cepas CU1R30, SL3R23, SL3R24, SL10R10 y CU6R30 aisladas de granos de cebada maltera colectados en Cuyuaco (CU), Puebla y Santa Lucía (SL), Edo de México. Estas últimas cepas se aislaron, identificaron y caracterizaron para este estudio (datos no mostrados) y se seleccionaron también por su alta capacidad para producir DON (Cuadro 2). En 2016 el Laboratorio de Fitopatología de Trigo del CIMMYT proporcionó la mezcla con las cuatro cepas para la inoculación de las plantas; mientras que en 2015 y 2017 las mezclas se prepararon en el Laboratorio de Enfermedades en Postcosecha, del Posgrado de Fitopatología, del Colegio de Posgraduados.
Para esto un fragmento de papel filtro WhatmanTM1 estéril con crecimiento del hongo, almacenado por 5 meses a 4°C, se colocó en una caja Petri con PDA y se incubó a temperatura ambiente. Después de 10 días se cortaron círculos de medio con crecimiento del hongo (4 mm) y se depositaron en matraces de 250 mL con 50 mL de medio líquido de frijol chino (Vigna radiata (L.) Wilezek) estéril para la producción de conidios (Bai, 1996). Los matraces inoculados se mantuvieron en agitación (Agitador Lab-Line®) con luz blanca continua (lámpara Fulgore®, 250 V, 32 W) a temperatura ambiente por 7 días. La suspensión obtenida se filtró a través de una gaza estéril y la concentración de conidios se estimó en un hematocitómetro. Todas las suspensiones se mantuvieron a -10 oC hasta su utilización.
Inoculación de espigas
En 2015 las espigas se inocularon en la etapa de grano lechoso por aspersión con un inóculo de 100 000 conidios mL-1; mientras que en 2016 por punción con 70 000 conidios mL-1, cuando las plantas empezaron a emitir las espigas. Para esto, una gota de 10 µL se colocó en la espiguilla central de la espiga con una jeringa hipodérmica (Engle et al., 2003). En 2017 la inoculación fue al inicio del espigamiento por aspersión con un inóculo de 70 000 conidios mL-1 (Cuadro 2).
A diferencia de los dos ciclos anteriores, en este ciclo las espigas se inocularon nuevamente a los cuatro días después de la primera inoculación por el mismo método y con la misma cantidad de inóculo. Inmediatamente después de cada inoculación, las espigas se cubrieron por 48 h con una bolsa de plástico trasparente, previamente asperjada con agua estéril. El piso de tierra del invernadero se regó diariamente para tener un ambiente de alta humedad relativa alrededor de las plantas.
Evaluación de incidencia, severidad e índice de fusariosis
Por línea se evaluaron de 1 a 10 espigas seleccionadas al azar a los 21 días después de la inoculación por aspersión; y a los 7, 14, 21 y 28 días después de la inoculación por punción. En la inoculación por aspersión se evaluó la resistencia Tipo I (resistencia a la infección inicial) mientras que en la inoculación por punción la resistencia Tipo II (resistencia a la dispersión a través del raquis) (McMullen et al., 2012). Por lo anterior las variables evaluadas fueron.
Porcentaje de incidencia (número de espigas con síntomas x 100/ total de espigas inoculadas); porcentaje de severidad de la enfermedad (número de granos con síntomas x 100/ total de granos por espiga) con una escala de 1 a 9 donde 1) <5%, 2) 5-17%, 3) 18-30%, 4) 31-43%, 5) 44-56%, 6) 57-69%, 7) 70-82%, 8) 83-95% y 9) >95% de granos con fusariosis. También se evaluó el índice de fusariosis (IFHB por sus siglas en inglés) (% incidencia x % severidad/ 100 (Groth et al., 1999).
Análisis estadístico
Los valores de las variables porcentaje de incidencia, porcentaje de severidad e índice IFHB mostraron una distribución no simétrica en una prueba de bondad de ajuste por lo que los datos se analizaron mediante un modelo lineal generalizado (Proc GLIMMIX) con distribución Poisson y función de enlace (link) log con el software SAS (Statistical Analysis Software) Versión 9.4 para Windows.
Análisis de toxina DON
Semilla de 56 líneas de los sublotes Elite (10 líneas), 01 (8 líneas), 02 (11 líneas), 02A (14 líneas) y 03 (13 líneas), del ciclo 2017, se analizaron con los reactivos y protocolo de RIDASCREEN®FAST DON (r-Biopharm) [inmunoensayo enzimático (ELISA)]. Con excepción de las líneas 15, 33 y 131 de los sublotes Elite, 01 y 03, respectivamente, todas esas líneas (53 líneas) se comportaron como resistentes a la fusariosis; las tres líneas restantes, como susceptibles. Así mismo se analizó semilla de los testigos Seebe, Shenmai 1 y Kasota como referencia de testigos R, MR y S, respectivamente. Toda la semilla de las líneas y la de los testigos se cosecharon en madurez fisiológica de espigas inoculadas con la mezcla de cepas SL10R10, CU6R30, SL3R24, SL3R23 y CU1R30, durante dicho ciclo.
La semilla de cada línea y testigo se molió por separado en un molino Iwatani 200W (AC 100 V, 50-60 HZ Pushon. Japón) por 40 s. La harina se recuperó en tubos Eppendorf de 50 mL con tapa rosca y se mezcló para homogeneizar. De cada material se tomó 2 g de harina y se depositó por separado en un tubo Eppendorf de 50 mL para su análisis con algunas modificaciones propuestas por CIMMYT. A esta muestra se le agregó 40 mL de agua y se agitó a 400 rpm por 3 min (Agitador Lab-line, modelo 4626). Un mililitro de la suspensión obtenida se depositó en un tubo Eppendorf de 2 mL para centrifugarse a 13,000 rpm (MIKRO 220R) por 15 min. Del sobrenadante obtenido se tomaron 200 µL y se almacenaron en refrigeración para su posterior análisis.
Las reacciones se establecieron por duplicado en placas con 96 pocillos, sensibilizadas con anticuerpos de captura contra anticuerpos anti-deoxinivalenol. De cada una de las 59 muestras y de cada estándar (0, 0.222, 0.666, 2 y 6 ppm de deoxinivalenol en agua) se tomaron 50 µL y se depositaron en la placa de acuerdo a un diagrama de carga preestablecido. La lectura de la placa se hizo en un lector de ELISA (LABOMED INC. EMR-500) a 450 nm y la visualización de los datos con el programa RIDAWIN.EXE Ver. 1.77.
Resultados y discusión
Respuesta de las líneas a la infección y a la dispersión de la enfermedad
En este estudio se encontró variación en la respuesta tanto entre las 131 líneas de cebada maltera como también entre las 10 líneas testigo (variedades) a la inoculación por punción y aspersión con las cepas de F. graminearum. Por lo anterior para clasificar la respuesta de resistente (R), moderadamente resistente (MR) y susceptible (S) se tomó como referencia el valor esperado de las variedades testigo (R, MR y S) de acuerdo al ciclo evaluado (Cuadro 3).
Cuadro 3 Índice de fusariosis (IFHB) registrado en las variedades testigo de cebada en respuesta a la inoculación con cepas de F. graminearum durante tres años de evaluación en invernadero.
Variedad |
2015 |
2016 |
2017 |
||||||||
Resistente o moderadamente resistente | |||||||||||
Gobernadora |
4.5 |
1.4 |
3 |
3.2 |
2.5 |
2.8 |
|||||
Seebe |
0 |
2.8 |
1.4 |
2.1 |
0.8 |
1.5 |
0.5 |
1.8 |
1.2 |
||
Moderadamente resistentes o moderadamente susceptible | |||||||||||
Adabella |
9.4 |
6.8 |
8.1 |
||||||||
Shenmai 1 |
3.5 |
7.3 |
5.4 |
12.1 |
3.6 |
7.8 |
|||||
CDC-Copeland |
0 |
11.8 |
5.9 |
0.9 |
1.8 |
1.3 |
22.1 |
0.9 |
11.5 |
||
Xena |
0 |
26.9 |
13.5 |
2.4 |
1 |
1.7 |
8 |
2.9 |
5.4 |
||
Moderadamente susceptible o susceptible | |||||||||||
Esmeralda |
4 |
2.1 |
3.1 |
||||||||
AC Lacombe |
8.9 |
8.8 |
8.8 |
4.4 |
2.3 |
3.3 |
14.9 |
0.9 |
7.9 |
||
AC Ranger |
4.4 |
2.2 |
3.3 |
1.7 |
0.3 |
1 |
17.1 |
1.4 |
9.3 |
||
Kasota |
6.7 |
13.3 |
10 |
2.8 |
5.9 |
4.4 |
12.4 |
24.9 |
18.6 |
||
†= 2015 por aspersión con cepa CIMFU1189. Por punción en 2016 con mezcla de cepas CIMFU1308, CIMFU1310, CIMFU1312 y CIMFU 1313; y aspersión en 2017 con mezcla de cepas CU1R30, SL3R23, SL3R24, SL10R10 y CU6R30. R1= repetición 1; R2= repetición 2. Ẋ= promedio.
La variación entre repeticiones (plantas) de una misma variedad (Cuadro 3) puede estar relacionada con la etapa fenológica de la planta al momento de la inoculación y con el microambiente generado cuando la planta se cubrió con la bolsa de plástico para favorecer la enfermedad. Es posible que la diferencia en valores entre las repeticiones de una variedad pueda ser resultado de un escape a la enfermedad y no un reflejo de la resistencia de la planta. Así, durante el ciclo 2015 se tomó como referencia el valor de IFHB registrado en Seebe (1.4, R), Adabella (8.1, MR) y Xena (13.5, S), quedando las 131 líneas clasificadas como: 82 líneas (63%) R; 34 líneas (26%) como MR; y 15 líneas (11%) como S; por haber registrado un valor de 0 a 5, de 6 a 10, y ≥11 de IFHB, respectivamente. Todos los sublotes registraron líneas con los tres tipos de respuesta.
En este ciclo las líneas del sublote 02A (líneas de dos hileras) se encontraron muy precoces y entre ellas destacó la línea 94 por haber registrado el mayor índice IFHB (29), seguida de la línea 6 (23) del sublote Elite y de la línea 33 (18) del sublote 01. Estas tres líneas registraron nivel 3 (18-30 %) en la escala de severidad (Chrpová et al., 2011) e incidencia de 0 a 100%. En el Cuadro 4 se ilustra la clasificación de 56 líneas por tipo de respuesta R, MR o S para el ciclo 2015.
Cuadro 4 Clasificación de las líneas de cebada en resistentes (R), moderadamente resistentes (MR) y susceptibles (S) de acuerdo a la respuesta de IFHB así como a la acumulación de DON.
|
Sublote Núm. de línea |
Respuesta IFHB |
DON (ppm kg-1)‡ ¥ |
|||||
2015 |
2016 |
2017 |
0 - 4 (R) |
5 - 11 (MR) |
≥12 (S) |
||
Elite 1 |
S |
MR |
R |
15.6 - 17.4 |
|||
3 |
MR |
MR |
R |
1.7 - 2 |
|||
4 |
R |
MR |
R |
3 - 3.1 |
|||
5 |
MR |
MR |
R |
13.4 - 20.3 |
|||
7 |
MR |
MR |
R |
10 - 14.8 |
|||
8 |
R |
MR |
R |
5.9 - 6 |
|||
9 |
MR |
MR |
R |
3.5 - 4.4 |
|||
12 |
MR |
MR |
R |
2.7 - 3.4 |
|||
14 |
R |
S |
R |
17.7 - 18 |
|||
15 |
S |
MR |
S |
17.1 - 19.1 |
|||
01 18 |
R |
MR |
R |
7.7 - 10.8 |
|||
21 |
MR |
R |
MR |
11.3 - 16.2 |
|||
23 |
S |
MR |
R |
21 - 30.4 |
|||
28 |
R |
MR |
R |
0.11 - 0.13 |
|||
33 |
S |
MR |
S |
16.7 - 19.7 |
|||
30 |
R |
R |
R |
0.7 - 0.8 |
|||
36 |
MR |
MR |
R |
13.9 - 19.4 |
|||
41 |
R |
R |
R |
5.1 - 5.1 |
|||
02 42 |
R |
S |
R |
2.1 - 2 |
|||
46 |
R |
S |
R |
16.3 - 20 |
|||
47 |
R |
R |
R |
0.6 - 0.8 |
|||
48 |
R |
MR |
R |
3.2 - 3.3 |
|||
53 |
MR |
MR |
R |
6.4 - 6.9 |
|||
56 |
R |
S |
R |
1.2 - 1.3 |
|||
58 |
R |
S |
R |
5.1 - 5.2 |
|||
63 |
R |
R |
R |
10.5 - 11.1 |
|||
65 |
R |
R |
R |
0.01 - 0.04 |
|||
67 |
R |
S |
R |
11.7 - 13.6 |
|||
68 |
S |
R |
R |
11.3 - 12.7 |
|||
02A 72 |
R |
MR |
R |
4.8 - 7.2 |
|||
73 |
R |
R |
R |
0.01 - 0.02 |
|||
75 |
R |
S |
R |
5.4 - 50 |
|||
79 |
R |
MR |
R |
8.4 - 10.1 |
|||
81 |
R |
S |
R |
6.8 - 8.9 |
|||
82 |
R |
S |
R |
24 - 22.8 |
|||
85 |
S |
MR |
R |
3.6 - 4.2 |
|||
87 |
R |
S |
R |
0.5 - 0.7 |
|||
90 |
MR |
S |
R |
0 - 0 |
|||
91 |
R |
R |
R |
0.1 - 0.1 |
|||
93 |
R |
S |
R |
2.7 - 4.2 |
|||
96 |
R |
MR |
R |
0 - 0 |
|||
98 |
R |
S |
R |
5.4 - 6 |
|||
100 |
S |
S |
R |
1.7 - 5.2 |
|||
03 103 |
R |
S |
R |
0.8 - 1.3 |
|||
106 |
R |
MR |
R |
7.7 - 9.3 |
|||
109 |
R |
R |
R |
9 - 11.8 |
|||
112 |
R |
MR |
R |
0.09 - 0.12 |
|||
114 |
MR |
MR |
R |
0. 8 - 0.8 |
|||
116 |
R |
MR |
R |
0.4 - 0.7 |
|||
119 |
MR |
R |
R |
19.1 - 22.4 |
|||
122 |
R |
MR |
R |
14.5 - 14.6 |
|||
123 |
R |
R |
R |
1.5 - 1.7 |
|||
124 |
R |
R |
R |
8.1 - 9.8 |
|||
125 |
R |
S |
R |
1 - 1.5 |
|||
127 |
MR |
S |
R |
2 - 4.8 |
|||
131 |
R |
MR |
S |
12.4 - 13.9 |
|||
†= inoculación por aspersión en 2015 y 2017 y por punción en 2016. ‡= referencia: Seebe (R, 3.4 -4), Shenmai 1 (MR, 5.8 - 10.1) y Kasota (S, 22.0-22.8). ¥= límite detección= 0.2 ppm kg-1; límite cuantificación= 0.36 ppm kg-1 para avena.
Durante el ciclo 2016 se tomó como referencia el índice IFHB registrado por los testigos Seebe (1.5, R), Gobernadora (3, MR) y Kasota (4.4, S) (Cuadro 3). En general se registró incidencia de 25 a 100%, severidad de nivel 2 (de 1 a 12%) e índice de IFHB de 0 a 10. En la mayoría de las espigas, de todas las líneas, la necrosis se desarrolló solo en la espiguilla en la que se depositó el inóculo; es decir, la dispersión de la enfermedad a través del raquis fue baja. Sin embargo, en las líneas 84 y 92 del sublote 02A la dispersión en el raquis fue más allá del punto de inoculación, aunque el síntoma de fusariosis no se observó en los granos cercanos.
Es posible que el hongo haya quedado confinado en el raquis de la espiga de estas líneas. De acuerdo a los resultados obtenidos: 18 líneas (13.7 %) se comportaron como R; 69 líneas (52.7 %) como MR y 44 líneas (33.6 %) como S por haber registrado un valor de 0 a 1, de 2 a 3, y ≥4 de IFHB, respectivamente. De las líneas resistentes sobresalieron las líneas 63 del sublote 02 y 73 del sublote 02A, por no registrar síntomas de enfermedad. McCallum y Tekauz (2002) y Geddes et al. (2008) mencionan que la cebada muestra de manera natural resistencia a la dispersión de la fusariosis a través del raquis; mientras que Jansen et al. (2005) indican que la infección por F. graminearum se ve limitada por la inhibición del crecimiento de las hifas en el raquis de este cultivo.
Los datos anteriores indican que las plantas inoculadas por aspersión (ciclo 2015) registraron mayor severidad (nivel 3) que las inoculadas por punción (nivel 2, ciclo 2016). McCallum y Tekauz (2002) mencionan que cuando la dispersión en la espiga es baja también lo es la severidad de la enfermedad. En el Cuadro 4 se ilustra la clasificación de 56 líneas por tipo de respuesta R, MR o S para el ciclo 2016. En el ciclo 2017 se tomó como referencia el índice de IFHB registrado por los testigos Seebe (1.2, R), Shenmai 1 (7.8, MR) y Kasota (18.6, S) (Cuadro 3) para la clasificación de las líneas. En este ciclo se registró incidencia de la enfermedad que varió de 0 a 100 % y de severidad con nivel máximo de 5 (0 a 43 %), dependiendo del sublote y de la línea.
Considerando lo anterior, las líneas se clasificaron como: 103 líneas (78.6 %) R; 13 líneas (9.9 %) MR; y 15 líneas (11.4 %) S por haber registrado un valor de 0 a 5, de 6 a 11, y ≥ de 12 de IFHB, respectivamente. Es posible que la mayor severidad (nivel 5) registrada en este ciclo de 2017 con relación a la registrada (nivel 3) en el 2015, en las espigas inoculadas por aspersión, se deba a varios factores: al inóculo empleado (5 cepas vs 1 cepa), nivel de inóculo (70 000 vs 100000 conidios mL-1), al origen de la cepa (cebada vs trigo), al número de inoculaciones (2 vs 1), a la etapa fenológica (emisión de espigas vs grano lechoso) y a las condiciones ambientales del invernadero. En el Cuadro 4 se ilustra la clasificación de 56 líneas por tipo de respuesta R, MR o S para el ciclo 2017.
En la Figura 1 se ilustran los síntomas de la fusariosis en las espiguillas y en el raquis de espigas inoculadas con F. graminearum.

Figura 1 Espigas de líneas de cebada maltera después de la inoculación por aspersión con mezcla de cepas CU1R30, SL3R23, SL3R24, SL10R10 y CU6R30 de F. graminearum en invernadero en 2017. Desarrollo de fusariosis: A) línea 29 sin síntomas; B) línea 123 con índice bajo; C) línea 62 con índice moderado; D) línea 33 con índice alto. Espigas en estado de grano masoso (A) y grano lechoso (B, C, y D). La flecha indica el oscurecimiento y chupado de espiguillas enfermas.
Los granos enfermos se caracterizaron por presentar aspecto arrugado, chupado y coloración café, síntomas característicos de la enfermedad. El análisis estadístico de los valores de incidencia y severidad de la enfermedad mostró diferencia (p≤ 0.01) entre las líneas y los sublotes (Cuadro 5).
Cuadro 5 Resultados del análisis con el Modelo Lineal Generalizado de las variables registradas en las líneas avanzadas de cebada maltera inoculadas con F. graminearum durante 3 años, en invernadero.
Factor |
GLN† |
GLD‡ |
% incidencia |
% severidad |
IFHB |
||||||
2015 |
2016 |
2017 |
2015 |
2016 |
2017 |
2015 |
2016 |
2017 |
|||
Sublote |
5 |
138 |
1 |
0.0001 |
0.0001 |
1 |
0.0001 |
0.0001 |
1 |
0.0001 |
0.0001 |
L (S) |
133 |
138 |
0.0001 |
0.0001 |
0.0001 |
0.0001 |
0.0001 |
0.0001 |
0.0001 |
0.0001 |
0.0001 |
Modelo Lineal Generalizado, distribución Poisson, link (log); †GLN= grados de libertad del numerador; ‡GLD= grados de libertad del denominador; L(S)= Líneas (Sublotes)
Esta diferencia se puede atribuir a los aspectos mencionados anteriormente o a una combinación de ambos además de la variabilidad genética entre líneas por el número de ciclos de selección que existe en cada sublote y al tipo de inoculación.
Detección y cuantificación de DON en semilla inoculada
La inoculación de espigas de 56 líneas de los sublotes Elite (10 líneas), 01 (ocho líneas), 02 (11 líneas), 02A (14 líneas) y 03 (13 líneas), por aspersión con la mezcla de cepas CU1R30, SL3R23, SL3R24, SL10R10 y CU6R30 de F. graminearum, en invernadero en 2017, resultó en la acumulación de toxina DON en semilla de 49 líneas de cebada maltera analizadas (Cuadro 4). La semilla de las líneas 28, 65, 73, 90, 91, 96 y 112 registró valores por debajo del límite de detección (0.2 ppm kg-1) y del límite de cuantificación (0.36 ppm kg-1) para la avena, de acuerdo al protocolo de RIDASCREEN®FAST DON (r-Biopharm).
Tomando como referencia el intervalo de valores de acumulación de DON en los testigos Seebe (R, 3.4 - 4 ppm kg-1), Shenmai 1 (MR, 5.8-10.1 ppm kg-1) y Kasota (S, 22.0-22.8 ppm kg-1), las líneas se clasificaron como: R, 26 líneas, MR, 14 líneas y S, 16 líneas (Cuadro 4). Se encontró bajo coeficiente de correlación (r = 0.5682) al relacionar los valores promedio de contenido de DON y de severidad de la enfermedad de las 56 líneas y de las tres variedades testigo (Figura 2).
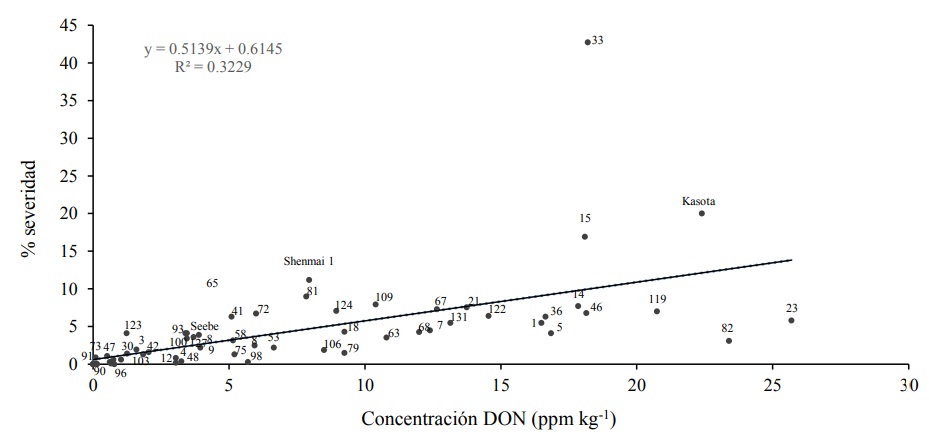
Figura 2 Correlación de severidad de la enfermedad y acumulación de DON en semilla de espigas inoculadas por aspersión con mezcla de cepas de F. graminearum, en invernadero en 2017. Los números dentro de la figura indican el número de la línea.
Esta baja correlación se ilustra con los valores obtenidos por las líneas 23, 82 y 119 (23.2 ppm kg-1 en promedio de DON) clasificadas como R en ese mismo año (Cuadro 4). En contraste las líneas 15, 33 y 131, clasificadas como S a la fusariosis en 2017, acumularon alto contenido de DON (16.4 ppm kg-1 en promedio). Por lo anterior las líneas 15 y 33 de los sublotes Elite y 01, respectivamente, podrían considerarse como testigos S en estudios futuros de cruzamientos para búsqueda de resistencia en cebada.
En la literatura no se encontró consenso en la relación entre el contenido de DON y las variables relacionadas con la producción. Mientras que Geddes et al. (2008) y Khatibi et al. (2012) reportaron baja correlación (r= 0.36 y r= 0.44, respectivamente) entre el contenido de DON y la severidad de la fusariosis en cebada, Chrpova et al. (2011) reportaron alta correlación positiva entre los valores de síntomas visuales de la enfermedad y los de contenido de DON (r= 0.76; p< 0.01) así como con los del contenido de DON y el peso de grano por espiga (r= 0.62; p< 0.01) y entre los del contenido de DON y los de peso de mil granos (r= 0.71; p< 0.01) en ese mismo cultivo.
Li et al. (2015) reportaron que plantas transgénicas de trigo que sintetizan la enzima UDP-glucosiltransferasa (HvUGT13248) de cebada exhibieron alta resistencia a la dispersión de la fusariosis en el raquis; es decir, mostraron resistencia tipo II. La enzima HvUGT13248 metaboliza a la toxina DON a un compuesto menos tóxico (DON3-O-glucósido). En el presente estudio se desconoce el o los mecanismos involucrados en aquellas líneas clasificadas en 2017 como R (26 líneas) o MR (14 líneas) a la acumulación de DON y a la infección por F. graminearum (resistencia tipo I). Al analizar las respuestas para índice IFHB y acumulación de DON del Cuadro 4, se deduce que, en general, conforme se fue avanzando en el ciclo de selección, es decir del sublote 03 (menor ciclo de selección) al sublote Elite (mayor ciclo de selección) se fueron presentando menos líneas resistentes a la fusariosis y más líneas susceptibles a la acumulación de DON.
Porque al transformar el número total de líneas de cada sublote en porciento, los sublotes de menor ciclo de selección presentaron un porcentaje más alto de líneas clasificadas como R. En contraste, los sublotes de mayor ciclo de selección registraron más porcentaje de líneas clasificadas como S.
Conclusiones
Durante tres años las líneas avanzadas 30, 47, 65 y 123, y las líneas 73, 91 y 96, de seis y dos hileras de grano, respectivamente, registraron síntomas muy bajos de fusariosis de la espiga en respuesta a la inoculación con F. graminearum. Aunque la severidad de la enfermedad no se relacionó (r= 0.5682) con la acumulación de DON, estas líneas registraron muy baja o nula producción de toxina. La información generada en este estudio es relevante ya que indica que estas siete líneas son resistentes a la enfermedad y tienen potencial para utilizarse como fuentes de resistencia en un programa de cruzamientos en cebada maltera que incluya en el conjunto de enfermedades a la fusariosis de la espiga.

