










Services on Demand
Journal
Article
 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista mexicana de ciencias agrícolas
Print version ISSN 2007-0934
Rev. Mex. Cienc. Agríc vol.10 n.2 Texcoco Feb./Mar. 2019
https://doi.org/10.29312/remexca.v10i2.732
Articles
Genetic diversity in accessions of 10 Mexican races of intermediate altitude maize
1Cátedras CONACYT-Universidad Autónoma Agraria Antonio Narro. Calzada Antonio Narro núm. 1923, Buenavista, Saltillo, Coahuila. CP. 25315. (msanchezv@conacyt.mx).
2Campus Montecillo-Colegio de Postgraduados. Carretera Mex-Tex km 36.5, Montecillo, Texcoco, México. CP. 56230. (sanchez.miriam@colpos.mx; lcordova@colpos.mx; asvarela@colpos.mx; fcastill@colpos.mx; arobledo@colpos.mx).
3Centro Universitario del Sur-Universidad de Guadalajara. Av. Enrique Arreola Silva 883, Ciudad Guzmán, Jalisco, México. CP. 49000. (claudia.saucedo@cusur.udg.mx).
The diversity of maize (Zea mays L.) in Mexico is wide and there is still a need to carry out works that allow to discern the variation within and between racial groups and establish collections of racial reference for their understanding. The morphological diversity of representative populations of 10 mexican maize races was assessed. In the spring-summer cycle of 2010, experiments were established with 88 representative accessions of Bofo, Celaya, Coscomatepec, Dulce de Jalisco, Elotes Occidentales, Mushito, Palomero de Jalisco, Serrano de Jalisco, Tablilla de Ocho, and Zamorano Amarillo races, in the localities of Roque, Guanajuato and Ciudad Guzmán, Jalisco, under an experimental design of incomplete blocks, 30 morphological characters were measured. The analysis of variance indicated significant differences among the accessions for all the characters (variables). The repeatability analysis showed that 16 characters had a coefficient ≥ 3.0, these were the least affected by the environment. In the analysis of main components and conglomerates, variation within and between races was observed, with a continuous dispersion, but which allowed the identification of four racial complexes. Group I was made up of accessions mainly from Bofo, Celaya II, Dulce de Jalisco III and Elotes Occidentales IV, the remaining races are mostly added in group II as subgroups. This study confirmed clustering of accessions in well-defined races and represented in germplasm banks. It is necessary to conduct deeper studies in races with few accessions to achieve a better racial definition.
Keywords: morphological characters; multivariate analysis; racial groups
La diversidad del maíz (Zea mays L.) en México es amplia y aún existe la necesidad de llevar a cabo trabajos que permitan discernir la variación dentro y entre grupos raciales y establecer colecciones de referencia racial para su entendimiento. Se valoró la diversidad morfológica de poblaciones representativas de 10 razas de maíz de México. En el ciclo primavera-verano de 2010 se establecieron experimentos con 88 accesiones representativas de las razas Bofo, Celaya, Coscomatepec, Dulce de Jalisco, Elotes Occidentales, Mushito, Palomero de Jalisco, Serrano de Jalisco, Tablilla de Ocho, y Zamorano Amarillo, en las localidades de Roque, Guanajuato y Ciudad Guzmán, Jalisco, bajo un diseño experimental de bloques incompletos, se midieron 30 caracteres morfológicos. El análisis de varianza indicó diferencias significativas entre las accesiones para todos los caracteres (variables). El análisis de repetibilidad reflejó que 16 caracteres presentaron un coeficiente ≥3, éstos fueron los menos afectados por el ambiente. En el análisis de componentes principales y conglomerados hubo variación dentro y entre razas, con una dispersión continua, que permitió identificar cuatro complejos raciales. El grupo I lo integraron accesiones de Bofo, el II de Celaya, el III de Dulce de Jalisco y el IV de Elotes Occidentales, las razas restantes se agregan en su mayoría en el grupo II como subgrupos. Este estudio confirmó agrupamientos de accesiones en razas bien definidas y representadas en los bancos de germoplasma. Es necesario realizar estudios más profundos en razas con pocas accesiones para lograr una mejor definición racial.
Palabras claves: análisis multivariado; caracteres morfológicos; grupos raciales
Introduction
Mexico is recognized as the center of origin and domestication of maize, so there is a great diversity of types that have been adapted to specific environmental and sociocultural conditions (Matsuoka et al., 2002). In Mexico, the genetic variability of maize is the result of human interaction for thousands of years with wild relatives and the environment (CONABIO, 2013). In this process, the mechanisms of dispersion are given, the genetic recombination by natural crosses between maize and teosinte (Z. mays: ssp. huehuetenanguensis, ssp. mexicana and ssp. parviglumis, among others); through, the geography of the country and the wide diversity of human groups, who make selection according to their customs in specific ecological niches (Orozco-Ramírez et al., 2016).
To understand genetic diversification, morphological differentiation among native maize populations has been studied by applying the concept of race (Anderson and Cutler, 1942). The physiological and morphological reproductive traits have been used, mainly as a tool to assess diversity (Wellhausen et al., 1952; Hortelano et al., 2008; Ángeles et al., 2010; Rocandio-Rodríguez et al., 2014). To achieve an accurate racial assessment, it is necessary to know the variation between variants within the races (Castillo, 1993), with the purpose of designing the use of some forms within the regional genetic diversity of the species.
To date, approximately 62 maize races have been described, a number that increases or decreases slightly according to the considerations of each study (Sánchez et al., 2000; Vielle-Calzada and Padilla, 2009; Hortelano et al., 2012). Many of the native maize populations collected in Mexico are intermediate variants between races; however, once the characteristics of the variants described as representative of races are known, possible routes of racial diversification can be proposed (Wellhausen et al., 1952).
Studies of maize diversity in Mexico have included few representative samples of the reported breeds (Wellhausen et al., 1952), or have focused on diversity in specific geographic areas (Herrera-Cabrera et al., 2004; Mijangos-Cortes et al., 2007; Ángeles et al., 2010). Knowledge of genetic diversity is essential to: expand the sources of germplasm, try to minimize the risks of genetic vulnerability, increase the probability of detecting favorable alleles and conserve plant genetic resources (Bellon, et al., 2009).
The contributions of the Wellhausen et al. (1952); Hernández and Alanis (1970); Ortega (1979); Sanchez and Goodman (1992); Sánchez et al. (2000), among others, in breeds such as Bofo, Celaya, Coscomatepec, Dulce de Jalisco, Elotes Occidentales, Mushito, Palomero de Jalisco, Serrano de Jalisco, Tablilla de Ocho and Zamorano Amarillo indicate that the diversity of maize has been studied step to bring together existing racial genetic variability, intensively in different ecological and ethnic regions of the country, as well as their specialized forms of consumption.
This has laid the basis for current racial classifications by means of morphological, physiological, genetic, biochemical and molecular characters; however, the racial characterizations of some accessions in germplasm banks have been identified and classified in collections that are typical and do not correspond to the racial descriptions proposed by these authors.
In this context, the present work aimed to study the genetic diversity of representative accessions of ten maize races with adaptation to low to intermediate altitudes (100-2 200 m) through morphological characters to estimate genetic diversity between and within breeds, that allows to classify them by their intrinsic characteristics and to determine the condition of the type accessions in the germplasm banks.
Materials and methods
Genetic material
88 maize accessions were characterized, cataloged according to their passport data as belonging to 10 breeds in Mexico: Bofo (8 accessions), Celaya (15), Coscomatepec (7), Dulce de Jalisco (12), Elotes Occidentales (23), Mushito (5), Palomero de Jalisco (1), Serrano de Jalisco (3), Tablilla de Ocho (7) and Zamorano Amarillo (7). The seed was provided by the germplasm banks of the International Maize and Wheat Improvement Center (CIMMYT), the Autonomous University of Chapingo (UACH), the National Institute of Agricultural and Livestock Forestry Research (INIFAP) and the Postgraduate College (CP).
Experimental sites and cultivation cycles
Two experiments were established in the spring-summer 2010 cycle, the first one in the Experimental Field of the Technological Institute of Roque, Celaya, Guanajuato (20º 34’ 55’’ North latitude, 100º 49’ 33’’ West longitude, altitude of 1 766 meters above sea level) and the second at Rancho Los Paredones, Cd. Guzmán, Jalisco (19º 43’ 09’’ North latitude, 103º 29’ 57’’ West longitude, altitude of 1 515 m), both with irrigation.
Design and experimental unit
The experiments were established under an experimental design in incomplete blocks 9 x 10, with two repetitions, in which the 88 accessions were included. The experimental unit consisted of two furrows 5 m long and 0.85 m wide. Three seeds per plant were planted 0.50 m apart, six weeks after sowing, thinning to leave two plants per plant for a population density of 47 000 plants ha-1 (Rocandio-Rodríguez et al., 2014).
Measured characters
From each plot, five plants with complete competence were randomly labeled, in which 30 variables were measured following the procedures indicated by Sánchez et al. (1993).
Vegetative and phenological characters: in the final stage of the vegetative development (when the last leaf appears at the apex of the plant), the plant height (AP), height at the ear (AMz), length of the cob leaf (LHMz) and width of the maize husk (AHMz), these expressed in cm, number of total leaves (HojP), leaves above the ear (HAMz) and of buds per plant (HijP). In the flowering stage, the days of female flowering (FF) and male flowering (FM) were recorded, days after sowing, the first when 50% of the plants of the experimental unit presented exposed stigmas and the second when 50% of the plants presented pollen emission, floral synchrony (SF, FF-FM) and the AMz/AP ratio.
Spike characters: one week after anthesis the total length of the spike (LE), the peduncle (LPE), of the central branch (LRCE, central axis from the insertion point of the highest lateral branch) was measured), and of the branched section (LTRE), these variables were measured in cm, the number of primary branches (RPE), and the LTRE/LE ratio were also evaluated.
Characteristics of the ear: length (LMz), diameter (DMz, in the center of the ear), peduncle length (LPMz), ear diameter (OD) and number of rows (HMz), were measured The LMz/DMz ratio was estimated.
Grain characters: the ears were individually shelled and from the central part a sample of 10 grains per ear was taken to measure the width (AGr), length (LGr), thickness (EGr), the three variables in mm and AGR/LGr ratio, another sample of 100 grains was taken, to measure the weight (P100Gr, g), volume (V100Gr, cm3) and the coefficient of shelling in percentage (CD=[(weight of grain) x (100)/total weight of the cob]).
Statistical analysis
An analysis of variance was applied through localities. The variance components for genotypes (σ2gen), localities (σ2loc) and their interaction (σ2gen × loc) were estimated and the repeatability coefficient r = [σ2gen / (σ2loc + σ2gen × loc)], was estimated. used to perform a selection of variables, it was taken as a criterion to accept the characters with r≥ 3 (Goodman and Paterniani, 1969). The selected variables were used to perform the analyzes described below.
A principal components analysis was applied based on the correlation matrix, the dispersion of the accessions was plotted in the plane determined by the first two components (biplot, described by Arellano et al., 2014) and a cluster analysis in the that the Euclidean distance coefficient and the grouping method of the unweighted arithmetic mean (UPGMA) were used.
The choice of the cut of the dendrogram was based on the visual comparison of the groupings obtained in the analysis of main components, which was based on significant correlations (α≤ 0.05) in addition, that the repeatability analysis ensures the use of uniform variables with greater variability for the formation of groups; with this information, the decision of the cut was established by the researcher which required knowledge of the plant material studied and an adequate base of assumptions that help in the definition of groups and distances of similarity (Núñez-Colin et al., 2004). Statistical statistical analysis system (SAS) version 9.3 packages for Windows (SAS Institute, 2002) and NTSYSpc version 2.21 h (Rohlf, 2009) were used.
Results and discussion
Variance analysis
The analysis of combined variance of the 88 genotypes across localities identified significant statistical differences in the totality of the evaluated variables, which indicates the presence of ample genetic diversity among the accessions of the 10 maize races studied (Table 1). These results agree with those reported by Herrera et al. (2000); Chávez-Servia et al. (2011); Hortelano et al. (2012) and those who point out that the statistical differences between native maize is an indicator of genetic diversity in them for the character under analysis.
Table 1 Mean squares of the analysis of variance combined through localities, for 30 morphological characters.
| Variable | Source of variation | Media | CV (%) | ||||
| Loc | Rep/Loc | Gen | Gen x Loc | Error | |||
| gl | 1 | 2 | 89 | 89 | 178 | ||
| FF | 2992.9** | 253.25** | 642.86** | 67.26* | 49.26 | 83.61 | 8 |
| FM | 1886.04** | 396.01** | 665.69** | 71.27* | 52.07 | 82.33 | 9 |
| SF | 513.61** | 8.62 NS | 8.03** | 6.48** | 3.2 | 5.08 | 35 |
| HijP | 0.53* | 0.05 NS | 0.3** | 0.17* | 0.12 | 0.44 | 80 |
| AP | 25959.3** | 579 NS | 10527.57** | 1041.71** | 389.92 | 245.95 | 8 |
| AMz | 88304.13** | 204.75 NS | 8360.16** | 922.63** | 280.96 | 146.38 | 11 |
| AMz/AP | 0.59** | 0 NS | 0.01** | 0NS | 0 | 0.58 | 7 |
| HojP | 235.38** | 10.13** | 15.62** | 1.7** | 0.96 | 13.87 | 7 |
| HojAMz | 72.63** | 0.36 NS | 1.27** | 0.26** | 0.16 | 5.34 | 8 |
| LHMz | 7758.08** | 114.25 NS | 484.15** | 57.41NS | 51.66 | 95.57 | 8 |
| AHMz | 0.09 NS | 1.74* | 4.11** | 0.86** | 0.54 | 8.5 | 9 |
| LE | 246.18** | 14.41 NS | 70.7** | 22.36NS | 17.35 | 43.6 | 10 |
| LTRE | 711.07** | 1.19 NS | 29.72** | 7.92** | 5.12 | 14.48 | 16 |
| LTRE/LE | 0.23** | 0 NS | 0.01** | 0** | 0 | 0.33 | 18 |
| LPE | 24.08 NS | 11.56 NS | 30.66** | 11.87 NS | 10.89 | 25.71 | 13 |
| LRCE | 130.3** | 8.99 NS | 40.98** | 21.47 NS | 16.62 | 29.37 | 14 |
| RPE | 409.17** | 1.24 NS | 40.49** | 12.01 NS | 9.07 | 15.46 | 19 |
| LMz | 38.12** | 1.23 NS | 24.49** | 3.54** | 1.84 | 18.32 | 7 |
| DMz | 0.53* | 0.01 NS | 0.61** | 0.11* | 0.08 | 4.56 | 6 |
| LMZ/DMz | 0.15 NS | 0.01 NS | 1.35** | 0.34* | 0.25 | 4.07 | 12 |
| HMz | 53.38** | 0.31 NS | 17.04** | 2.02** | 0.99 | 12.36 | 8 |
| LPMz | 2220.95** | 19.93 NS | 37.72** | 16.57** | 9.77 | 14.42 | 22 |
| DO | 0 NS | 0 NS | 0.3** | 0.06* | 0.04 | 2.42 | 9 |
| LGr | 15.57** | 0.25 NS | 4.92** | 0.73 NS | 0.62 | 12.55 | 6 |
| AGr | 11.26** | 0.25 NS | 7.81** | 0.55* | 0.36 | 9.74 | 6 |
| AGr/LGr | 0.21** | 0 NS | 0.05** | 0 NS | 0 | 0.78 | 11 |
| EGr | 4.01** | 0.18 NS | 1.02** | 0.22* | 0.15 | 4.72 | 8 |
| P100Gr | 2026.79** | 0.26 NS | 375.02** | 55.35** | 31.37 | 42.04 | 13 |
| V100Gr | 8120.2** | 35.73 NS | 979.02** | 136.31** | 72.99 | 65.03 | 13 |
| CD | 323.88** | 43.13 NS | 49.75** | 24.82 NS | 22.13 | 83.77 | 6 |
*, **, NS = significant difference α= 0.05, α= 0.01, not significant, respectively; gl= degrees of freedom. Loc= localities; Rep/Loc= repetitions nested in localities; Gen= genotypes; Gen × Loc= genotype interaction by location.
In the interaction genotypes by locality, 70% of the total of the variables were statistically significant, indicating that at least one of the accessions presented a differential response in the characters when changing locality, these results agree with those reported by Rocandio-Rodríguez et al. (2014).
Selection of variables
The repeatability analysis (r) showed very marked differences, with values between 0.09 and 11.78 for the characters studied and of the total of them, 16 had a value of r≥ 3, 63% of these correspond to cob characters (Table 2), which is in agreement with that reported by Hortelano et al. (2012).
Table 2 Estimators of the components of variance and repeatability value (r).
| Variable | σ2Gen x Loc | σ2Gen | σ2Loc | r |
| Flowering female (FF) | 9 | 143.9 | 15.12 | 5.97 |
| Male flowering (FM) | 9.6 | 148.6 | 8.17 | 8.36 |
| Floral synchrony (SF) | 1.64 | 0.39 | 2.79 | 0.09 |
| Buds by plant (HijP) | 0.02 | 0.03 | 0 | 1.24 |
| Plant height (AP) | 325.9 | 2371.46 | 137.38 | 5.12 |
| Cob height (AMz) | 320.83 | 1859.38 | 485.88 | 2.3 |
| AMz/AP ratio | 0 | 0 | 0 | 1.17 |
| Total leaves per plant (HojP) | 0.37 | 3.48 | 1.25 | 2.15 |
| Leaves above the ear (HAMz) | 0.05 | 0.25 | 0.4 | 0.57 |
| Length of the cob leaf (LHMz) | 482.67 | 2541.86 | 4009.55 | 0.57 |
| Width of the cob leaf (AHMz) | 0.16 | 0.81 | -0.01 | 5.32 |
| Total length of the spike (le) | 2.51 | 12.08 | 1.26 | 3.21 |
| Length of the branched section of the spike (ltre) | 1.4 | 5.45 | 3.93 | 1.02 |
| LTRE/LE ratio | 0 | 0 | 0 | 0.65 |
| Length of the spindle peduncle (LPE) | 0.49 | 4.7 | 0.06 | 8.52 |
| Length of the central branch of the spike (LRCE) | 2.42 | 4.88 | 0.65 | 1.59 |
| Primary branches of the spike (RPE) | 1.47 | 7.12 | 2.25 | 1.91 |
| Length of ear (LMz) | 0.85 | 5.24 | 0.2 | 5.02 |
| Diameter of ear (DMz) | 0.02 | 0.12 | 0 | 6.08 |
| LMz/DMz ratio | 0.04 | 0.25 | 0 | 5.75 |
| Rows per maize (HMz) | 0.52 | 3.75 | 0.29 | 4.65 |
| Length of the cob peduncle (LPMz) | 3.4 | 5.29 | 12.19 | 0.34 |
| Diameter of the olote (DO) | 0.01 | 0.06 | 0 | 7.53 |
| Grain length (LGr) | 0.05 | 1.05 | 0.08 | 7.63 |
| Grain width (AGr) | 0.09 | 1.82 | 0.06 | 11.78 |
| Agr/LGr ratio | 0 | 0.01 | 0 | 5.22 |
| Thickness of grain (EGr) | 0.04 | 0.2 | 0.02 | 3.47 |
| Weight of 100 grains (P100Gr) | 11.99 | 79.92 | 11.13 | 3.46 |
| Volume of 100 grains (V100Gr) | 31.66 | 210.9 | 44.56 | 2.77 |
| Grain with respect to the ear (% GrMz) | 1.34 | 6.23 | 1.54 | 2.16 |
σ2Gen= component of variance for genotypes; σ2Loc= for localities; σ2Gen x Loc= for interaction between genotypes and localities and r= repeatability coefficient.
The characters with r≥ 3 correspond mostly to reproductive descriptors, which are less affected by environmental factors, in this sense, these characters reflect high genetic variability and contribute in a better way to their explanation (González et al., 2013; Rocandio-Rodríguez et al., 2014).
Principal component analysis
The principal components analysis indicated that the first five components explain 88% of the total observed variation and the first two components close to 60% (data not shown). Ferraz et al. (2013) mention that a high proportion explained with a low number of components, allows better interpretation of the variation and studies carried out with characters related to yield in maize, show that the models that involve the biplot graphs are useful to determine the intensity and nature of native maize populations under certain environmental conditions (Martínez-Sánchez et al., 2016).
The variables that showed the highest absolute values in the coefficients of the eigenvectors of the first two components, and that determine in a relevant way the racial dispersion, as well as the formation of groups were: for the component one FM, LMz, AGr and P100Gr and for component two AHMz, HMz, AGr/LGr and EGr (Table 3).
Table 3. Own vectors associated with the first two main components of the analysis of 16 variables, in 88 type accessions of 10 maize races
| Variable | Vector own | |
| CP-2 | CP-2 | |
| Days to feminine flowering (FF) | 0.298 | 0.242 |
| Days to male flowering (FM) | 0.301 | 0.223 |
| Plant height (AP) | 0.27 | 0.268 |
| Width of the cob leaf (AHMz) | 0.23 | 0.343 |
| Length of the spike (LE) | 0.296 | 0.069 |
| Length of the spindle peduncle (LPE) | -0.021 | -0.105 |
| Length of the ear (LMz) | 0.346 | 0.033 |
| Diameter of the ear (DMz) | 0.086 | 0.321 |
| LMz/DMz ratio | 0.277 | -0.157 |
| Number of rows of cob (HMz) | -0.262 | 0.333 |
| Diameter of the olote (DO) | 0.136 | 0.189 |
| Grain length (LGr) | 0.103 | 0.295 |
| Grain width (AGr) | 0.331 | -0.226 |
| AGr/LGr ratio | 0.246 | -0.387 |
| Thickness of the grain (EGr) | 0.187 | -0.316 |
| Weight of 100 grains (P100Gr) | 0.316 | -0.155 |
The dispersion of the accessions according to the first two main components makes it possible to differentiate the following four groups (Figure 1): group I: composed of 100% of the accessions of the Bofo breed, 57% Zamorano Amarillo, 43% Tablilla de Echo and 17% of Elotes Occidentales, which share morphological characteristics related to LPE (26.97 cm), EGr (5.01 mm), AGr (10.9 mm), P100Gr (49.46 g) and the LMz/DMz (4.62) and AGr/LGr (0.9) ratios.
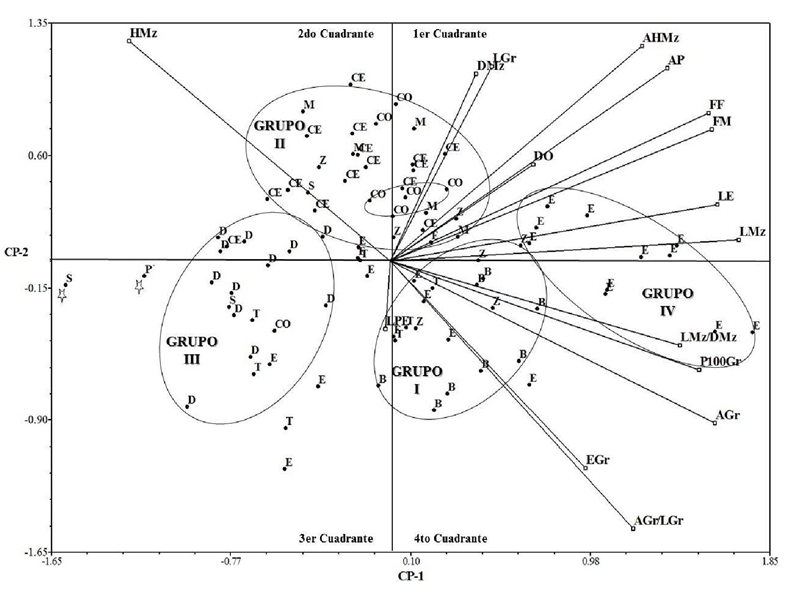
Figure 1 Dispersion of 88 representative accessions of 10 maize races based on the first two main components. B= Bofo; CE= Celaya; CO= Coscomatepec; D= Dulce de Jalisco; E= Elotes Occidentales; M= Mushito; P= Palomero de Jalisco; S= Serrano de Jalisco; T= Tablilla de Ocho; Z= Zamorano Amarillo.
The total grouping of the accessions of the Bofo breed is indicative of a marked genetic differentiation of these. The low number of accessions of the other breeds in this group is possibly due to the fact that these accessions share a certain geographical area in their distribution (100 to 1400 meters above sea level) and greater variability, although it is also possible that a priori racial identification does not It has been successful. These races were classified by Sánchez and Goodman (1992); Sánchez et al. (2000) within a group of maize called ‘Ocho Hileras’ and in particular Zamorano Amarillo as a subgroup within it.
According to Wellhausen et al. (1952) and Hernández and Alanis (1970), the Bofo and Tablilla de Ocho breeds share the Tabloncillo breed as a common ancestor, which also partly explains the location of both breeds in this group.
Group II: integrates 100% of the accessions of Mushito, 86% of Celaya, 86% of Coscomatepec, 43% of Zamorano Amarillo, 33% of Serrano de Jalisco and 8% of Elotes Occidentales. The morphological characteristics that distinguish this group are: HMz (13.13 rows), DMz (4.8 cm), LGr (13.11 mm), AHMz (9.20 cm), AP (261.04 cm), FF (86.81 days) and FM (84.79 cm).
Considering the number of accessions, in this group the Celaya race predominates, which is a well-defined race (Wellhausen et al., 1952; Sánchez and Goodman, 1992), while Mushito is mentioned as an ill-defined race (Wellhausen et al., 1952; Bretting and Goodman, 1989), which also does not have a number of considerable and representative accessions in germplasm banks, like the rest of the breeds in this group and even when they have some resemblance to the Celaya breed, its characteristics suggest that they constitute different genetic groups, which need to be studied in greater depth, this agrees with Hernández and Alanis (1970); Sánchez and Goodman (1992).
Group III: 92% of the accessions of Dulce de Jalisco, 33% of Serrano de Jalisco, 28% of Tablilla de Ocho, 14% of Coscomatepec, 6% of Celaya and 4% of Elotes Occidentales. In general they are the earliest populations, low in height of plant and ear, reduced size of spike, ear and grain (72.57 days at FF, 71.51 days at FM, 194.95 cm from AP, 13.45 HMz, 7.38 cm from AHMz , 39.03 cm of LE, 25.52 cm of LPE, 15.68 cm of LMz, 36.19 g in P100Gr, 4.45 cm of DMz, 11.55 mm of LGr, 8.95 mm of AGr and 4.88 mm of EGr), the group is located predominantly in the third quadrant of Figure 1; that is, the accessions generally have negative values of the first two main components.
Group IV: composed of 48% of Elotes Occidentales accessions which are characterized by being late FF and FM (106.45, 106.68 days), with large spike and ears (LMz, 21.10 cm, LE, 49.07 cm), heavy grain (P100Gr, 57.96 g) and LMz/DMz of 4.60, which indicates own slender ears for fresh consumption.
The Palomero de Jalisco breed was not integrated into any group, due to the low representativeness of accessions that occurred in the study. These results agree with those reported by Vega-Alvárez et al. (2017), who mention that a small number of accessions in studies of group differentiation in maize, can affect the formation of these.
Cluster analysis. In the dendrogram (Figure 2) there is a continuous differentiation from lower to greater Euclidean distance between accessions and groups. In this regard, Orozco-Ramírez et al. (2016) report that the maize races are grouped in a continuum with respect to a geographical area, environmental variation, morphological characters and the relationship with the social origin of the maize races in southern Mexico.

Figure 2 Dendrogram of 88 accessions of 10 maize races, constructed with the UPGMA method using Euclidean distances considering 16 morphological variables. B= Bofo; CE= Celaya; CO= Coscomatepec; D= Dulce de Jalisco; E= Elotes Occidentales; M= Mushito; P= Palomero de Jalisco; S= Serrano de Jalisco; T= Tablilla de Ocho; Z= Zamorano Amarillo.
The diversity found in this analysis is manifested in four groups from a Euclidean distance of 5.16, which coincide between 68 and 100% with the trends of groupings in the analysis of major components (Figure 1), confirming that the best-defined breeds are for group one Bofo, for two, Celaya with the rest of the races in small groups or under a continuum; for the three Dulce de Jalisco and for the four Elotes Occidentales. Perales et al., (2005) and mention that using morphological characteristics that are under the selection of farmers, it has been demonstrated that maize populations are maintained at different regional scales much smaller; likewise, Wellhausen et al. (1952) and later Vega-Alvárez et al. (2017) mention that studies based on morphological characteristics indicate that some of the local varieties arise from the hybridization of other previously existing breeds.
Results similar to those of this research are reported by Rocandio-Rodríguez et al. (2014), in a study of morphological and agronomic characterization of seven maize races of the High Valleys of Mexico, where they confirmed that the groupings and racial interrelations are more precise than other studies, due to the large sample sizes used.
Conclusions
There is a high phenotypic diversity for the characters evaluated in the accessions of the ten maize races studied. The representation of the type accessions was confirmed in the differentiation of the races Bofo, Celaya, Elotes Occidentales and Dulce de Jalisco, while those of Coscomatepec, Mushito and Zamorano Amarillo, tend to group in small conglomerates as a continuum. Serrano de Jalisco, Palomero de Jalisco, Tablilla de Ocho, are not defined in any grouping, so it is necessary to deepen this type of studies for these breeds with a greater number of accessions and with additional strategies such as molecular markers.
Literatura citada
Anderson, E. and Cutler, H. C. 1942. Races of Zea mays L. Their recognition and classification. Annals of the Missouri Botanical Garden. 29:69-88. doi:10.2307/2394331. [ Links ]
Ángeles, G. E.; Ortiz, T. E.; López, P. A. y López, R. G. 2010. Caracterización y rendimiento de poblaciones de maíz nativas de Molcaxac, Puebla. Rev. Fitotec. Mex. 33:287-296. [ Links ]
Arellano, V. J. L.; Rojas M., I.; Gutiérrez H., G. F. 2014. Variedades de maíz azul Chalqueño seleccionadas por múltiples caracteres y estabilidad del rendimiento. Rev. Mex. Cienc. Agríc. 5:1469-1480. [ Links ]
Bellon, M. R.; Barrientos, P. A. F.; Colunga, G. M. P.; Perales, H.; Reyes, A. J. A.; Rosales, S. R. y Zizumbo, V. D. 2009. Diversidad y conservación de recursos genéticos en plantas cultivadas. In: capital natural de México. Vol. II: estado de conservación y tendencias de cambio. Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (CONABIO). México, DF. 355-382 pp. [ Links ]
Bretting, P. K. and Goodman, M. M. 1989. Karyotypical variation in Mesoamerican races of maize and its systematic significance. Econ. Bot. 43:107-124. doi.org/10.1007/BF02859330. [ Links ]
Castillo, G. F. 1993. La variabilidad genética y el mejoramiento genético de los cultivos. Ciencia. 44:69-79. [ Links ]
Chávez, S. J. L.; Diego, F. P. y Carrillo, R. J. C. 2011. Complejos raciales de poblaciones de maíz evaluadas en San Martín Huemelulpan, Oaxaca. Rev. Soc. Cult. Des. Sust. Ra Ximhaí 7:107-115. [ Links ]
CONABIO. 2013. La diversidad biológica de México: maíces-razas de maíz. Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (CONABIO). México. http://www.biodiversidad.gob.mx/usos/maices/razas2012.html. [ Links ]
Ferraz, T. Y.; Permuy, A. N. y Acosta, R. R. 2013. Evaluación de accesiones de maíz (Zea mays L.) en condiciones de sequía en dos zonas edafoclimáticas del municipio Gibara, provincia Holguin. Evaluación morfoagronómica y estudios de la interacción genotipo x ambiente. Cultivos Tropicales. 34:24-30. [ Links ]
González, C. M. E.; Palacios, R. N.; Espinoza, B. A. y Bedoya, S. C. A. 2013. Diversidad genética en maíces nativos mexicanos tropicales. Rev. Fitotec. Mex. 36:239-338. [ Links ]
Goodman, M. M. and Paterniani, E. 1969. The races of maize: III. Choices of appropriate characters for racial classification. Econ. Bot. 23:265-273. [ Links ]
Hernández, X. E. y Alanís, F. G. 1970. Estudio morfológico de cinco nuevas razas de maíz de la Sierra Madre Occidental de México. Implicaciones filogenéticas y fitogeográficas. Agrociencia 5:3-30. [ Links ]
Herrera, C. B. E.; Castillo, G. F.; Sánchez, G. J. J.; Ortega, P. R. y Goodman, M. M. 2000. Caracteres morfológicos para valorar la diversidad entre poblaciones de maíz en una región: caso la raza Chalqueño. Rev. Fitotec. Mex. 23:355-354. [ Links ]
Herrera, C. B. E.; Castillo, G. F.; Sánchez, G. J. J.; Hernández, C. J. M.; Ortega, P. R. y Goodman, M. M. 2004. Diversidad de maíz Chalqueño. Agrociencia. 38:191-206. [ Links ]
Hortelano, S. R. R.; Gil, M. A.; Santacruz, V. A.; Miranda, C. S. y Córdova, T. L. 2008. Diversidad morfológica de maíces nativos del Valle de Puebla. Agric. Téc. Méx. 34:189-200. [ Links ]
Hortelano, S. R. R.; Gil, M. A.; Santacruz, V. A.; López, S. H.; López, P. A. y Miranda, C. S. 2012. Diversidad fenotípica de maíces nativos del altiplano centro-oriente del estado de Puebla, México. Rev. Fitotec. Mex. 35:97-109. [ Links ]
Martínez, S. J.; Espinosa, P. N. y Villegas, A. Y. 2016. Interacción genotipoambiente en poblaciones de maíz nativo de Chiapas. Rev. Mex. Agroecos. 3:38-48. [ Links ]
Matsuoka, Y.; Vigouroux, Y.; Goodman, M. M.; Sánchez, G. J.; Buckler, E. and Doebley, J. 2002. A single domestication for maize shown by multilocus microsatellite genotyping. Proceedings of the National Academy of Sciences of the United States of America. 99:6080-6084. https://doi.org/10.1073/pnas.052125199. [ Links ]
Mijangos, C. J. O.; Corona, T. T.; Espinosa, V. D.; Muñoz, O. A.; Romero, P. J. and Santacruz, V. A. 2007. Differentiation among maize (Zea mays L.) landraces from the Tarasca Mountain Chain, Michoacan, Mexico and the Chalqueño complex. Genetic Res. Crop Evol. 54:309-325. DOI 10.1007/s10722-005-4775-y. [ Links ]
Núñez, C. C. A.; Rodríguez, P. J. E.; Nieto, A. R. y Barrientos, P. A. F. 2004. Construcción de dendrogramas de taxonomía numérica mediante el coeficiente de distancia χ2: una revisión. Rev. Chapingo Ser. Hortic. 10:229-237. [ Links ]
Orozco, R. Q.; Ross, I. J.; Santacruz, V. A. and Brush, S. 2016. Maize diversity associated with social origin and environmental variation in Southern Mexico. Heredity. 116:477-484. DOI: 10.1038 / hdy.2016.10. [ Links ]
Ortega, P. A. 1979. Reestudio de las razas mexicanas de maíz. Informe Anual. Campo Agrícola Experimental de la Mesa Central. INIA, Chapingo, México. [ Links ]
Rocandio, R. M.; Santacruz, V. A.; Córdova, T. L.; López, S. H.; Castillo, G. F. y Lobato, O. R.; Ortega, P. R. 2014. Caracterización morfológica y agronómica de siete razas de maíz de los Valles Altos de México. Rev. Fitotec. Mex. 37:351-361. [ Links ]
Rohlf, F. J. 2009. NTSYSpc: numerical taxonomy system. Version 2.21h. Exeter Software: Setauket: New York. [ Links ]
Sánchez, G. J. J. and Goodman, M. M. 1992. Relationships among the Mexican races of maize. Econ. Bot. 46:72-85. DOI 10.1007/BF02985256. [ Links ]
Sánchez, G. J. J.; Goodman, M. M. and Rawlings, O. 1993. Appropriate characters for racial classification in maize. Econ. Bot. 47:44-59. https://doi.org/10.1007/BF02862205. [ Links ]
Sánchez, G. J. J.; Goodman, M. M. and Stuber, C. W. 2000. Isozymatic and morphological diversity in the races of maize of Mexico. Econ. Bot. 54:43-59. https://doi.org/10.1007/BF02866599. [ Links ]
SAS Institute. 2002. The SAS® System for Windows® (Version 9.3). Statistical Analysis System Institute Inc. Cary, NC. USA. 4424 p. [ Links ]
Vega, A. I.; Santacruz, V. A.; Rocandio, R. M.; Córdova, T. L.; López, S. H.; Muñoz, O. A. and Hernández, B. A. 2017. Genetic diversity and structure of native maize races from Northwestern Mexico. Pesquisa Agropecuária Brasileira. 52:1023-1032. DOI: 10.1590 / S0100-204X2017001100008. [ Links ]
Vielle, C. J. P. and Padilla, J. 2009. The Mexican landraces: description, classification and diversity. In: Bennetzen, J. L. and Hake, S. C. (Ed.). Handbook of maize: its biology. New York: Springer. 543-561 p. DOI: 10.1007/978-0-387-79418-1-27. [ Links ]
Wellhausen, E. J.; Roberts, L. M. y Hernández, X. E. 1952. Razas de maíz en México. Su origen, características y distribución. Oficina de Estudios Especiales. Secretaría de Agricultura y Ganadería. México, D F. Folleto técnico núm. 5. 237 p. [ Links ]
Received: January 2019; Accepted: February 2019
 Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
