











 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroducción
El arroz es el cereal más importante en el mundo, por la extensión de cultivo que ocupa y su consumo por casi 3 billones de personas (Zhao y Mc Grath, 2009); sin embargo, se informa como una planta hiperacumuladora de diversos metales pesados, siendo los más dañinos el cadmio, por su movilidad y toxicidad y el plomo, aunque no es un metal móvil, es muy tóxico (Reeves et al., 2000).
Los suelos agrícolas pueden contaminarse con metales pesados a través de desechos industriales, gasolina, pintura, desechos mineros, fertilizantes, pesticidas, abonos animales, irrigación de aguas residuales y otras fuentes (Khan et al., 2008). Los metales pesados más comúnmente encontrados en sitios contaminados son plomo (Pb), cadmio (Cd), mercurio (Hg), cromo (Cr), arsénico (As), zinc (Zn), cobre (Cu) y níquel (Ni) (Su et al., 2014).
Los metales pesados liberados al medio ambiente por actividades antropogénicas no pasan por un proceso de degradación química o biológica (microbiana) (Kirpichtchikova et al., 2006) y su concentración total en los suelos persiste durante mucho tiempo después de su introducción (Adriano, 2003). El nivel crítico de toxicidad en las especies cultivadas está en el rango de 10 μg g-1 de masa seca (MS) en plantas sensibles y 50 μg g-1 de MS en especies de plantas moderadamente sensibles y tolerantes (Marschner, 1995).
Varios autores han informado que la toxicidad del Pb y Ni podría producir daño celular oxidativo al generar especies reactivas de oxígeno (ROS) (Islam et al., 2011). Las ROS son el resultado del estrés oxidativo, que a su vez conduce a varios efectos perjudiciales sobre las células vegetales (Islam et al., 2008) por que las enzimas antioxidantes pueden disminuir o prevenir los efectos tóxicos de la ROS inducidas por el estrés metálico. Estas enzimas antioxidantes son superóxido dismutasa (SOD), ascorbato peroxidasa (APX), catalasa (CAT) y glutatión reductasa (GR). Estas enzimas pueden mantener el equilibrio entre la producción y la destrucción de ROS (Hassan y Mansoor, 2014).
Se ha estudiado la eficacia antioxidante bajo estrés de metales pesados en raíces y hojas, pero los informes sobre el sistema antioxidante en esos órganos debido a la presencia de Ni y Pb, que inducen estrés oxidativo en las mismas condiciones experimentales, son limitados. Por esta razón, el objetivo de esta investigación fue evaluar la influencia del Ni y el Pb en el desarrollo y el sistema antioxidante del arroz, mediante la determinación de las actividades de superóxido dismutasa (SOD) y glutatión reductasa (GR) en raíces y hojas en idénticas condiciones experimentales.
Materiales y métodos
Manejo del cultivo y diseño experimental
La investigación se realizó de septiembre de 2010 a marzo de 2011 en la Escuela Superior de Agricultura Luiz de Queiroz (ESALQ) en Brasil, en el laboratorio de Genética. La línea 8092 de arroz (O. sativa L.) fue proporcionada por el Instituto Nacional de Ciencias Agrícolas (INCA, Cuba). Las semillas se esterilizaron con hipoclorito de sodio al 5% durante 15 min, luego se enjuagaron con agua destilada tres veces, se sembraron en macetas llenas de agua destilada y se cubrieron con un paño fino para evitar la evaporación del agua. Las macetas se colocaron en un invernadero con una temperatura de 35 °C y una humedad relativa en un rango de 32-35%.
Después de una semana, las plántulas se trasplantaron a macetas de un litro con arena y solución nutritiva de Hoagland (600 ml), en cuatro tratamientos de cloruro de níquel (NiCl2) y acetato de plomo (PbAc2). El diseño experimental fue completamente aleatorizado, con cuatro niveles de NiCl2 y PbAc2 (0, 50, 100 y 300 mg kg-1). Todos los experimentos se realizaron por triplicado.
Los experimentos se llevaron a cabo en un invernadero bajo luz natural con temperaturas en el rango de 20-30 C. La solución de Hoagland con Ni y Pb se cambió semanalmente y el volumen total se completó con agua una vez por semana.
Muestreo
Las muestras vegetales se obtuvieron a los 30 días después de la siembra. Todas las muestras de raíces y hojas se tomaron en estado de madurez fisiológica. El material se desinfectó con detergente no iónico (1%), se enjuagó tres veces en agua destilada y luego se transfirió a papel de filtro. En cada muestreo, se empleó la materia fresca de las raíces y las hojas para el análisis de las enzimas antioxidantes y el perfil de isoenzimas.
Análisis de las muestras
Desarrollo de las plantas
Se evaluó el desarrollo de las plantas en presencia de los metales en estudio (NiCl2 y PbAc2) comparándolas con un control sin metales pesados.
Concentración de proteínas totales solubles
Se utilizó albúmina de suero bovino (BSA) como patrón para la determinación de la concentración de proteínas, por el método de Bradford (1976).
Extracción y análisis de enzimas antioxidantes
Las muestras de raíces u hojas se homogenizaron (volumen de tampón: masa fresca 2:1) en un mortero con tampón fosfato de potasio 100 mM pH 7,5 (ácido etilendiaminotetraacético (EDTA) 1 mM, DL-ditiotreitol 3 mM y PVP insoluble 5% p/v). El homogeneizado se centrifugó a 10 000 × g durante 30 min y el sobrenadante se mantuvo almacenado en alícuotas separadas a -80 °C, antes de los análisis GR y SOD. Todos los pasos de la extracción se llevaron a cabo a 4 °C.
La actividad de SOD se determinó a 560 nm (Beyer y Fridovich, 1987), basada en la inhibición de la reducción del nitroblue tetrazolium (NBT). Una unidad de actividad enzimática se definió como la cantidad de SOD requerida para una inhibición de 50% de la reducción de NBT. La actividad de SOD se expresó como μmol min-1 mg-1 de proteína.
La actividad GR total se determinó según lo descrito por Azevedo et al. (1998) a 412 nm durante 2 min, analizando la velocidad de reducción del glutatión oxidado (GSSG). La actividad de GR se expresó como μmol min-1 mg-1 de proteína.
Determinación de los perfiles isoenzimáticos de SOD y GR
Los ensayos de separación de isoenzimas SOD y GR y la clasificación de isoenzimas de SOD se ejecutaron según lo descrito por Gomes-Junior et al. (2007).
La electroforesis se llevó a cabo en geles no desnaturalizantes de poliacrilamida (PAGE) 8% durante 4 h a 4 °C, con una corriente constante de 20 mA/gel con 60 mg de proteína en cada carril.
Las bandas SOD se clasificaron usando peróxido de hidrógeno y KCN-teniendo en cuenta los patrones de inhibición con ambos productos químicos- como Mn-SOD (SOD I-resistente a ambos inhibidores) y Fe-SOD (SOD II-inactivada por uno de los inhibidores).
Análisis estadístico
Después de la comprobación de la normalidad y homocedasticidad de los datos (a través de las pruebas de Kolmogorov-Smirnov y de Bartlett, respectivamente), los datos de las actividades enzimáticas se sometieron a un análisis de varianza simple. Las diferencias entre las medias de los tratamientos se compararon mediante la prueba de Tukey (p< 0.05). Para los análisis se utilizó el paquete estadístico Statgraphics 5.1 (2001).
Resultados
Desarrollo de las plantas
El desarrollo de las plantas se afectó por ambos metales (Ni y Pb) (Figura 1). En comparación con el control, las plantas que crecieron en presencia de NiCl2 no desarrollaron completamente sus hojas y la altura de la planta se redujo en todas las concentraciones analizadas, especialmente las tratadas con 300 mg kg-1. Lo mismo ocurrió con las plantas a las que se aplicó PbAc2 en las concentraciones de 100 y 300 mg kg-1.

Análisis de enzimas antioxidantes
Superóxido dismutasa (SOD)
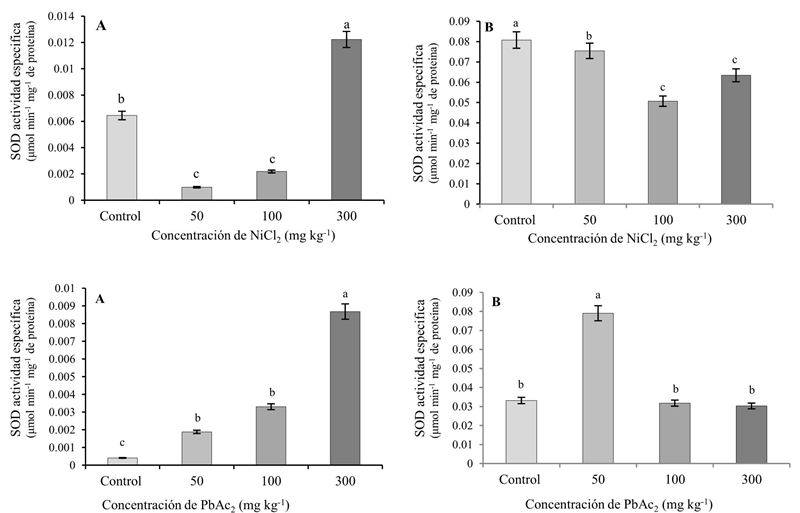
Cuando se utilizó NiCl2, la actividad específica de SOD en las raíces disminuyó significativamente con respecto al control en las concentraciones de 50 y 100 mg kg-1 y se incrementó significativamente en la mayor concentración (300 mg kg-1). Al aplicar PbAc2 la actividad de la enzima fue significativamente superior a la del control al utilizar 50 y 100 mg kg-1, alcanzándose el valor más alto con la concentración de 300 mg kg-1.
En las hojas de las plantas que crecieron en presencia de NiCl2, la actividad de la SOD disminuyó significativamente con respecto al control, alcanzándose los menores valores con 50 y 100 mg kg-1. El empleo de PbAc2 (50 mg kg-1) incrementó la actividad enzimática, obteniéndose valores estadísticamente similares al control en las concentraciones superiores (Figura 2).
Glutatión reductasa (GR)
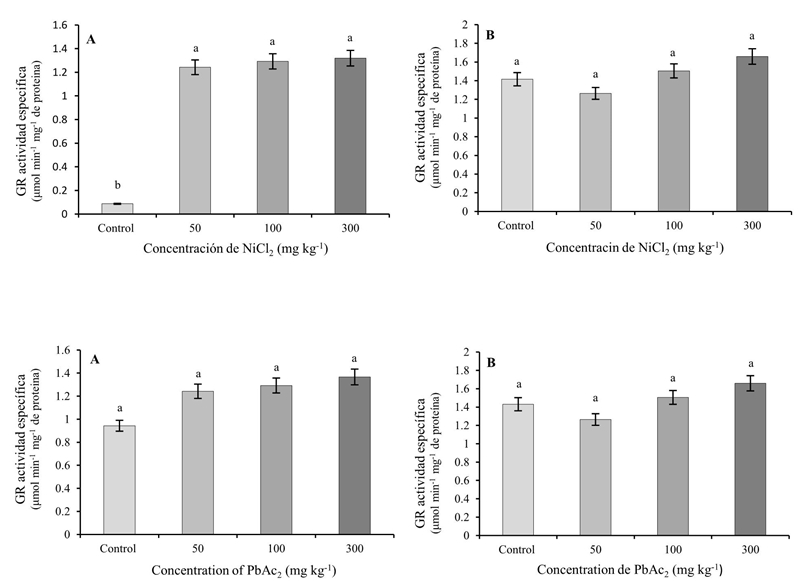
No se encontraron diferencias significativas en la actividad de esta enzima en las hojas y raíces debidas al empleo de PbAc2. En cambio, la aplicación de NiCl2 en todas las concentraciones usadas causó incrementos significativos en la actividad de GR en las raíces con respecto al control, sin provocar variaciones significativas en las hojas (Figura 3).
Perfiles isoenzimáticos
Superóxido dismutasa (SOD)
La presencia de bandas SOD fue similar al control en todas las concentraciones probadas (Figura 4). En cambio, se pudieron visualizar algunos cambios en el perfil, particularmente en las hojas, donde las bandas Fe-SOD a 50 y 100 mg kg-1 de NiCl2 exhibieron incrementos en su intensidad. El tratamiento con PbAc2 mostró en las raíces a 0 mg kg-1 que la Mn-SOD2 estaba ausente, mientras que cuatro bandas fueron muy intensas en las hojas a 100 mg kg-1 y ninguna banda se visualizó a 300 mg kg-1.
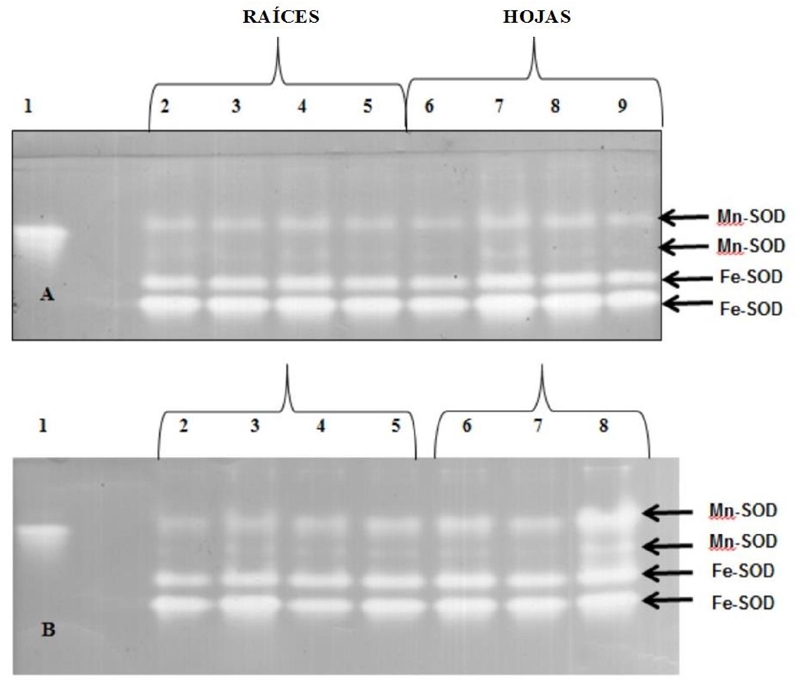
Figura 4 Perfiles isoenzimáticos (SOD) en PAGE de plantas de arroz (O. sativa L.) línea 8092. A- níquel (NiCl2), B- plomo (PbAc2). Carril 1= albúmina de suero bovino; Carril 2= control (0 mg kg-1); Carril 3= 50 mg kg-1; Carril 4= 100 mg kg-1; Carril 5= 300 mg kg-1; Carril 6=control (0 mg kg-1); Carril 7= 50 mg kg-1; Carril 8= 100 mg kg-1; Carril 9= 300 mg kg-1. Las flechas indican las isoenzimas SOD identificadas por tinción de los geles después de la incubación con KCN o H2O2 (5 mM en ambos casos).
Glutatión reductasa (GR)
Se identificaron cinco isoenzimas GR (Figura 5). GR V no apareció en las raíces de las plantas tratadas con NiCl2, pero sí estuvo presente en las hojas, con mayor intensidad en las bandas correspondientes a los tratamientos con 50 y 100 mg kg-1, en estos dos, GR IV también fue más intensa. A 300 mg kg-1 de NiCl2, GR IV y GR V exhibieron bandas de baja intensidad en las muestras de hojas.
El tratamiento con 50 mg kg-1 de PbAc2 en las raíces indujo la supresión de GR V. En las hojas, la GR I estaba ausente en todos los tratamientos y también en el control, el empleo de 100 mg kg-1 redujo la intensidad de GR IV y GR V, mientras la concentración de 300 mg kg-1 provocó la supresión de todas las bandas.
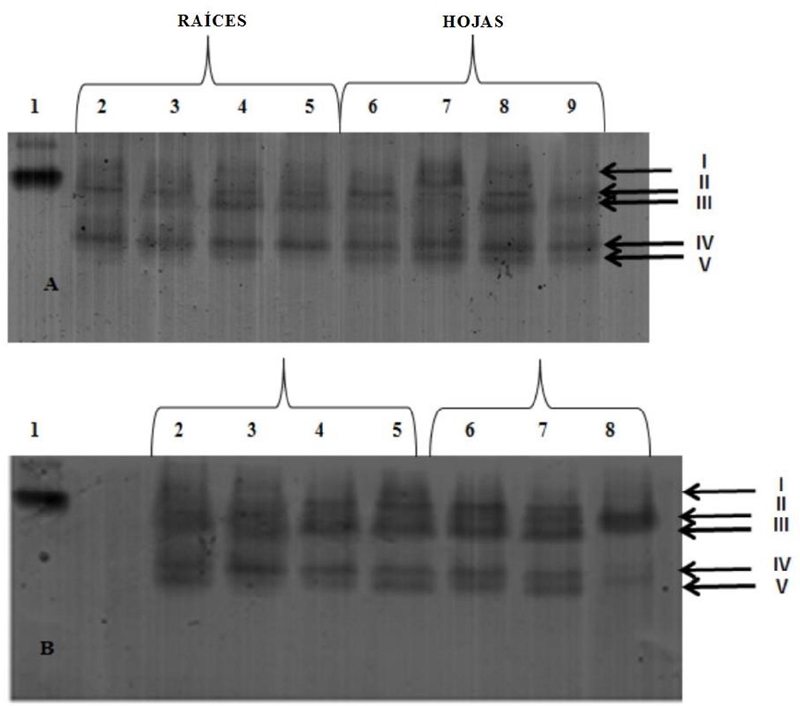
Figura 5 Perfiles isoenzimáticos (GR) en PAGE de plantas de arroz (O. sativa L.) línea 8092. A- níquel (NiCl2), B- plomo (PbAc2). Carril 1= GR patrón de Saccharomyces cerevisiae; Carril 2= control (0 mg kg-1); Carril 3= 50 mg kg-1; Carril 4= 100 mg kg-1; Carril 5= 300 mg kg-1; Carril 6= control (0 mg kg-1); Carril 7= 50 mg kg-1; Carril 8= 100 mg kg-1; Carril 9= 300 mg kg-1. Las flechas indican las isoenzimas GR identificadas.
Discusión
Los metales pesados (Pb y Ni) a concentraciones de 100 y 300 mg kg-1 causaron reducción en el normal desarrollo de las plantas de arroz, un síntoma común que ha sido informado por varios autores (Dogan et al., 2009; Maestri et al., 2010). Este daño se relaciona con la baja actividad de SOD causada por ambos metales en las concentraciones más altas (100 y 300 mg kg-1) en las hojas, lo que indica que el sistema de defensa de las plantas de arroz se ve afectado con estas concentraciones, mostrando debilidades en la vía de eliminación de superóxidos y H2O2 debido al estrés por Ni y Pb. Se ha demostrado que la SOD es la primera línea de defensa contra las ROS, porque esta enzima participa en su detoxificación (Elstner, 1991).
Maheshwari y Dubey (2009) observaron aumentos en las actividades de todas las isoformas de SOD en plantas de arroz sometidas a altas concentraciones de Ni (400 μM). Las SOD en plantas tratadas con Ni deben conferir defensas contra el daño oxidativo, en esta investigación se observaron aumentos en la actividad de SOD en la mayor concentración de Ni y Pb (100 y 300 mg kg-1) en las raíces, en contra de lo que ocurre en trigo y guisante (Gajewska y Sklodowska, 2008).
Cai-Lin et al. (2013) estudiaron los cambios en la actividad de la SOD con diferentes concentraciones de Cd2+, Cu2+ y Hg2+ en plantas florecidas de arroz encontrando que esta actividad fue mayor en las raíces, seguida de la panícula y por último de las hojas. En otro estudio realizado en cuatro cultivares de arroz (S17, HA63, T3028, S8258) utilizando cadmio (50 y 250 µM) se analizó la actividad de diferentes enzimas antioxidantes (POD-peroxidasa, SOD, CAT-catalasas, entre otras) en los tejidos de raíces y hojas y se detectó un aumento en la actividad de la mayoría de las enzimas, entre ellas la SOD en ambos tejidos analizados (Wang et al., 2013).
Tres isoenzimas SOD según su cofactor metálico han sido informadas (Mn-SOD, Fe-SOD y CuZn- SOD) en cultivos como Olea europea L. (Corpas et al., 2006), Nicotiana tabacum L. (Pompeu et al., 2008), Lepidium sativum L. (Manaa et al., 2014) y otros. Estas isoenzimas tienen una distribución heterogénea entre las especies de plantas superiores, tal vez porque su función específica depende de su localización (Corpas et al., 2006).
En arroz sometido a estrés por cadmio, la intensidad de las isoformas de SOD se incrementa en las concentraciones más bajas del metal, pero disminuye en la medida que la concentración aumenta, sobre todo si se combina este factor con el estrés térmico (Shah y Nahakpam, 2012).
El alto nivel de actividad de SOD observado en el control puede ser indicativo de un estrés inverso (debido a la ausencia de Ni). Esto puede explicarse porque el Ni es un metal importante en varios procesos metabólicos, como la ureólisis, el metabolismo del hidrógeno y algunos otros (Mulrooney y Hausinger, 2003).
En los tratamientos con PbAc2, las raíces mantuvieron una mayor actividad de SOD que las hojas, especialmente a 50 mg kg-1. Verma y Dubey (2003) obtuvieron resultados similares en plantas de arroz en crecimiento, en las que la actividad de SOD era más alta en las raíces que en los brotes.
El Pb se acumula principalmente en los tejidos de las raíces, porque es la primera barrera para esta translocación de metales (Blaylock y Huang, 2000), pero puede concentrarse en los tejidos del floema y moverse a través del xilema hacia las hojas (Pinho y Ladeiro, 2012).
Las alteraciones de la actividad de la SOD en el arroz, como en otras especies, se asocian al estrés abióticos de varios tipos, entre ellos los provocados por metales pesados como el manganeso (Srivastava y Dubey, 2011), el cadmio (Shah y Nahakpam, 2012), el cobre (Thounaojam et al., 2012), el arsénico (Tripathi et al., 2013) y el aluminio (Bhoomika et al., 2013).
En las raíces, la intensidad de las bandas de GR fue similar en todos los casos, independientemente del metal utilizado. Ching-Lin y Ching-Hue (2005) no encontraron efecto de la aplicación de Ni en la actividad de GR en raíces de plántulas de arroz. En este estudio, el Ni indujo aumentos en la intensidad de las bandas de las muestras de hojas, particularmente a 50 y 100 mg kg-1, indicando estrés oxidativo. Por el contrario, el tratamiento con plomo a 100 mg kg-1 provocó una disminución en la intensidad de las bandas en las hojas y a 300 mg kg-1 no se observó ninguna banda.
Las isoenzimas GR mostraron diferencias según el órgano de la planta analizado y la concentración utilizada. Las isoenzimas GR III y IV estuvieron presentes en todas las concentraciones y órganos de la planta analizados. Estas dos isoenzimas son responsables de la mayoría de la actividad de GR (Pompeu et al., 2008).
La cantidad de isoenzimas GR varía según la especie; por ejemplo, en Arabidopsis se encontraron dos GR (Xiang y Oliver, 1998) y en trigo se han identificado cinco (Yannarelli et al., 2007) al igual que en esta investigación. El estrés por cadmio en arroz no incrementa la expresión de los genes que controlan la GR excepto cuando la presencia del metal se asocia al estrés térmico (Chou et al., 2012). En cambio, el cobre incrementa la actividad de estas enzimas (Thounaojam et al., 2012) lo que sugiere que su expresión puede ser selectiva en dependencia del metal presente.
La tinción de GR reveló que GR-V es una isoforma específica de las hojas cuando hay Ni presente. En O. sativa, Wu et al. (2013) han descrito dos isoformas cloroplásticas pero Kaminaka et al. (1998) habían purificado y caracterizado la GR en esta especie, encontrando que aunque la enzima se encuentra predominantemente en los cloroplastos, también se halla en pequeñas cantidades en mitocondrias y citosol.
Conclusiones
La enzima superóxido dismutasa (SOD) mostró diferentes comportamientos en hojas y raíces para los tratamientos con los dos metales pesados, en cambio sólo el NiCl2 provocó incrementos significativos en la actividad de la glutatación reductasa (GR) en las raíces en todas las concentraciones con respecto al control. El PbAc2 no causó diferencias en la actividad de GR con respecto al control en ninguno de los órganos analizados.
Se encontró una isoforma específica de glutatión reductasa (GR-V) en las hojas de las plantas tratadas con NiCl2.

