











 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroducción
El cultivo de Capsicum sp. tiene relevancia cultural y económica en México; en los últimos años, ha comenzado a adquirir una importancia económica considerable en todo el mundo. A pesar de que México es considerado uno de los centros de origen y diversificación del chile, Montes et al. (2010); Pérez-Castañeda et al., 2015) este cultivo se cultiva con éxito en China, Turquía, Indonesia, España y Estados Unidos de América; según la FAO (2014), estos países junto con México son los seis mayores productores de chile en todo el mundo. En 2013, China fue el primer país productor con 15 800 000 t, seguido de México con una producción de 2 294 400 t, lo que equivale al 56 y al 8% de la producción mundial, respectivamente, FAO (2014). También hay especies locales que tienen éxito comercial y su cultivo está protegido por denominación de origen. En particular, para Capsicum chinense Jacq (mejor conocido como chile habanero), el sector agrícola de la Península de Yucatán, México, obtuvo la denominación de origen en 2005 y en 2012, su producción anual alcanzó 2 615 t con un valor de $28 487 070.00 pesos (SIAP, 2012; Moo-Muñoz et al., 2016).
Desafortunadamente, los productores de C. chinense han reportado importantes problemas asociados con su cultivo, como las semillas de baja calidad, la pérdida de viabilidad debido al tiempo de almacenamiento y, como resultado, los altos costos de las semillas que se reflejan en un aumento en los costos de producción Garruña-Hernández (2014); Moo-Muñoz et al. (2016). Algunos autores atribuyen estos problemas a las características fisiológicas de C. chinense Garruña-Hernández (2014), mientras que otros los asocian con un manejo deficiente durante el cultivo o el secado y almacenamiento de las semillas (Moo-Muñoz et al., 2016). Para superar estas desventajas, se han explorado diferentes estrategias: i) aseguramiento de la calidad fisiológica durante el desarrollo de la semilla mediante la optimización del proceso de secado; ii) uso de fitorreguladores para mejorar la producción de frutos viables y, por lo tanto, la viabilidad de las semillas Zárate-García et al. (2014); y iii) analizar el efecto del preacondicionamiento del agua complementado con ácido giberélico (GA3) y ácido abscícico (ABA) para mejorar la germinación y disminuir la pérdida de viabilidad de la semilla (Moo-Muñoz et al., 2016).
La investigación ha demostrado que la calidad de la semilla está determinada genéticamente (procesos fisiológicos de acondicionamiento), aunque hay otros factores involucrados, como la nutrición y la salud de la planta madre y las condiciones de almacenamiento. Sin embargo, dado que la disminución de la viabilidad y la pérdida de la capacidad de germinación aún no están claramente explicadas, es interesante investigar si este comportamiento puede revertirse con tratamientos antes de la siembra que reduzcan el tiempo requerido para la radicación de la protuberancia (germinación) y la emergencia de las plántulas. y que juntos se denominan precondicionamiento (Nicasio-Arzeta et al., 2011). Existen varios métodos de preacondicionamiento que pueden clasificarse en a) osmóticos; b) matricial; c) estrés hídrico (Heydeker et al., 1973; Taylor, 1981; Iqbaar y Ashraf, 2005; Artola et al., 2010; Hacisalhoglu y Ross, 2010; Mavi et al., 2010; Sánchez y Muñoz, 2010; Moo-Muñoz et al., 2016). En el preacondicionamiento por estrés hídrico, la disponibilidad de agua para la germinación se regula limitando la cantidad y/o el tiempo de hidratación (Nicasio-Arzeta et al., 2011).
Estos estudios se centraron en una etapa fenológica específica y no brindan detalles sobre el efecto de los tratamientos evaluados en las etapas de desarrollo posteriores, sino solo en la germinación, emergencia y establecimiento (brote de la primera hoja verdadera), lo que significa que no se ha investigado. Si el efecto de los tratamientos de preacondicionamiento en el desarrollo de la planta es duradero. Dado que una imbibición adecuada permite la reactivación de procesos metabólicos como la activación de enzimas, mecanismos para la reparación de membrana, ADN y proteínas, elongación celular y la aparición de la radícula, se ha informado ampliamente que la disponibilidad de agua es esencial para la germinación de semillas Dubreocq et al. (2000); Maldonado et al. (2003), por lo tanto, las plantas deben sincronizar sus ciclos de crecimiento con un suministro de agua adecuado (Foley y Fenhimoe, 1998; Maldonado et al., 2003). No hay información sobre los requisitos de agua para Capsicum chinense; El objetivo de esta investigación fue determinar si el preacondicionamiento del agua de Capsicum chinense Jacq var. Jaguar es suficiente para mejorar los porcentajes de germinación y emergencia.
Materiales y métodos
Material biológico
Las semillas maduras de Capsicum. chinense, var. Jaguar se obtuvieron del campo experimental de INIFAP Las Huastecas, Tampico, Tamaulipas, México. Las semillas se sembraron en el ciclo productivo de diciembre de 2015 a febrero de 2016. Las semillas fueron tratadas con PIRIMFOS y THIRAM 42% por el proveedor y luego envasadas al vacío en una bolsa oscura. En el laboratorio, las semillas se dividieron en subconjuntos de 300 mg de semillas, se empaquetaron en bolsas de polietileno transparente de 7 x 5 cm y se almacenaron a aproximadamente 25 °C.
Experimento de preacondicionamiento y evaluación de germinación
Primero, realizamos la prueba de germinación utilizando papel de enrollar, según los criterios de ISTA (2016, capítulo 5, subsección 5.6.2.1.1, ‘Between paper’) y notamos que los porcentajes máximos de germinación eran muy bajos. Además, al realizar las evaluaciones morfométricas, las raíces solían lesionarse y, finalmente, era difícil controlar la cantidad de agua suministrada. Luego, optamos por realizar otro método ISTA, ‘Top of paper’ en placas de Petri, la secuencia se ilustra en la Figura 1.

Figura 1 Precondicionamiento de la cápsula de Petri de Capsicum chinenese. Los números se describen en el texto.
El preacondicionamiento y la germinación se realizaron en placas de Petri de 8.5 cm de diámetro (ISTA, 2016 cap. 5 subsección 5.6.2.1.1), con las siguientes modificaciones: dos círculos de grado alimenticio de papel (estraza) se colocaron dentro (1) y se humedecieron con los diferentes volúmenes de agua: 2.5, 3.5, 5, 7.5 y 10 ml (3), se distribuyeron 20 semillas de manera equidistante en este círculo de papel (4). Más tarde, una capa de franela blanca, previamente lavada tres veces con agua destilada y seca, se colocó sobre las semillas (5). El volumen de agua de 7.5 ml se utilizó como control. Este volumen de agua se ajustó al diámetro empleado de la placa de Petri (8.5 cm de diámetro), considerando que Garruña-Hernández (2014) empleó el volumen de 8 ml en la placa de Petri de 9 cm de diámetro. Finalmente, las placas se sellaron con una capa de película de poli (cloruro de vinilo) de calidad alimentaria (Egapack®) (6) y se incubaron en una cámara de germinación durante 4 días a 29 ±0.5 °C, con control de fotoperíodo a 12 h de luz/12 h de oscuridad (7). Desde el día 4 después del inicio de la imbibición (DAIS), el sello se abrió rápida y cuidadosamente, para verificar la germinación diaria.
Se evaluaron cuatro variables en los experimentos de preacondicionamiento: 1) porcentaje de germinación acumulativa; 2) porcentaje acumulativo de emergencia; 3) longitud de la radícula; y 4) período de latencia de la semilla (días transcurridos hasta la primera semilla germinada).
La germinación acumulada se registró diariamente hasta el día 9 después del inicio de la imbibición (9 DAIS) porque en el experimento piloto, cuando el porcentaje de germinación en 7.5 ml fue similar al reportado por Garruña-Hernández (2014), el tratamiento rápido se completó en la emergencia dentro de placa de Petri. El criterio de germinación fue que la raíz saliente alcanzó 1 mm de largo. Las semillas germinadas a 9 DAIS se trasplantaron a macetas de poliestireno con 5 g de Miracle-Gro (The Scotts Lawn Company, Ohio, EE. UU.), 0.21 N -0.11 P -0.16 K. Las semillas se colocaron a una profundidad de 2.5 cm y las macetas se incubaron en una cámara de crecimiento en las condiciones descritas anteriormente, se riega diariamente con 3 ml de agua destilada y se observa durante los siguientes 10, 13, 14 y 15 DAIS, para registrar la aparición de los cotiledones del sustrato. Se realizaron tres réplicas independientes con tres repeticiones cada una para cada volumen evaluado (n= 9). Los valores cuantitativos se calcularon de la siguiente manera:
Capacidad total de germinación o porcentaje total acumulado de germinación (CTG%).
Donde: nFPR= radícula saliente final a los 9 días después del inicio de la imbibición (DAIS); n= número de semillas utilizadas por repetición.
Tiempo inferido para alcanzar 50% de germinación (GT50) y 90% de germinación (GT90). Se calculó mediante la prueba PROBIT utilizando el valor de germinación acumulativa.
Porcentaje total de emergencia o porcentaje acumulado de emergencia (CE%).
Donde: nTSEC= número de plántulas totales con cotiledones emergidos a 15 DAIS; n= número de semillas utilizadas por repetición.
Momento en que ha emergido 90% de las plántulas (ET90). Se calculó mediante la prueba PROBIT utilizando los valores de emergencia acumulados.
Longitud acumulada de la radícula (CRL).
Donde:
Los valores de semillas germinadas por placa de Petri se convirtieron a porcentaje considerando 20 semillas como 100%. Los valores del porcentaje de germinación acumulativa se graficaron y analizaron para determinar las diferencias significativas entre los tratamientos.
Se utilizó un diseño experimental completamente al azar. Los datos se analizaron de una manera Anova seguido de la prueba de Tukey. Se calculó el coeficiente de correlación de Pearson en 9 DAIS, p≤ 0.05. Todos los análisis estadísticos se realizaron en el software Minitab® (Minitab Inc. 2007), versión 15 para Windows, State College, Pennsylvania, EE. UU. Los gráficos se hicieron utilizando Office Excel 2016 (ver. 1611).
Resultados y discusión
El conjunto de semillas utilizado para el estudio se evaluó de acuerdo con los criterios ISTA; todas las repeticiones alcanzaron más del 70% de germinación; por lo tanto, se consideraron adecuados para los ensayos de Garruña-Hernández et al. (2014). En el presente trabajo, estudiamos el efecto del preacondicionamiento del agua en la germinación y emergencia de las semillas; el porcentaje de germinación se cuantificó en placas de Petri, mientras que el porcentaje de emergencia se determinó después de que las semillas germinadas se trasplantaron en macetas con sustrato (Figura 2).
Figura 2 Estrategia experimental. Línea de tiempo que indica los días después del inicio de la imbibición (DAIS). La germinación se evaluó diariamente hasta el trasplante a 9 DAIS y la emergencia hasta 15 DAIS.
La germinación se cuantificó hasta las 9 DAIS, ya que en un ensayo piloto se observó que en este momento, las semillas de dos tratamientos alcanzaron al menos 70% de la germinación. Los resultados del porcentaje de germinación acumulativa con respecto al tiempo se muestran en la Figura 3a, donde se puede observar que 2.5 y 3.5 ml de agua de imbibición aumentaron significativamente (p≤ 0.05) el porcentaje de germinación acumulada en comparación con 7.5 ml (volumen de referencia). Esta diferencia aparece desde el cuarto día después del inicio de la imbibición y, excepto en este momento, no hubo diferencias significativas entre 2.5 y 3.5 ml (p˃ 0.05). La Figura 3b muestra los resultados del porcentaje de germinación acumulativa en respuesta al acondicionamiento con agua con 5 y 10 ml en comparación con 7.5 ml (volumen de referencia). El porcentaje de germinación acumulativa aumentó solo en el tratamiento de imbibición de agua de 5 ml desde 7 DAIS (p≤ 0.05).
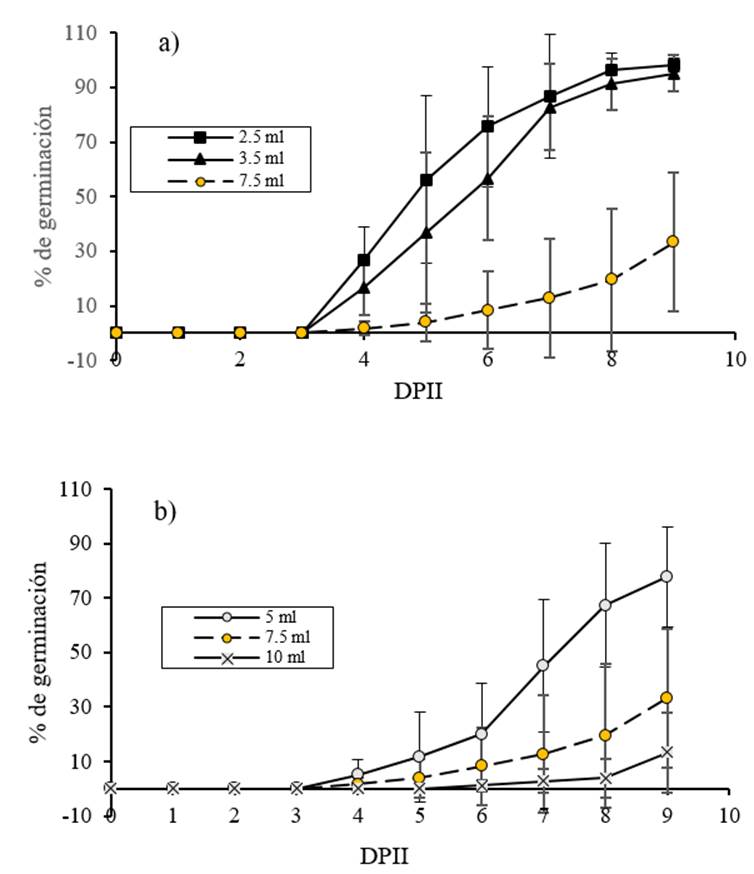
Figura 3 Efecto de diferentes volúmenes de imbibición de agua sobre la germinación de C. chinense. a) las semillas se embebieron con 2.5 y 3.5 ml en comparación con el volumen de referencia (7.5 ml); y b) las semillas se embebieron con 5 y 10 ml en comparación con el volumen de referencia (7.5 ml). La germinación se evaluó diariamente después del inicio de la imbibición.
La capacidad de germinación o el porcentaje máximo acumulado de germinación a 9 DAIS mostró una relación inversamente proporcional a la cantidad de agua utilizada para la imbibición de semillas. El coeficiente de correlación de Pearson fue -0.905, p= 0.000. No hubo diferencias significativas en el porcentaje máximo de germinación acumulativa a 9 DAIS entre los tratamientos 2.5, 3.5 y 5 ml de agua (p˃ 0.05), aunque mostraron diferentes promedios que pueden ser debidos a la alta variabilidad biológica inherente a las semillas de Capsicum chinense (SIPRUFO, 2009; Zárate-García y Sánchez-Azcorra, 2014; Garruña-Hernández et al., 2014; Moo-Muñoz et al., 2016). Además, hubo una disminución significativa en el porcentaje máximo de germinación acumulativa en 10 ml con respecto a 7.5 ml (el volumen de referencia) a 9 DAIS (p≤ 0.05) (Cuadro 1). En la Figura 2, se puede observar que a medida que aumenta el volumen de imbibición, también aumenta la variabilidad, lo que no permite demostrar estadísticamente las diferencias observadas entre todos los resultados.
Cuadro 1 Capacidad de germinación a los 9 DAIS y período de latencia que muestra la desviación estándar, así como comparaciones de prueba de Tukey en pares.
| Volumen de preacondicionamiento | N | Capacidad de germinación o porcentaje de germinación final* |
Período de latencia* |
Inferido GT50 (DAIS)** |
|---|---|---|---|---|
| 2.5 | 9 | 98.3 ±2.5 A | 3.2 ±0.4 A | 5.19 ±1.4 A |
| 3.5 | 9 | 95 ±6.6 A | 3.7 ±2.5 A | 5.78 ±1.6 A |
| 5 | 9 | 78 ±18.3 A | 5 ±1.2 A | 7.35 ±1.8 AB |
| 7.5 | 9 | 33.3 ±25.5 B | 7 ±2.3 B | 9.78 ±2.3BC |
| 10 | 9 | 13.3 ±14.8 B | 8.7 ±1.9 B | 11.19 ±2.1C |
*= las medias con letras diferentes son significativamente diferentes, p≤ 0.05; **= datos inferidos por el método Probit utilizando MInitab.
Las radículas de las plántulas se midieron durante la imbibición mostrando resultados similares al patrón de germinación; el estrés por agua limitada favoreció el aumento en la longitud de la radícula (Figura 4), con un coeficiente de correlación de Pearson a 9 DAIS de -085, p= 0.001. Sin embargo, el tratamiento con 2.5 ml, hay una disminución en los valores de longitud en 9 DAIS, lo que puede indicar que una disminución excesiva del agua afecta el crecimiento. Estos resultados podrían explicarse, ya que las raíces son el órgano clave para la adaptación a la sequía Maldonado et al. (2003); Foolad, (2007); Florido y Fundora (2014) y por lo tanto, el crecimiento y arquitectura responden a las necesidades de los órganos aéreos. Heydecker et al. (1973); Petruzelli et al. (2003). En otras especies vegetales, se ha determinado que un aumento temprano en la longitud de la raíz es un indicador de resistencia al estrés (Florido y Fundora, 1998; Farooq et al., 2008).
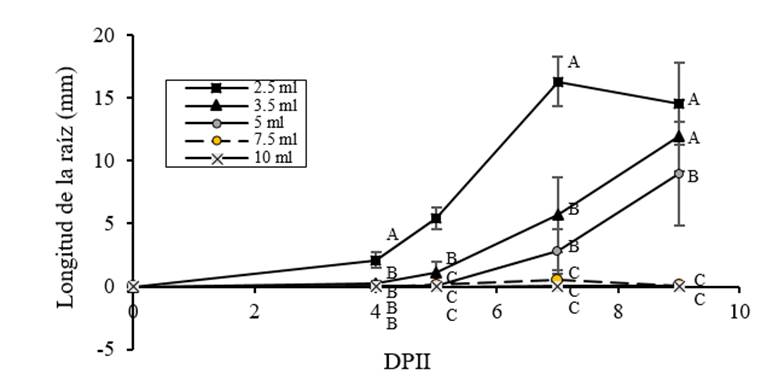
Figura 4 Efecto de diferentes volúmenes de imbibición de agua en la longitud de la radícula de C. chinense. Llas semillas se embebieron con 2.5, 3.5, 5, 7.5 y 10 ml de agua destilada y la longitud de la radícula se midió a 4, 5, 7 y 9 DAIS. El volumen de referencia fue de 7.5 ml.
Para determinar el efecto del volumen de agua en la tasa de germinación, el GT50 se calculó utilizando el método Probit que permite hacer inferencias, asumiendo una distribución normal de los datos. Este enfoque se adoptó por dos razones: a) el aumento del volumen de imbibición produce una mayor variabilidad en los datos; y b) el porcentaje de germinación de 7.5 y 10 ml no alcanzó 50% de germinación en el tiempo evaluado. Los resultados obtenidos permitieron calcular el tiempo necesario para alcanzar 50% de germinación para cada tratamiento de imbibición (Cuadro 1). El volumen de referencia (7.5 ml) tuvo un GT50 de 9.78 días; los volúmenes de 2.5 y 3.5 ml disminuyeron significativamente este tiempo (5.19 y 5.78 días, respectivamente, p≤ 0.05), aunque no hubo diferencias significativas entre ellos, mientras que 5 ml no mostraron diferencias significativas en comparación con 7.5 ml. Por el contrario, 10 ml aumentaron significativamente el GT50 (11.19 días). Estos resultados muestran que el estrés por agua limitada, aumenta la tasa de germinación.
Además, el período de latencia en semillas embebidas en volúmenes bajos se reduce significativamente en comparación con el volumen de referencia y 10 ml. El efecto observado es similar a los resultados obtenidos en otros modelos, donde el preacondicionamiento de las semillas incrementó la tasa de germinación y el porcentaje final (Mc Donald, 2000; Foolad, 2007; Labate et al., 2007; Morandi-Dezfulli et al., 2008).
En trabajos anteriores, se menciona que las condiciones de estrés relacionadas con la disponibilidad de agua favorecen la germinación de Capsicum chinense (Garruña-Hernández et al., 2014), debido a características genéticas que permiten a las semillas adaptarse a limitaciones de agua (Maldonado et al., 2003). En otros modelos, como algunas variedades de Zea mays y Helianthus annuus L., una limitación controlada de agua favorece la tasa de germinación tanto con osmóticos como con preacondicionamiento del agua (Dubreuq et al., 2000; Mc Donald, 2000; Ramón y Mendoza, 2002; Rojo, 2005; Zhang et al., 2007; Farooq et al., 2008; Guan et al., 2009; FAOSTAT, 2014).
En general, la tendencia de los parámetros evaluados muestra que aumentar la imbibición del agua aumenta el tiempo de germinación, el período de latencia y el GT50 (Tabla 1). Además, otros factores en el sistema deben ser considerados; por ejemplo, oxígeno, que es esencial para el metabolismo aerobio de las semillas (Prisco et al., 1992; Koorneef et al., 2002; Ramón y Mendoza, 2002) y que podría estar menos disponible al aumentar el volumen de agua, formando una película más gruesa que disminuye difusión de este gas. Además, es importante tener en cuenta algunos elementos de la cubierta de la semilla que cuando se remojan en agua dificultan la entrada de oxígeno al embrión (Rojo, 2005; Doria, 2010).
Otra variable importante a considerar es que el sistema utilizado en el presente trabajo está sellado con una película de cloruro de polivinilo, y esto también debe tomarse en cuenta para la acumulación de gases Mavi et al (2010) que son productos del metabolismo de las semillas, como el etileno gassemiano que está estrechamente relacionada con la germinación cuando es inhibida por ABA (Beaudoin et al., 2000). En este sentido, se ha informado que el etileno promueve la germinación de las semillas al antagonizar con ABA (Beaudoin et al., 2000; Limkies et al., 2009; Ghassemian et al., 2014). Además, el etileno induce la expresión de genes que codifican enzimas relacionadas con la protuberancia de la radícula, como la β-1,3-glucanasa (Petruzzelli et al., 2003).
Por otro lado, con respecto al promedio del porcentaje de emergencia acumulativa (Figura 5), se puede observar que los valores más altos corresponden a los volúmenes más bajos (3.5, 5 y 2.5 ml, respectivamente) en comparación con el volumen de referencia (7.5 ml), mientras que Los valores más bajos corresponden a 10 ml. También se observó que cuando el volumen de imbibición aumentaba, la dispersión de los datos también aumentaba a medida que se producía la germinación. Debido a este aumento en la dispersión, solo se encontraron diferencias significativas en 13 DAIS. El coeficiente de correlación de Pearson fue de -0.641, p= 0.01 para la distribución de los valores del porcentaje de emergencia frente a los volúmenes de imbibición. Se puede observar que los tratamientos con 3.5 y 5 ml se distribuyen por encima de la línea de regresión, mientras que los datos de 2.5 ml del tratamiento se distribuyen por debajo de la línea de regresión. Los datos del volumen de referencia (7.5 ml) y el tratamiento de 10 ml tuvieron una gran dispersión.
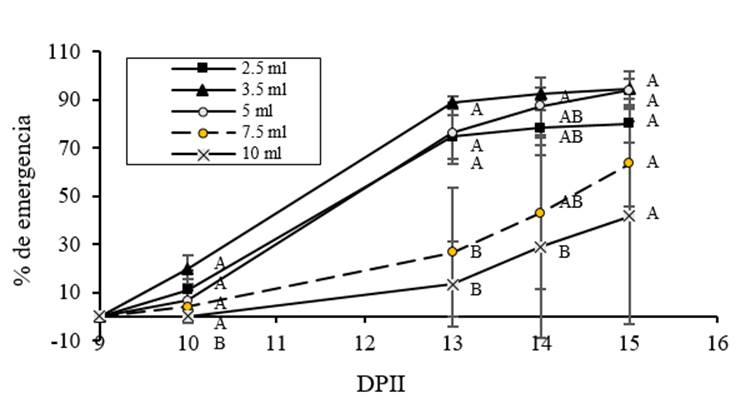
Figura 5 Efecto del volumen de imbibición sobre la emergencia de C. chinense. Las semillas embebidas con 2.5, 3.5, 5 y 10 ml de agua y el volumen de referencia (7.5 ml) se trasplantaron al suelo y la emergencia de la planta se evaluó 1, 4, 5 y 6 días después del trasplante.
Es importante notar que a pesar de que 2.5 ml fue el mejor tratamiento para la germinación, al analizar la emergencia, este tratamiento ocupó el tercer lugar. Este comportamiento puede sugerir que, aunque el estrés hídrico limitado favorece la germinación, cuando es excesivo, tiene un efecto negativo en la emergencia (Heydecker et al., 1973; Sánchez et al., 2001; Rojo, 2005; Stepunh y Raney, 2005; Moradi-Dezfulli et al., 2008; Doria et al., 2010; Nicasio-Arzeta et al., 2011; Florido and Fundora, 2014).
Finalmente, GT90 y ET90 se calcularon mediante el método Probit utilizando el software Minitab. Se trazó un gráfico de barras apiladas con estos valores, para comparar la suma de los tiempos estimados totales en los que se alcanzaría el 90% de emergencia después de trasplantar semillas con 90% de germinación (Figura 6). El tiempo total para las semillas embebidas con el volumen de referencia (7.5 ml) fue de 22.6 días, mientras que para 3,5 ml fue de 13 días, lo que corresponde a aproximadamente la mitad del tiempo (0.59 veces). La suma de GT90 y ET90 de las semillas embebidas con 2.5, 5 y 10 ml fueron 15, 16 y 24 días, respectivamente, y corresponden a 0.68, 0.72 y 1.09 veces la suma de las semillas tratadas con el volumen de referencia. Los resultados muestran que los tratamientos con 3.5 y 5 ml, en ese orden, favorecen la emergencia y aunque 2.5 ml fue el mejor tratamiento para estimular la germinación, ocupó el tercer lugar para la emergencia. Por lo tanto, se puede concluir que una mayor tasa de germinación no necesariamente produce una mejora en el porcentaje de emergencia.
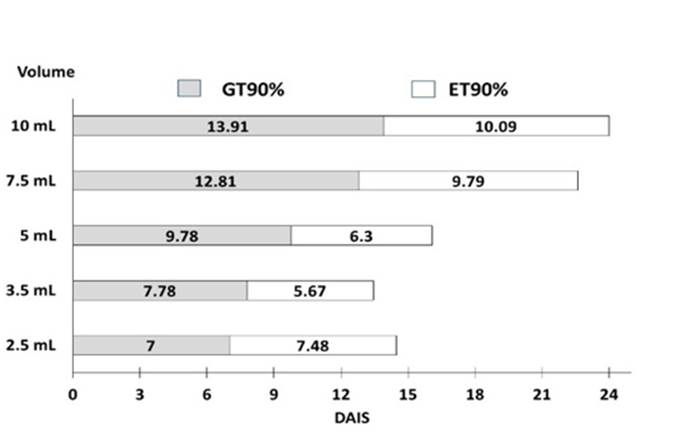
Conclusiones
El preacondicionamiento del agua en las semillas de C. chinense fue suficiente para estimular la germinación y disminuir el GT50 y el tiempo de latencia de forma inversamente proporcional, es decir, el mayor porcentaje de germinación y la tasa se obtuvieron con los volúmenes de imbibición de agua más bajos. Sin embargo, cuando se trasplantaron semillas de C. chinense, para analizar la emergencia, no hubo una mayor correlación inversa entre el volumen de imbibición y el porcentaje de emergencia, lo que sugiere que una mayor tasa de germinación inducida por bajos volúmenes de agua de imbibición (2.5 ml), no necesariamente resulta en una mejora en la emergencia.
La contribución de este trabajo fue evidenciar que la correcta hidratación de la semilla tiene efecto en etapas posteriores a la germinación, incluso, sin agregar reguladores de crecimiento o inductores. Además, se observó que el preacondicionamiento del agua disminuye el comportamiento errático (grandes desviaciones estándar) de los parámetros analizados durante la germinación.
Se sugiere en estudios futuros el desarrollo de plántulas de C. chinense, el preacondicionamiento del agua se debe considerar como la primera opción para optimizar las tecnologías de producción, y GT90 y ET90 se utilizan para predecir tiempos óptimos de germinación y emergencia. Para poder asesorar adecuadamente a los productores, es necesario analizar más etapas fenológicas.

