










Servicios Personalizados
Revista
Articulo
 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de ciencias agrícolas
versión impresa ISSN 2007-0934
Rev. Mex. Cienc. Agríc vol.9 no.7 Texcoco sep./nov. 2018 Epub 30-Sep-2020
https://doi.org/10.29312/remexca.v9i7.1253
Articles
Physiological and biochemical affectations in vitroplants of sugarcane in response to hydric and saline stress
1Universidad Veracruzana-Facultad de Ciencias Químicas. Prolongación de Oriente 6 núm. 1009, Orizaba, Veracruz, México. CP. 94340. Tel. 01(272) 7241779. (mpastelin@uv.mx).
The agricultural production is negatively affected by biotic and abiotic stress, being responsible for great economic losses in the world. Sugarcane is the raw material for obtaining cane juice that is transformed into sucrose and the production of second generation ethanol. In the present study, the number of buds and leaves, the concentration of proline, chlorophyll a, b and total and the concentration of sugars in response to hydric and saline stress in two varieties of sugarcane in vitro were evaluated. Individualized buds of 5 cm in length of the MotzMex 91-207 and SP 71-6180 varieties of sugarcane were cultivated in vitro under water stress (PEG 6000) and saline (NaCl). The two varieties of sugar cane analyzed in vitro presented different responses to osmotic stress. The variety MotzMex 91-207 was better than the variety SP 71-6180 to respond to water stress with a greater accumulation of proline (82.34 mg g-1 PS), did not present decrease in the chlorophyll a, b and total content and generated 20.8 buds per explant even under stress conditions. On the other hand, the response to salt stress with 50 mM NaCl in vitro was presented by the variety SP 71-6180, generating a greater accumulation of carbohydrates such as galactose, glucose and mannose.
Keywords: Saccharum; abiotic stress; high resolution liquid chromatography
La producción agrícola es afectada negativamente por estrés biótico y abiótico, siendo responsables de grandes pérdidas económicas en el mundo. La caña de azúcar es la materia prima para obtener jugo de caña que se transforma en sacarosa y en la producción de etanol de segunda generación. En el presente estudio se evaluó el número de brotes y hojas, la concentración de prolina, clorofila a, b y total y la concentración de azúcares como respuesta al estrés hídrico y salino en dos variedades de caña de azúcar in vitro. Brotes individualizados de 5 cm de longitud de las variedades MotzMex 91-207 y SP 71-6180 de caña de azúcar se cultivaron in vitro bajo estrés hídrico (PEG 6000) y salino (NaCl). Las dos variedades de caña de azúcar analizadas in vitro presentaron respuestas diferentes al estrés osmótico. La variedad MotzMex 91-207 fue mejor que la variedad SP 71-6180 para responder al estrés hídrico con una mayor acumulación de prolina (82.34 mg g-1 PS), no presentó disminución en el contenido de clorofilas a, b y total y generó 20.8 brotes por explante aun en condiciones de estrés. Por otro lado, la respuesta al estrés salino con 50 mM NaCl in vitro la presentó la variedad SP 71-6180 al generar una mayor acumulación de carbohidratos como galactosa, glucosa y manosa.
Palabras clave: Saccharum; cromatografía líquida de alta resolución; estrés abiótico
Global warming increases the frequency of extreme weather events such as floods and droughts negatively affecting agricultural productivity. Mexico is the second country in the world most vulnerable to the effects of climate change, which would result in a possible reduction in agricultural production of 30% if the necessary measures are not taken to face it (Moyer, 2010).
The survival of plants under adverse environmental conditions is based on the metabolic and structural changes that arise to adapt to stress (Golldack et al., 2014). Drought and salinity are stressful abiotic factors that impact on the development of the plant and productivity; therefore, they cause large losses in agricultural yield (OIA, 2009; Agarwal et al., 2013).
The responses to abiotic stress in the physiology of the plant are expressed negatively in photosynthesis, through the reduction of leaf area, chlorophyll content, stomatal conductance and carbohydrate synthesis that can lead to the death of the plant (Chávez-Suárez et al., 2015). The pressure of turgor depends on the osmotic potential of the vacuole; therefore, osmoregulation or osmotic adjustment occurs when cells accumulate solutes, such as proline, in response to some environmental stimulus, such as drought or salinity. This allows the pressure of the turgor to remain high to allow various processes such as the decrease in photosynthesis, reduction of transpiration, stomatal closure and reduction of leaf area caused by water stress (Inman-Bamber and Smith, 2005).
Saline stress has different adverse effects on plants ranging from the rapid decrease in the rate of growth and the reduction in the water flow in the roots which causes water stress, to the accumulation of ions such as Na+ and Cl- in the plant cell that cause toxic effects (Munns et al., 2006).
Polyethylene glycol (PEG) is a polymeric alcohol with high water solubility and low toxicity, acts as a non-penetrating osmotic agent by decreasing the water potential of the culture medium, produces water deficiency in plant cells and an imbalance of metabolism in general, widely used in the performance of experimental simulation studies of drought effects in the laboratory (Fontana et al., 2001; Almansouri et al., 2001; Burnett et al., 2005).
Sugarcane is an important industrial crop, Mexico ranks sixth with a production of 6.18 million tons of sugar (CONADESUCA, 2016). The water deficit is the main factor that influences the productivity of sugarcane. It directly affects tillering and stem height, which results in uncertain sucrose production (Reddy et al., 2004; Sugiharto, 2004).
The objective of this work was to evaluate morphological, physiological and biochemical characteristics in two varieties of sugarcane in vitro (MotzMex 91-207 and SP 71-6180) in response to water stress (PEG 6000) and salt stress (NaCl).
Plant material and culture medium
The present investigation was carried out in the Laboratory of Plant Biotechnology and Cryobiology, Faculty of Chemical Sciences, Veracruz University, located in extension of Oriente 6 No. 1009 Orizaba, Veracruz Mexico.
The biological material used was sugarcane plants (Saccharum spp.) of the MotzMex 91-207 varieties (currently released for the commercial field) and SP 71-6180 (variety used as a female parent in the crosses to obtain the genetic seed via sexual).
The culture medium was a 100% MS (Murashige and Skoog, 1962), supplemented with 2% sucrose (w/v), 100 mg L-1 myo-inositol, 50 mg L-1 thiamin, 100 mg L-1 pyridoxine, 50 mg L-1 nicotinic acid, 300 mg L-1 glycine, 100 mg L-1 biotin, 50 mg L-1 arginine and 50 mg L-1 ascorbic acid. The pH of the medium was adjusted to 5.7 ±0.1 and remained in a liquid state during the development of the experiment. 20 mL of liquid culture medium was deposited in vials with a capacity of 150 mL and sterilized in a vertical autoclave (Lab-Tech model LAC5060s, Namyangju, South Korea) at 120 °C for 20 min.
Effect of polyethylene glycol (PEG 6000) and sodium chloride (NaCl) on different physiological responses
Individualized buds of 5 cm in length were deposited in glass bottles with a capacity of 150 mL, to which 20 mL of liquid culture medium was added. Hydric stress was induced by the addition of polyethylene glycol 6000 (PEG) to the culture medium in concentrations of 0, 5, 10 and 15% (to generate osmotic potentials of -0.18, -0.45, -0.65 and -0.80 MPa, respectively). Saline stress was induced with NaCl at 0, 50, 75 and 100 mM.
The treatments established in the in vitro culture medium were maintained in a controlled room incubation room, with fluorescent white light, whose active photosynthetic radiation was 50 μmol m-2 s-1, photoperiod 16 h light and 8 h darkness, fluorescent white light with a flow of photons between 40 and 50 μmol m-2 s-1 at a temperature of 25 ±2 °C for a period of 30 days was used.
Each treatment had five repetitions and each of them consisted of a bottle with three explants. The data obtained were analyzed through an Anova using the statistical package SAS (SAS, 2011).
Analysis of proline content
The concentration of proline was carried out according to the methodology described by Bates et al. (1973). To do this, fresh foliar tissue was used and analyzed in a Hewlett Packard® 8452 spectrophotometer, at 520 nm, using Merck® L-proline for the calibration curve.
Determination of chlorophylls
The chlorophyll a, b and total contents were made by the method described by Harborne (1973) in which the fresh sample was taken and quantified in a spectrophotometer (ThermoFisher®, Genesys 10S UV-VIS; China) with an absorbance of 663 and 645 nm.
Growth variables
After 30 days of its establishment, in the liquid culture medium and with four levels of PEG and NaCl, the plants were extracted to analyze the number of leaves, buds, proline concentration, chlorophyll content and sugar content.
The percentage increase in the shank number was determined, considering as reference the initial height of the shank. The heights were measured with a flexometer considering as initial height 5 cm in all cases.
Analysis of carbohydrates by high performance liquid chromatography (HPLC)
The carbohydrates were determined by high performance liquid chromatography (HPLC). The equipment used was an HPLC brand Dionex® model ICS-3000 equipped with an electrochemical detector, column CarboPac PA1 2 × 250 mm, of the brand ThermoFisher® using water grade HPLC as mobile phase with an isocratic flow of 0.5 mL min-1 and 300 mM NaOH for the electrochemical reaction. The temperature of analysis was 30 °C, with a running time of 80 minutes. The standards used were arabinose, galactose, glucose, xylose and fructose ≥ 95%, Sigma-Aldrich® brand with nine different concentrations (0.1, 0.2, 0.5, 0.8, 1, 2, 3, 4 and 5%), the extraction was performed with 30 mg of plant material, the injection volume was 25 μL and the quantification was performed by means of a calibration curve and external standard.
Effect of PEG and NaCl
The sugarcane vitroplants increased the accumulation of proline (138.78 mg g-1 PMF) when they were subjected to the stress agent PEG 5%, increasing the number of buds (15). Despite the increase in PEG concentration, chlorophyll levels were higher (Table 1).
Table 1 Effect of the stress agent (PEG and NaCl) on the concentrations of proline, chlorophyll, number of buds and leaves of vitroplants of sugarcane.
| Stressful agent | Proline (mg g-1 PMF) | Chlorophyll a | Chlorophyll b | Total chlorophyll | Buds | Leaves | |
|---|---|---|---|---|---|---|---|
| (mg g-1 PMF) | (number) | ||||||
| PEG (%) | |||||||
| 0 | 30.56 ±0.01 c | 0.3 ±0.02 c | 0.25 ±0.02 cd | 0.55 ±0.03 c | 9.9 ±0.55 c | 23.3 ±0.77 ab | |
| 5 | 138.78 ±90.83 a | 0.4 ±0.03 b | 0.35 ±0.02 b | 0.75 ±0.05 b | 15 ±6.35 abc | 32.4 ±14.24 ab | |
| 10 | 35.22 ±5.84 bc | 0.5 ±0.02 a | 0.4 ±0.02 a | 0.9 ±0.03 a | 11.7 ±2.74 bc | 27.7 ±6.9 ab | |
| 15 | 59.53 ±28.08 bc | 0.54 ±0.08 a | 0.4 ±0.04 a | 0.95 ±0.04 a | 9.8 ±1.75 c | 21.6 ±3.94 b | |
| NaCl (mM) | |||||||
| 0 | 30.56 ±0.01 c | 0.3 ±0.02 c | 0.25 ±0.02 cd | 0.55 ±0.03 c | 9.9 ±0.55 c | 23.3 ±0.77 ab | |
| 50 | 53.2 ±1.46 bc | 0.3 ±0.03 c | 0.26 ±0.02 c | 0.56 ±0.04 c | 17.9 ±0.99 a | 34.7 ±7.12 a | |
| 75 | 68.52 ±18.24 bc | 0.22 ±0.02 d | 0.21 ±0.01 d | 0.44 ±0.03 d | 16.1 ±2.3 ab | 31.8 ±3.29 ab | |
| 100 | 97.82 ±16.78 ab | 0.22 ±0.01 d | 0.22 ±0.03 cd | 0.44 ±0.04 d | 13 ±3.51 abc | 29.8 ±5.48 ab | |
Mean ± SD with different letters in each column indicate that there are no significant statistical differences between treatments (p≤ 0.05).
The response to salt stress was presented in the highest levels of NaCl (100 mM), reaching the highest accumulation of proline (97.82 mg g-1 PMF) (Table 1). Chlorophyll levels decreased in response to salt stress. In the same way García et al. (2010), when evaluating two cultivars of Phaseolus in shading, they found a decrease of the relative chlorophyll content and attributed this reduction to the inhibition in the synthesis of precursors of this molecule which, in general, is accentuated as it is prolonged the period of stress.
According to the results obtained in this study, the vitroplants of sugar cane presented a greater susceptibility to salt stress than to water stress. The content of chlorophyll plays an important role in the growth and development of plants (Jahan et al., 2014) and the decrease in its content is the main cause of alterations in the process of photosynthesis (Khairi et al., 2015).
In the cultivation of sugarcane, the degradation of chlorophyll, when exposed to osmotic stress, causes damage to the photosynthetic capacity, since the levels tend to decrease depending on the exposure time, which leads to a categorical damage to the complex protein of photosystems (Cha-Um and Kirdmanee, 2008).
In relation to the changes presented in arabinose, galactose, xylose and mannose, there were no significant differences in terms of the stress agent to which the sugar cane vitroplants were exposed.
Effect of the variety factor
By inducing the in vitro osmotic stress of sugarcane due to the PEG and NaCl factors, there were different behaviors among the varieties.
The highest accumulation of proline (82.34 mg g-1 PS) occurred in the variety MotzMex 91-207 compared to the variety SP 71-6180 (46.21 mg g-1 PS) (Table 2). The chlorophyll contents showed similarity in the two varieties of sugar cane subjected to water stress, there was no significant statistical difference.
Table 2 Effect of the variety factor on the concentrations of proline, chlorophyll, number of buds and leaves of vitroplants of sugarcane under osmotic stress during 30 days.
| Variety | Proline (mg g-1 PS) | Cl a | Cl b | Cl total | Buds | Leaves | |
|---|---|---|---|---|---|---|---|
| (mg g-1 PF) | (number) | ||||||
| MotzMex 91-207 | 82.34 ±30.44a | 0.36 ±0.06a | 0.3 ±0.03a | 0.67 ±0.09a | 15.05 ±1.94a | 32.93 ±3.93a | |
| SP 71-6180 | 46.21 ±8.88b | 0.33 ±0.14a | 0.28 ±0.1a | 0.62 ±0.1a | 10.78 ±1.46b | 23.23 ±2.02b | |
Mean ± SD with different letters in each column indicate significant statistical differences between varieties evaluated (p≤ 0.05).
The accumulation of sugars in the two varieties under study presented different behaviors. The highest concentrations of arabinose, glucose, xylose, mannose and fructose were found in the sugarcane variety SP 71-6180, while the MotzMex variety only showed the highest concentrations in galactose (Figure 1).
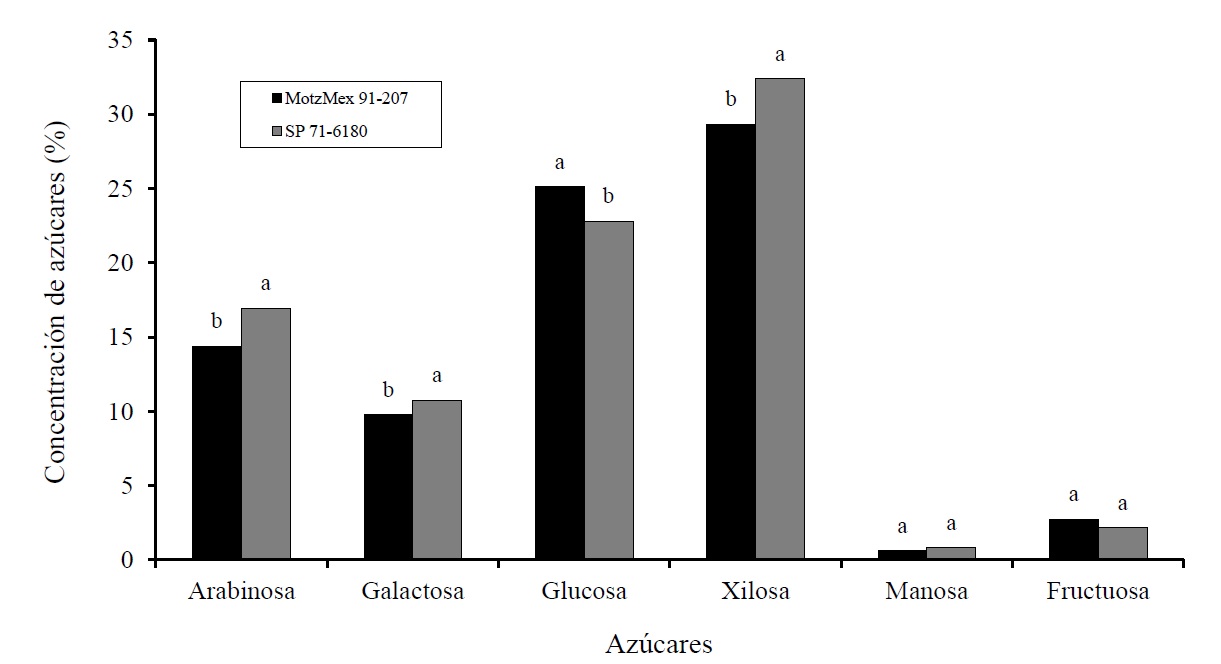
Figure 1 Effect of the variety factor (MotzMex 91-207 and SP 71-6180) on the accumulation of carbohydrates in vitroplants of sugarcane under osmotic stress for 30 days. Different letters on the columns of each figure indicate significant differences between treatments (p≤ 0.05).
The accumulation of proline has frequently been linked to a greater tolerance to salt stress and drought. Although its role in the osmotolerance of the plant remains controversial, it is believed that proline contributes to osmotic adjustment, to the detoxification of reactive oxygen species and to the protection of membrane integrity (Kuznetsov and Schevyakova, 1999; Molinari et al., 2007). In tobacco, transgenic plants have been produced that overproduce proline, improving its tolerance to salts (Kishor et al., 1995). It has been shown that the levels of proline in the roots, although a little lower, are proportional to the foliar concentrations (García and Medina, 2009).
Sugars have functions in plants such as signal transduction of abiotic constraints that have been used to induce changes related to tolerance of plants by transformation with foreign genes and altered expression of the sucrose route, mainly due to modifications in the balance of carbon partition and photosynthesis (Chinnusamy et al., 2005).
Effect of PEG and NaCl interaction
The variety of sugar cane MotzMex 91-207 in concentrations 5% of PEG to the culture medium showed an increase in proline (221.7 mg g-1 PS), being able to regenerate buds even under stressful conditions (20.8 outbreaks) and number of leaves (45.4) (Table 3).
Table 3 Average values for the variables measured in two varieties of sugar cane in vitro (Saccharum spp.) subjected to hydric and saline stress for 30 days.
| Variety | Stressful agent | Proline | Cl a | Cl b | Cl t | Buds | Leaves |
|---|---|---|---|---|---|---|---|
| (mg g-1 PS) | (mg g-1 PF) | (number) | |||||
| PEG (%) | |||||||
| MotzMex 91-207 | 0 | 30.56 ±0.1j | 0.31 ±0.1fg | 0.26 ±0.1ef | 0.58 ±0.1f | 10.4 ±0.1i | 24 ±0.1defg |
| 5 | 221.7 ±0.1a | 0.37 ±0.1e | 0.33 ±0.1d | 0.71 ±0.1e | 20.8 ±0.1a | 45.4 ±0.1a | |
| 10 | 40.55 ±0.1h | 0.51 ±0.1b | 0.41 ±0.1ab | 0.93 ±0.1b | 14.2 ±0.1f | 34 ±0.1bc | |
| 15 | 85.17 ±0.1c | 0.61 ±0.1a | 0.37 ±0.1c | 0.99 ±0.1a | 11.4 ±0.1h | 25.2 ±0.1def | |
| SP 71-6180 | 0 | 30.56 ±0.1j | 0.29 ±0.1gh | 0.23 ±0.1ghi | 0.52 ±0.1g | 9.4 ±0.1k | 22.6 ±0.1efg |
| 5 | 55.86 ±0.1e | 0.42 ±0.1d | 0.36 ±0.1c | 0.79 ±0.1d | 9.2 ±0.1l | 19.4 ±0.1fg | |
| 10 | 29.89 ±0.1k | 0.49 ±0.1bc | 0.39 ±0.1c | 0.88 ±0.1c | 9.2 ±0.1l | 21.4 ±0.1fg | |
| 15 | 33.89 ±0.1i | 0.47 ±0.1c | 0.44 ±0.1ab | 0.92 ±0.1b | 8.2 ±0.1m | 18 ±0.1g | |
| NaCl (mM) | |||||||
| MotzMex 91-207 | 0 | 30.56 ±0.1j | 0.31 ±0.1fg | 0.26 ±0.1ef | 0.58 ±0.1f | 10.4 ±0.1i | 24 ±0.1defg |
| 50 | 51.8 ±0.1g | 0.32 ±0.1f | 0.27 ±0.1ef | 0.6 ±0.1f | 18.8 ±0.1b | 41.2 ±0.1a | |
| 75 | 85.17±0.1c | 0.24 ±0.1i | 0.22 ±0.1hij | 0.46 ±0.1h | 18.2 ±0.1c | 34.8 ±0.1b | |
| 100 | 113.14 ±0.1b | 0.23 ±0.1ij | 0.25 ±0.1efg | 0.48±0.1h | 16.2 ±0.1e | 34.8 ±0.1b | |
| SP 71-6180 | 0 | 30.56 ±0.1j | 0.29 ±0.1gh | 0.23±0.1gh | 0.52 ±0.1g | 9.4 ±0.1k | 22.6 ±0.1efg |
| 50 | 54.53 ±0.1f | 0.28 ±0.1h | 0.24 ±0.1fgh | 0.52 ±0.1g | 17 ±0.1d | 28.2 ±0.1cde | |
| 75 | 51.87 ±0.1g | 0.2 ±0.1j | 0.21 ±0.1ij | 0.41 ±0.1i | 14 ±0.1g | 28.8 ±0.1bcd | |
| 100 | 82.5 ±0.1d | 0.22 ±0.1ij | 0.19 ±0.1j | 0.41 ±0.1i | 9.8 ±0.1j | 24.8 ±0.1def | |
Mean ± SD with different letters in each column indicate that there are no significant statistical differences between treatments (p≤ 0.05).
The MotzMex variety also had the highest content of chlorophyll an and total (0.61 and 0.99 mg g-1 PF) when 15% of PEG was added to the culture medium.
The highest accumulation of xylose (30.53%) was obtained with the variety SP 71-6180 in concentrations of 15% of PEG in the culture medium.
The sugar cane vitroplants of the SP 71-6180 variety accumulated the highest amount of carbohydrates galactose, glucose and mannose under 50 mM NaCl to induce salt stress. The addition of 100 mM NaCl to the culture medium had the highest concentration of mannose (39.2%) in the variety SP 71-61-80 (Table 4).
Table 4 Effect of the interaction of variety and stress agent in the concentrations of carbohydrates of vitroplants of sugarcane.
| Variety | Stressful agent | Galactose | Glucose | Xylose | Mannose | Fructose |
|---|---|---|---|---|---|---|
| (%) | ||||||
| PEG (%) | ||||||
| MotMex 91-207 | 0 | 15.44 ±0.1g | 10.67 ±0.1g | 24.48 ±0.1h | 34.55 ±0.1c | 0.84 ±0.1b |
| 5 | 14.4 ±0.1j | 10.43 ±0.1h | 30.15 ±0.1b | 28.88 ±0.1j | 0.71 ±0.1d | |
| 10 | 13.65 ±0.1l | 9.31 ±0.1l | 25.17 ±0.1f | 29.36 ±0.1h | 0.52 ±0.1f | |
| 15 | 9.77 ±0.1n | 6.54 ±0.1n | 26.64 ±0.1d | 20.49 ±0.1n | 0.42 ±0.1h | |
| SP 71-6180 | 0 | 15.74 ±0.1f | 10.86 ±0.1f | 24.65 ±0.1g | 34.33 ±0.1d | 0.75 ±0.1c |
| 5 | 13.35 ±0.1m | 8.59 ±0.1m | 17.1 ±0.1m | 24.46 ±0.1m | 0.31 ±0.1i | |
| 10 | 16.12 ±0.1e | 11.21 ±0.1c | 19.15 ±0.1j | 29.87 ±0.1g | 0.52 ±0.1f | |
| 15 | 14.08 ±0.1k | 9.61 ±0.1j | 30.53 ±0.1a | 25.24 ±0.1l | 0.43 ±0.1h | |
| NaCl (mM) | ||||||
| MotMex 91-207 | 0 | 15.44 ±0.1g | 10.67 ±0.1g | 24.48 ±0.1h | 34.55 ±0.1c | 0.84 ±0.1b |
| 50 | 16.67 ±0.1d | 11.16 ±0.1d | 25.2 ±0.1f | 30.65 ±0.1f | 0.51 ±0.1fg | |
| 75 | 14.88 ±0.1i | 9.52 ±0.1k | 18.85 ±0.1k | 27.01 ±0.1k | 0.48 ±0.1g | |
| 100 | 15.1 ±0.1h | 9.97 ±0.1l | 25.9 ±0.1e | 29.04 ±0.1l | 0.57 ±0.1e | |
| SP 71-6180 | 0 | 15.74 ±0.1f | 10.86 ±0.1f | 24.65 ±0.1g | 34.33 ±0.1d | 0.75 ±0.1c |
| 50 | 20.61 ±0.1a | 12.12 ±0.1a | 17.79 ±0.1l | 38.25 ±0.1b | 0.41 ±0.1h | |
| 75 | 19.74 ±0.1c | 11.83 ±0.1b | 27.09 ±0.1c | 33.27 ±0.1e | 3 ±0.1a | |
| 100 | 20.29 ±0.1b | 10.92 ±0.1e | 21.24 ±0.1i | 39.29 ±0.1a | 0.3 ±0.1i | |
Mean ± SD with equal letters in each column indicate that in each evaluation there are no significant differences (p≤ 0.05).
Sugar cane is a crop considered moderately sensitive to osmotic stress and what affects its growth and radical morphology, the growth of the aerial system and photosynthetic processes (García and Medina, 2009).
Salinity generates foliar senescence that is accompanied by a remobilization of nutrients, and a consequent decrease in the content of chlorophyll before abscission of the leaf (Keller, 2005; Quesada and Valpuesta, 2008).
The importance of organic solutes, such as sugars, is highlighted in that they are associated with tolerance to salinity (Abdel, 2007). According to Kumar et al. (1994) the high demand for photoassimilates that involves the growth processes of expanding tissues is determined by the ability of the plant to synthesize and accumulate sugars (Pérez-Alfocea et al., 1996). Also, Wahid (2004) found in salinized sugarcane plants, a greater increase in the content of soluble sugars in a tolerant variety, with respect to a sensitive one, which suggests that these solutes play an important role in the osmotic adjustment and in tolerance to salts in this crop.
Ionic homeostasis at the plant level requires precise coordination between cellular mechanisms and those that operate at the intercellular level, in tissues and organs. At the cellular level, the mechanisms of tolerance to salinity rest on several interrelated aspects, such as: the maintenance of osmotic balance in relation to the tonoplast and with the external environment requires the synthesis in the cytoplasm of compatible organic solutes such as sugars (Munns et al., 2006). The roots can perform osmotic adjustment through the accumulation of sugars (Willadino and Camara, 2004, Romero et al., 2004), which decreases the osmotic potential of the root and in this way favors the absorption of water that is limited by salinity. Gupta and Sharma (1990) found that tomato plants treated with 50 mM NaCl levels had a biomass growth similar or superior to the control, in correspondence with an increase in the content of total reducing sugars in the stem and root, it has also been found that salinity affects the dry biomass of the seedlings, but its reduction is lower compared to fresh biomass (Torres and Echevarria, 1994).
Conclusions
The variety MotzMex 91-207 was better than the variety SP 71-6180 under conditions of water stress with the addition of PEG to present a greater accumulation of proline, chlorophyll a, b and total and at the same time continue with its capacity of regeneration of buds and leaves in vitro level. The variety SP 71-6180 was superior in response to salt stress than the MotzMex 91-207, generating a greater accumulation of carbohydrates such as galactose, glucose and mannose.
Literatura citada
Abdel, M. 2007. Physiological aspects of mungbean plant (Vigna radiata L. Wilezek) in response to salt stress and gibberellic acid treatment. Res. J. Agric. Biol. Sci. 3(4):200-213. [ Links ]
Agarwal, P. K.; Shukla, P. S.; Gupta, K. and Jha, B. 2013. Bioengineering for salinity tolerance in plants: state of the art. Mol. Biotechnol. 54(1):102-123. [ Links ]
Almansouri, M. J.; Kinet, M. and Luttss, S. 2001. Effect of salt and osmotic stresses on germination in durum wheat (Triticum durum Desf.). Plant and Soil. 231(2):243-254. [ Links ]
Bates, L. S.; Waldren, R. P. and Teare, I. D. 1973. Rapid determination of free proline for water-stress studies. Plant Soil. 39(1):205-207. [ Links ]
Burnett, S. E.; Pennisi, S. V.; Thomas, P. A. and van Iersel, M. W. 2005. Controlled drought affects morphology and anatomy os Salvia splendens. J. Amer. Soc. Hort. Sci. 130(4):775-781. [ Links ]
Cha, U. S. and Kirdmanee, C. 2008. Effect of osmotic stress on proline accumulation, photosynthetic abilities and growth of sugarcane plantlets (Saccharum officinarum L.). Pakistan J. Bot. 40(6):2544-2552. [ Links ]
Chávez, S. L.; Álvarez, F. A.; Ramírez, F. R.; Infante, F. S.; Licea, C. L.; García, R. B.; García, A. A. y Fonseca, A. M. 2015. Efecto de la salinidad sobre el contenido relativo de agua y la concentración de pigmentos en tres genotipos de frijol. (Phaseolus vulgaris L.) Centro Agricola. 42(3):19-24. [ Links ]
Chinnusamy, V.; Liming, X. and Jian, K. C. 2005. Chapter two: use of genetic engineering and molecular biology approaches for crop inprovement for strees environments. In: Ashraf, M. y Harris, P. (Eds.). Abiotic stresses plant resistance through breeding and molecular approaches. Food Products Press, Binghamton. 47-95 pp. [ Links ]
CONADESUCA. 2016. Diversificación de la caña de azúcar para otros fines. Nota técnica informativa del sector de la caña de azúcar. http://www.gob.mx/cms/uploads/ attachment/file/114368/CAMBIOSNotaABRIL2016.pdf. [ Links ]
Fontana, G.; Licciardi, M.; Mansueto, S.; Schillaci, D. and Giammona, G. 2001. Amoxicillin-loaded polyethylcyanoacrylate nanoparticles: Influence of PEG coating on the particle size, drug release rate and phagocytic uptake. Biomaterial. 22(21):2857-2865. [ Links ]
García, M. y Medina, E. 2009. Acumulación de iones y solutos orgánicos en dos genotipos de caña de azúcar, estresados con sales simples o suplementadas con calcio. Bioagro. 21(1):3-14. [ Links ]
García, M.; García, G. y Sanabria, M. 2010. Efecto de la salinidad sobre el crecimiento, daño oxidativo y concentración foliar de metabolitos secundarios en dos variedades de caraota (Phaseolus vulgaris L.). Interciencia 35(11):840-846. [ Links ]
Golldack, D.; Li, C.; Mohan, H. and Probst, N. 2014. Tolerance to drought and salt stress in plants: Unraveling the signaling networks. Front. Plant Sci. 5(10):151. [ Links ]
Gupta, S. K. and Sharma, S. K. 1990. Response of crops to high exchangeable sodium percentage. Irrigation Sci. 11(3):173-179. [ Links ]
Harborne, J. B. 1973. Phytochemical methods: a guide to modern techniques of plant analysis. Chapman and Hall Ltd, London. 205-207 pp. [ Links ]
Inman, B. N. G. and Smith, D. M. 2005. Water relations in sugarcane and response to water deficits. Field Crops Res. 92(2-3):185-202. [ Links ]
Jahan, M. S.; Nozulaidi, M.; Moneruzzaman, M.; Ainun, A. and Husna N. 2014. Control of plant growth and water loss by a lack of light-harvesting complexes in photosystem-II in Arabidopsis thaliana ch1-1 mutant. Acta Physiol. Plantarum 36(7):1627-1635. [ Links ]
Keller, M. 2005. Deficit irrigation and vine mineral nutrition. Amer. J. Enol. Vitic. 56(3):267-283. [ Links ]
Khairi, M.; Nozulaidi, M. and Sarwar, M. 2015. Effects of different water levels on physiology and yield of salinity rice variety. Australian J. Basic Appl. Sci. 9(2):339-345. [ Links ]
Kishor, K.; Hong, Z.; Miao, G.; Hu, C. and Verma, D. 1995. Overexpression of Pyrroline 5-Carboxylate Syntetase increases proline production and confer osmotolerance in transgenic plants. Plant Physiol. 108(4):1387-1394. [ Links ]
Kumar, S.; Naidu, K. and Sehtiya, H. 1994. Causes of growth reduction in elongating and expanding leaf tissue of sugarcane under saline conditions. Aust. J. Plant Physiol. 21(1):71-83. [ Links ]
Kuznetsov, V. and Schevyakova, N. 1999. Proline under stress: biological role, metabolism and regulation. Russian J. Plant Physiol. 46(2):274-287. [ Links ]
Molinari, H. B. C.; Marur, C. J.; Daros, E.; de Campos, M. K. F.; de Carvalho, J. F. R. P.; Filho, J. C. B.; Pereira, L. F. P.; Vieira, L. G. E. 2007. Evaluation of the stress-inducible production of proline in transgenic sugarcane (Saccharum spp.): osmotic adjustment, chlorophyll fluorescence and oxidative stress. Physiologia Plantarum. 130(2):218-229. [ Links ]
Moyer, M. 2010. How much is left? A graphical accounting of the limits to what one planet can provide. Scientific American - Environment. 74-81 pp. [ Links ]
Munns, R.; James, R. A. and Lauchli, A. 2006. Approaches to increasing the salt tolerance of wheat and other cereals. J. Bot. 57(5):1025-1043. [ Links ]
Murashige, T. and Skoog, F. 1962. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plantarum. 15(3):473-497. [ Links ]
OIA. 2009. Conferencia Internacional. OIA/Egipto ‘World perspectives for sugar crops as food and energy. 23 p. [ Links ]
Pérez, A. F.; Balibrea M.; Santa, C. A. and Estaño, M. 1996. Agronomical and physiological characterization of salinity tolerance in a commercial tomato hybrid. Plant Soil . 180(2):251-257. [ Links ]
Quesada, M. A. y Valpuesta, V. 2008. Senescencia y abscisión. In: fundamentos de fisiología vegetal. Azcón-Bieto, J. y Talón, M. (Eds.). McGraw-Hill Interamericana. Madrid. 559-576 pp. [ Links ]
Reddy, A. R.; Chaitanya, K. V. and Vivekanandan, M. 2004. Drought-induced responses of photosynthesis and antioxidant metabolism in higher plants. J. Plant Physiol. 161(11):1189-1202. [ Links ]
Romero, C. A; Espinosa, R. M. C.; Cutanda, C.; Cortina, P.; Hernández, F. A. y Culiáñez, M. 2004. La osmoregulación: mecanismos y significado. In: la ecofisiología vegetal, una ciencia de síntesis. Thomson Editores. España. Madrid. 603-620 pp. [ Links ]
SAS, 2011. SAS/STAT® 9.3 User’s Guide. SAS Institute Inc. Cary, NC, USA. 178 p. [ Links ]
Sugiharto, B. 2004. Biochemical and molecular studies on sucrose-phosphate synthase and drought inducible-protein in sugarcane (Saccharum officinarum). J. Ilmu. Dasar. 5(1):62-67. [ Links ]
Torres, W. and Echevarría, I. 1994. Germination and seedling growth of rice (Oryza sativa L.) at different NaCl concentration. Cultivos tropicales. 15(2):44-47. [ Links ]
Wahid, A. 2004. Analysis of toxic and osmotic effects of sodium chloride on leaf growth and economic yield of sugarcane. Bot Bull Acad Sin. 45(1):133-141. [ Links ]
Willadino, L. y Camara, T. 2004. Origen y naturaleza de los ambientes salinos. In: la ecofisiología vegetal, una ciencia de síntesis. Reigosa, M.; Pedrol, N. y Sánchez, A. (Eds). Thomson Editores Spain. Madrid. 300-303 pp. [ Links ]
Received: August , 2018; Accepted: October , 2018
 Este es un artículo publicado en acceso abierto bajo una licencia
Creative Commons
Este es un artículo publicado en acceso abierto bajo una licencia
Creative Commons
