











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
El teocintle representa una fuente importante de genes para el mejoramiento del maíz (Zea mays L.) en aspectos de calidad nutritiva, resistencia a enfermedades, adaptación a condiciones de estrés abiótico y calidad agronómica (Cohen y Galinat 1984; Flint-García et al., 2009; Mano y Omori 2007; Wang et al., 2008). La hibridación con maíz se ha documentado en todas las razas y especies de teocintle en México y Guatemala (Wilkes, 1967) híbridos espontáneos se han producido entre el maíz y el teocintle, que ha permitido la transferencia de genes entre ellos (Ellstrand et al., 2007).
El teocintle está representado por especies anuales y perenes, diploides y tetraploides, las cuales se encuentran en áreas tropicales y subtropicales de México, Guatemala, Honduras, Nicaragua, El Salvador y Costa Rica (Reeves, 1950; Sánchez et al., 2011; Sánchez et al., 2018). Las poblaciones de teocintle no tienen una distribución geográfica uniforme y el lugar de su distribución está estrechamente relacionado con el clima, el suelo y actividades antrópicas (Sánchez et al., 2011). Se adapta a elevaciones diversas (Wilkes, 1967; Ruiz et al., 2001), lo que ha propiciado que su desarrollo se produzca bajo diferentes condiciones de temperatura, luz y humedad. Las poblaciones silvestres de teocintle han convivido milenariamente con condiciones climáticas particulares, las cuales probablemente han propiciado diferentes tipos de respuesta a elementos del clima como la temperatura y el fotoperíodo.
Esta condición sugiere la posibilidad de contar con teocintles con diferente nivel de tolerancia a diferentes tipos de estrés abiótico, como la tolerancia a altas y bajas temperaturas. En este sentido, los valores de temperaturas cardinales pueden constituir indicadores de dicha tolerancia, ya que una temperatura umbral máxima elevada o una temperatura umbral mínima baja pudieran estar indicando una planta tolerante a altas temperaturas en el primer caso y a bajas temperaturas en el segundo caso.
No obstante, la importancia que representa el teocintle como pariente silvestre y recurso genético para el maíz, poco se ha avanzado en la caracterización de sus respuestas a las diversas condiciones ambientales. Una de ellas es la relación temperatura-desarrollo, relación que es considerada una de las más determinantes para la adaptación y distribución de las especies (Monteith, 1977; Valladares et al., 2014). En ausencia de sensibilidad al fotoperíodo o vernalización, la tasa de desarrollo de una planta es una función lineal positiva de la temperatura (T) en el intervalo entre la T base (Tb) y la T óptima (To) (Ruiz et al., 2002). La tasa de desarrollo tiene un valor 0 a T iguales o inferiores a Tb, y un valor máximo para To. Arriba de la To, la tasa de desarrollo disminuye hasta una T umbral máxima (Tu). Esto corresponde a una relación sigmoidal entre la T y la tasa de desarrollo de los cultivos (Orchard, 1976; Ruiz et al., 2002). Sin embargo, para datos termo-fenológicos de campo hay dos casos: a) datos de T localizados dentro del intervalo subóptimo; y b) datos de T incluyendo intervalos óptimo y subóptimo (Ruiz et al., 2002). Dentro del intervalo de T subóptimas, la relación tasa de desarrollo-temperatura se describe por la siguiente expresión (Monteith, 1977).
Donde: F= es el tiempo desde el inicio hasta el final del periodo fenológico; T= es la temperatura media del periodo; y ß0 y ß1= son parámetros de una ecuación de regresión lineal simple y equivalen a constantes genotípicas. El valor de Tb es obtenido mediante el cociente -ß0/ß1 (Summerfield et al., citados por Ruiz et al., 2002).
Para datos experimentales que incluyen temperaturas óptimas y subóptimas y describen aproximadamente una nube de puntos con forma no lineal, la determinación de temperaturas umbrales debe hacerse con un modelo curvilíneo. La Tb (umbral mínima), To y Tu (umbral máxima) son conocidas también como temperaturas cardinales del desarrollo y son particulares para cada especie, genotipo y etapa fenológica (Del Pozo et al., 1987; Derieux y Bonhomme, 1990; Durán et al., 2011). Robertson (1983) sugiere que la relación desarrollo-temperatura se describa con una función cuadrática si se quiere determinar el valor de Tb y To, o una función cuadrática-exponencial si se quieren determinar las tres temperaturas cardinales.
El objetivo del presente estudio, fue determinar las temperaturas cardinales de los teocintles Zea perennis, Zea diploperennis, Zea mays subsp. mexicana razas Chalco, Durango, Mesa Central y Nobogame y Zea mays subsp. parviglumis o raza Balsas.
Materiales y métodos
Poblaciones de teocintle
Para tomar en consideración la influencia de la interacción genotipo-ambiente sobre la posible variación de los valores de las temperaturas cardinales, la información fenológica y la determinación de los valores de las temperaturas cardinales se organizó tomando como unidad de estudio las poblaciones de teocintle dentro de cada raza o subespecie. A continuación, se describe cada una de las poblaciones estudiadas utilizando el término de región geográfica.
Zea mays subsp. parviglumis (raza Balsas)
Región 1: comprende las localidades de Zacatongo, en el municipio de Mascota, Jal.; El Tablillo, ubicado en el km 10 carretera Guachinango-La Cienega y km 10 carretera Tepuzhuacán-Zacatongo, ambos pertenecientes al municipio de Guachinango, Jalisco. En el estado de Nayarit incluye la localidad de Amatlán de Cañas en el municipio del mismo nombre. El rango latitudinal de esta población de teocintle va de 20.62861 a 20.85556º norte, mientras que el rango longitudinal abarca de 104.40833 a 104.54861º oeste, con un intervalo altitudinal de 620 a 1 400 m.
Región 2: se ubica en el estado de Guerrero y comprende las localidades de Acahuizotla y el Rincón de la Vía (km 45 carretera 95), en el municipio de Mochitlán y la localidad de Palo Alto en el municipio de Chilpancingo de los Bravos. El rango latitudinal de esta población de teocintle va de 17.28667 a 17.39806º norte, mientras que el rango longitudinal abarca de 99.47306 a 99.4783º oeste, con un intervalo altitudinal de 827 a 1 260 m.
Región 3: también se ubica en el estado de Guerrero y comprende las localidades de Ahuacatitlán (km 1.5 del entronque), Rincón del Sauce y Alcholoa (km 11 carretera Teloloapan-Arcelia) en el municipio de Teloloapan. También comprende la localidad de Ixcateopan (La Tranca) ubicada en el municipio del mismo nombre. El rango latitudinal de esta población de teocintle va de 18.33 a 17.41778º norte, mientras que el rango longitudinal abarca de 99.40778 a 99.92139º oeste, con un intervalo altitudinal de 1 450 a 1 920 m.
Región 4: se localiza en la localidad de El Coyotomate, municipio de Ejutla, en el estado de Jalisco. El rango latitudinal de esta población de teocintle va de 19.9675 a 19.97333º norte, mientras que el rango longitudinal abarca de 104.06861 a 104.58417º oeste, con un intervalo altitudinal de 950 a 1 300 m.
Región 5: se ubica en el Estado de México y comprende las localidades de El Puente (km 22 carretera Luvianos-Zacazonapan, El Puerto (km 18 carretera Luvianos-Zacazonapan) y el sitio ubicado en el km 2 carretera Zacazonapan-Otzoloapan, municipio de Zacazonapan. También comprende la localidad de Sifón Colorines en el municipio de Valle de Bravo. El rango latitudinal de esta población de teocintle va de 19.03889 a 19.18028º norte, mientras que el rango longitudinal abarca de 100.22917 a 100.29258º oeste, con un intervalo altitudinal de 1 085 a 1 468 m.
Región 6: se localiza en el estado de Jalisco y abarca las localidades de La Lima y El Rodeo, municipio de Tolimán. El rango latitudinal de esta población de teocintle va de 19.52417 a 19.52833º norte, mientras que el rango longitudinal abarca de 104.06139 a 104.06389º oeste, con un intervalo altitudinal de 1 450 a 1 460 m.
Región 7: se localiza en el Estado de México en las Ruinas de Malinalco a una latitud de 18.95333º norte, longitud de 99.50278º oeste, y altitud de 1 850 m.
Región 8: se localiza en la localidad de Queretanillo (km 24 carretera Huetamo-Morelia), municipio de Tzitzio, en el estado de Michoacán. El rango latitudinal de esta población de teocintle va de 19.55056 a 19.58333º norte, mientras que el rango longitudinal abarca de 100.91667 a 100.91806º oeste, con un intervalo altitudinal de 1 342 a 1 500 m.
Zea mays subsp. mexicana (raza Chalco)
Región 1: se localiza en San Antonio Soyatzingo, municipio de Amecameca y en Temamatla, ambas en el Estado de México. El rango latitudinal de esta población de teocintle va de 19.08167 a 19.28194º norte, mientras que el rango longitudinal abarca de 98.87167 a 98.88167º oeste, con un intervalo altitudinal de 2 200 a 2 480 m.
Región 2: se localiza principalmente en el municipio de Chalco y en parte del municipio de Temamatla, Estado de México. El rango latitudinal de esta población de teocintle va de 19.18806 a 19.28194º norte, mientras que el rango longitudinal abarca de 98.87167 a 98.88167º oeste, con un intervalo altitudinal de 2 200 a 2 480 m.
Zea mays subsp. mexicana (raza Durango)
Región 1: se ubica en las localidades de Francisco Villa (km 3) y en Puente Dalila (km 7) carretera 45 a Zacatecas, en el estado de Durango. El rango latitudinal de esta población de teocintle va de 24.01917 a 24.08528º norte, mientras que el rango longitudinal abarca de 104.48583 a 104. 56056º oeste, con un intervalo altitudinal de 2 200 a 2 480 m.
Zea mays subsp. mexicana (raza Mesa Central)
Región 1: se ubica en San Jerónimo (km 4 N), municipio de Ayotlán en el estado de Jalisco, a una latitud de 20.41389 a 20.42056º norte, una longitud de 102.34667º oeste y una altitud de 1 550 a 1 600 m.
Región 2: se ubica en Cojumatlán de Régules (km 5.7 SW) en el estado de Michoacán, a una latitud de 20.11389 º norte, longitud de 102.89583 º oeste y una altitud de 1700 m.
Región 3: esta población de teocintle se distribuye en los estados de Michoacán y Guanajuato. En Michoacán se ubica en los sitios San Agustín del Maíz, municipio de Copándaro; (km 2 sur de Cuamio) municipio de Cuitzeo, km 267.5 autopista Guadalajara-Morelia, municipio de Chucándiro; Hacienda La Estancia y en el km 0.5 sur de La Estancia, municipio de Huandacareo. En Guanajuato se ubica en la localidad de Uriangato. El rango latitudinal de esta población de teocintle va de 19.87306 a 20.17º norte, mientras que el rango longitudinal abarca de 101.15611 a 101.38694º oeste, con un intervalo altitudinal de 1 785 a 2 019 m.
Región 4: corresponde al teocintle de la localidad de El Salitre en el municipio de Ixtlán, estado de Michoacán, en los 20.15333-20.15361º latitud norte, 102.36417º longitud oeste y 1 530-1 574 m.
Región 5: se ubica en la localidad de Ciudad Manuel Doblado en el estado de Guanajuato, en los 20.7225º norte, 101.94194º oeste y a 1 710 msnm.
Región 6: abarca los siguientes sitios: Batuecas (km 1 hacia Cuitzeo), km 2 carretera Puruándiro-Cuitzeo, km 3 carretera Puruándiro-Las Pilas, km 2 carretera Puruándiro-Tortugas, km 6 carretera Puruándiro-Villachuato, municipio de Puruándiro, estado de Michoacán.
Región 7: se ubica en el estado de Michoacán e incluye las localidades de Jalapa (km 0.5 SW Jalapa) y el km 1 carretera Zináparo-Churintzio, en el municipio de Zináparo. Además, en el municipio de Churintzio incluye el Cerro Churintzio, Changitiro (km 1 carretera a Purépero), el sitio ubicado en el km 334.5 de la autopista Morelia-Guadalajara, el sitio en el km 0.5 al norte de Churintzio, y el sitio ubicado en el km 3.2 de la carretera Churintzio-La Noria. Latitudinalmente va de 20.16944 a 20.17806º latitud norte y longitudinalmente va de 102.00972 a 102.10694º de longitud oeste, con un intervalo altitudinal de 1801 a 1940 m.
Zea mays subsp. mexicana (raza Nobogame)
Región 1: esta población de teocintle se distribuye en las cercanías del arroyo Tarahumares, y en el transecto entre las poblaciones Tejamanil-La Rinconada, en el municipio de Guadalupe y Calvo, en el estado de Chihuahua. Se ubica en los 26.21694-26.22389º latitud norte, 104.58417-106.9625º oeste, con un intervalo altitudinal de 1 919 a 1 950 m.
Zea diploperennis
Región 1: se distribuye en las localidades de Manantlán, Las Joyas y Corralitos en el municipio de Cuautitlán, Jalisco, con un rango latitudinal de 19.59056 a 19.61667º norte, un rango longitudinal de 104.1159 a 104.30778º oeste y una altitud de 1 854 a 1 870 m.
Zea perennis
Región 1: se distribuye en las localidades de La Tinaja y Piedra Ancha, municipio de San Gabriel, y La Mesa, municipio de Zapotlán El Grande, en el estado de Jalisco. Se ubica en los 19.635-19.64333º latitud norte, 103.56667-103.57722º longitud oeste y a 2 100-2 130 msnm.
Datos de fenología y temperatura
Se utilizaron datos de experimentos de campo realizados entre 1986 y 2005 donde se registraron datos de días a floración (F) y de temperatura (T). En el Cuadro 1 se puede ver la descripción de los ambientes de estudio. La semilla de siembra fue obtenida del Banco de Germoplasma de Maíz del INIFAP y del Colegio de Postgraduados, de los cuales 105 accesiones fueron sembradas durante 1986, 110 durante 1988 y 52 en 1991. Cada accesión fue sembrada en un surco de 10 m con 25 plantas cada uno, bajo un diseño experimental de bloques completos al azar con tres repeticiones.
Cuadro 1 Ambientes de observación fenológica.
| Localidad | Año | Fecha de siembra |
Latitud (N) |
Longitud (W) |
Altitud (m) |
Temperatura media junio-octubre (°C) |
|---|---|---|---|---|---|---|
| Chapingo, México | 1986 | Junio 22 | 19° 29’26 | 98° 53’06 | 2 250 | 17.69 |
| Iguala, Guerrero | 1986 | Junio 22 | 18° 19’06 | 99°32’54 | 767 | 25.91 |
| Las Agujas, Jalisco | 2003 2004 2005 |
Junio 24 Junio 22 Junio 27 |
20° 44’45 20° 44’45 20° 44’45 |
103°30’47 103°30’47 103°30’47 |
1 660 1 660 1 660 |
20.87 21 22 |
| Tepatitlán, Jalisco | 1986 | Junio 22 | 20° 48’35 | 102°45’54 | 1 800 | 19.56 |
| Tlajomulco, Jalisco | 1991 | Junio 22 | 20° 28’25 | 103°26’52 | 1 589 | 20.01 |
| Zapopan, Jalisco | 1988 | Junio 22 | 20°41’40 | 103°23’41 | 1 579 | 21.97 |
Procedimiento de análisis
Para los períodos de floración, en los siete ambientes de observación, se obtuvieron tasas de desarrollo mediante el cálculo de 1/F. Se graficaron estos valores en función de la temperatura para describir la tendencia de los datos. Dado que los datos describían una forma curvilínea, se decidió utilizar el modelo de regresión cuadrática (Robertson, 1983) para calcular la temperatura base y la temperatura óptima:
Donde: Tm es la temperatura media de F. A partir, de esta ecuación la temperatura base se estimó como se expresa en la siguiente expresión:
Donde: Tb= temperatura base; y la temperatura óptima se calculó con la ecuación:
Una vez calculados Tb y To, se determinó la temperatura umbral máxima con la ecuación:
Donde: ln es el logaritmo natural; y C es igual a 1.039. Este valor se obtuvo con un procedimiento de aproximaciones sucesivas.
Resultados y discusión
La Figura 1 muestra la relación observada entre la temperatura y la tasa de desarrollo para los teocintles de las regiones geográficas tipo. Como puede verse en las gráficas de dicha figura, estas dos variables describen una relación curvilínea, por lo que fue posible estimar los valores de las temperaturas cardinales, mediante la metodología planteada. Sin embargo, como puede notarse, el rango de temperaturas bajo el cual se observó el desarrollo del teocintle fue relativamente reducido. No obstante, el rango de temperaturas explorado con los experimentos de campo abarcó casi en todos los casos, niveles térmicos contenidos en los intervalos subóptimo y óptimo para el desarrollo del teocintle. Sólo los casos de los incisos a, d, f y p de la Figura 1, muestran una curva sin la meseta del óptimo térmico, por lo que a futuro deberían procurarse observaciones fenológicas adicionales que generaran mejores modelos. El modelo de regresión cuadrática representó adecuadamente en todos los casos la relación temperatura-tasa de desarrollo. Por el tipo de curva ajustada, fue posible identificar gráficamente la temperatura base y la temperatura óptima; no así la Tu, por lo que fue necesario utilizar un ajuste exponencial.
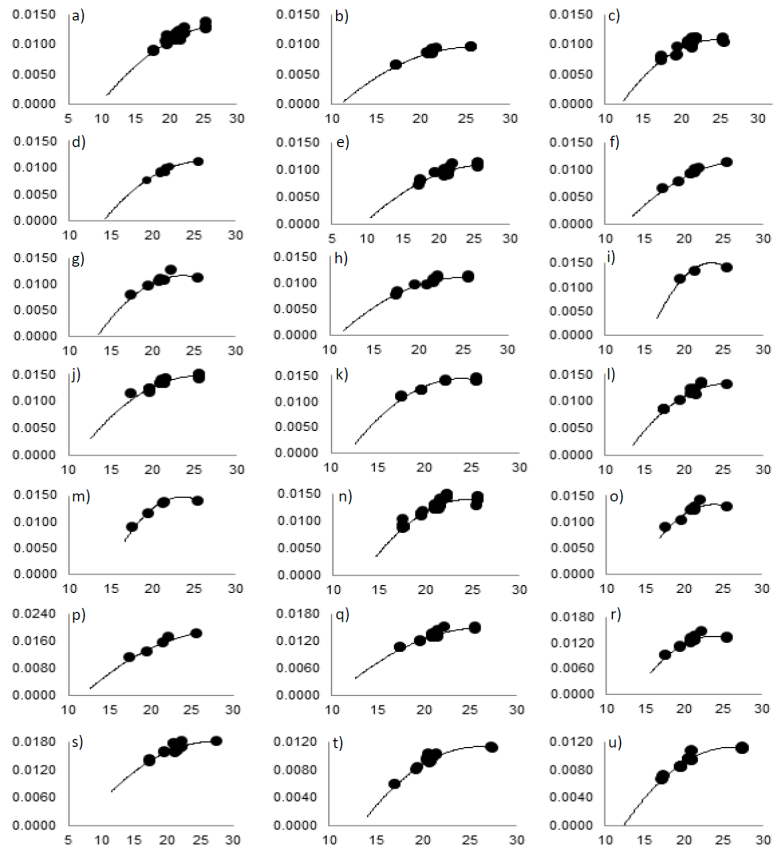
Figura 1 Relación entre temperatura (Eje X) y tasa de desarrollo (Eje Y) en teocintle: a) Balsas Guachinango; b) Balsas Chilpancingo; c) Balsas Teloloapan; d) Balsas Ejutla; e) Balsas Zacazonapan; f) Balsas Tolimán; g) Balsas Malinalco; h) Balsas Tzitzio; i) Chalco Amecameca; j) Chalco Chalco; k) Durango; l) Mesa Central Ayotlán; m) Mesa Central Cojumatlán; n) Mesa Central Chucándiro-Cuitzeo; o) Mesa Central Ixtlán; p) Mesa Central Manuel Doblado; q) Mesa Central Puruándiro; r) Mesa Central Churintzio; s) Nobogame; t) Zea diploperennis; y u) Zea perennis.
En el Cuadro 2, se muestran los valores de los parámetros de la regresión cuadrática y de las temperaturas cardinales obtenidas a través de los modelos generados.
Cuadro 2 Parámetros de regresión y valores de temperaturas cardinales para teocintles.
| Raza/región | Región | ß0 | ß1 | ß2 | Tb | To | Tu | R2 | Tu-Tb |
|---|---|---|---|---|---|---|---|---|---|
| Balsas (subsp. parviglumis) | |||||||||
| Guachinango | 1 | -0.0177 | 0.0022 | -0.00004 | 9.79 | 27.5 | 35.83 | 0.84 | 26.04 |
| Chilpancingo | 2 | -0.0193 | 0.0022 | -0.00004 | 11 | 27.5 | 34.66 | 0.94 | 23.66 |
| Teloloapan | 3 | -0.0316 | 0.0035 | -0.00007 | 11.82 | 25 | 31.29 | 0.82 | 19.47 |
| Ejutla | 4 | -0.0423 | 0.0041 | -0.00008 | 14.31 | 25.62 | 29.43 | 0.93 | 15.12 |
| Zacazonapan | 5 | -0.0149 | 0.0019 | -0.00003 | 9.17 | 31.67 | 40.61 | 0.8 | 31.44 |
| Tolimán | 6 | -0.0266 | 0.0027 | -0.00005 | 12.96 | 27 | 32.15 | 0.98 | 19.19 |
| Malinalco | 7 | -0.0501 | 0.0052 | -0.0001 | 12.77 | 26 | 31.34 | 0.86 | 18.57 |
| Tzitzio | 8 | -0.0236 | 0.0028 | -0.00005 | 10.33 | 28 | 35.78 | 0.91 | 25.45 |
| Chalco (subsp. mexicana) | |||||||||
| Amecameca | 1 | -0.0312 | 0.0036 | -0.00007 | 11.03 | 25.71 | 32.79 | 0.79 | 21.76 |
| Chalco | 2 | -0.0256 | 0.0031 | -0.00006 | 10.31 | 25.83 | 33.63 | 0.82 | 23.32 |
| Durango (subsp. mexicana) | 1 | -0.0419 | 0.0047 | -0.0001 | 11.95 | 23.5 | 31.69 | 0.81 | 19.74 |
| Mesa Central (subsp. mexicana) | |||||||||
| Ayotlán | 1 | -0.0388 | 0.0041 | -0.00008 | 12.52 | 25.62 | 31.22 | 0.93 | 18.7 |
| Cojumatlán | 2 | -0.1056 | 0.0103 | -0.0002 | 14.13 | 25.75 | 29.74 | 0.93 | 15.61 |
| Chucándiro-Cuitzeo | 3 | -0.0518 | 0.0054 | -0.0001 | 12.47 | 27 | 32.64 | 0.86 | 20.17 |
| Ixtlán | 4 | -0.0603 | 0.0062 | -0.0001 | 12.08 | 31 | 37.04 | 0.85 | 24.96 |
| M. Doblado | 5 | -0.0337 | 0.0036 | -0.00006 | 11.6 | 30 | 36.51 | 0.96 | 24.91 |
| Puruándiro | 6 | -0.0254 | 0.003 | -0.00006 | 10.8 | 25 | 32.32 | 0.87 | 21.52 |
| Churintzio | 7 | -0.0584 | 0.006 | -0.0001 | 12.22 | 30 | 35.89 | 0.89 | 23.67 |
| Nobogame (subsp. mexicana) | 1 | -0.0194 | 0.0029 | -0.00006 | 8.02 | 24.17 | 34.26 | 0.73 | 26.24 |
| Zea diploperennis | 1 | -0.0358 | 0.0036 | -0.00007 | 13.47 | 25.71 | 30.36 | 0.95 | 16.89 |
| Zea perennis | 1 | -0.0296 | 0.0031 | -0.00006 | 12.64 | 25.83 | 31.31 | 0.94 | 18.67 |
Tb= temperatura base (°C); To= Temperatura óptima (°C); Tu= temperatura umbral máxima (°C). ß0, ß1; ß2= parámetros de la regresión cuadrática.
Los valores de r2 se ubicaron por arriba de 0.8, con excepción de dos casos en los que el valor de este estadístico se ubicó entre 0.73 y 0.79. El menor ajuste en estos modelos podría estar señalando la influencia de otros factores sobre la fenología del teocintle, como el fotoperíodo (Hung et al., 2012) o la humedad del suelo (Payero et al., 2006), considerando que los datos analizados son de siembras de temporal.
A pesar de que está comprobado que el fotoperíodo ejerce una influencia importante en la duración del ciclo de desarrollo del teocintle, retrasando la floración en ambientes de día largo (Hung et al., 2012), en los datos de campo analizados y considerando el ajuste de los modelos obtenidos, se puede decir que el efecto de la variable fotoperíodo se mantuvo constante en todos los estudios de campo utilizados, ya que todos se desarrollaron en el ciclo primavera-verano.
Como puede verse en el Cuadro 2, tanto los valores de temperatura base, como de temperatura umbral máxima y temperatura óptima varían entre las poblaciones de teocintle estudiadas. Esto muestra una capacidad de adaptación térmica variante entre los teocintles que habitan distintas regiones eco-geográficas, lo cual es producto de la interacción planta-ambiente experimentada por siglos en estas plantas milenarias (Wilkes, 1967). Al comparar los valores de Tb obtenidos para los teocintles con los valores de Tb para las diferentes razas de maíz (Ruiz et al., 1998), es posible notar que el maíz ha logrado una mayor adaptación a ambientes más templados, dado que las razas Apachito, Cristalino de Chihuahua, Azul, Gordo, Palomero de Chihuahua, Palomero Toluqueño, Arrocillo, Cónico, Elotes Cónicos, Cacahuacintle, Chalqueño, Mushito, Serrano de Jalisco, Cónico Norteño, Ancho, Bofo, Elotes Occidentales, Tablilla de Ocho, Zamorano Amarillo, Bolita, Jala, Comiteco, Coscomatepec, Olotón, Ratón, Reventador, Onaveño y Dulcillo muestran una Tb inferior a 8 °C.
Con relación al maíz, cuya temperatura base típica (10 °C) está ampliamente citada (Gilmore y Rogers, 1958; Cross y Zuber, 1972; Neild, 1982; Cutforth y Shaykewich, 1989; Ruiz et al., 2002), los teocintles que mayor semejanza presentaron (10 ±1°C) fueron Balsas Guachinango, Balsas Chilpancingo, Balsas Zacazonapan y Balsas Tzitzio, así como Mesa Central Puruándiro y Chalco Amecameca, por lo que estas poblaciones pueden tener una capacidad de adaptación térmica similar al maíz original que de acuerdo con Matsuoka et al. (2002) se derivó de Zea mays ssp. parviglumis.
La adaptación y desarrollo de las especies vegetales está condicionada por las temperaturas umbrales de desarrollo mínima y máxima, también llamadas ceros fisiológicos. Por debajo del umbral mínimo y por arriba del umbral máximo, la actividad celular y por tanto el desarrollo del organismo se consideran nulos (Ruiz et al., 2002). De acuerdo con esto, el teocintle que tendría actividad celular y por tanto crecimiento y desarrollo a las más bajas temperaturas sería el teocintle Nobogame, cuyo valor de temperatura base es de 8 ºC (Cuadro 2). En tanto los teocintles que tendrían actividad celular y fotosintética a las más altas temperaturas serían los teocintles Balsas Zacazonapan, Mesa Central Ixtlán, Mesa Central Manuel Doblado y Mesa Central Churintzio, con un valor de Tu de 40.61, 37.04, 36.51 y 35.89 ºC, respectivamente (Cuadro 2). Estos mismos teocintles destacan también por tener el valor más alto de temperatura óptima, por arriba de 30 ºC. Estas poblaciones de teocintle podrían indicar la presencia de germoplasma funcional en ambientes cálidos, de altas temperaturas, lo cual podría representar una característica ventajosa del teocintle, adicional a las ya reportadas para el teocintle, como son la adaptación a sequía (Sánchez et al., 2018), ambientes inundados (Mano y Omori, 2015), resistencia a enfermedades (Nault, 1983) y otras más. Estas poblaciones podrían constituir una fuente de genes para incorporar tolerancia a altas temperaturas en programas de mejoramiento genético de maíz.
Los teocintles mexicanos anuales muestran sub-estructuraciones genéticas a lo largo de líneas geográficas (Fukunaga et al., 2005), lo que establece una interacción del taxón con condiciones ambientales variantes, y una presión de selección de plantas (Coop et al., 2010), asegurando que las diversas poblaciones de estos teocintles estén adaptadas a ciertas condiciones particulares de temperatura, precipitación (agua) y radiación solar (insumos ambientales básicos para una planta). Esto puede explicar la variabilidad intra e inter-racial observada en los valores de las temperaturas cardinales estudiadas. Algunos valores altos de Tb en el Cuadro 2 resultan con poca correspondencia con la región geográfica de la población, tal es el caso del teocintle Mesa Central Cojumatlán, con una Tb de 14.13 °C, en una región geográfica a 1 700 m de altitud con un clima semicálido y una temperatura media anual alrededor de los 19 °C. Dado que el teocintle de esta región debe estar aclimatado a estas condiciones ambientales, es conveniente tomar con reservas este valor de Tb y procurar en el futuro cercano hacer un re-análisis incluyendo más datos de fenología de esta población.
Conclusiones
Existen diferencias intra e inter-raciales con relación a los valores de las temperaturas cardinales de las diversas poblaciones de teocintle, lo que señala que el teocintle posee una adaptación a diferentes condiciones de temperatura a través de los tipos de teocintle que se conocen.
Los valores de temperaturas cardinales obtenidos permiten concluir que considerando las diferentes poblaciones de teocintle, los ceros fisiológicos (temperaturas umbrales) van más allá del rango térmico de un maíz típico (Tb= 10 °C; Tu= 30 °C), por lo que existe un potencial interesante de incorporación de temperaturas umbrales del teocintle hacia el maíz, sobre todo considerando mejorar la adaptación de este cultivo a los ambientes de producción cada vez más cálidos que está trayendo consigo el cambio climático.
Es deseable en el futuro mediato incorporar más datos de fenología y temperatura al análisis de temperaturas cardinales de algunas de las poblaciones estudiadas para la obtención de resultados más robustos con relación a la determinación de los valores de las temperaturas cardinales de los teocintle. Sin embargo, los resultados del presente estudio pueden considerarse como un valioso primer reporte cuantitativo acerca del tema de temperaturas cardinales del teocintle.

