










Serviços Personalizados
Journal
Artigo
 texto em
texto em  Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista mexicana de ciencias agrícolas
versão impressa ISSN 2007-0934
Rev. Mex. Cienc. Agríc vol.9 no.6 Texcoco Ago./Set. 2018
https://doi.org/10.29312/remexca.v9i6.1585
Articles
In vitro cultivation of petals of four varieties of Begonia elatior
1Fruit Growing Program-Institute of Genetic Resources and Productivity-Postgraduate College, highway Montecillo-Texcoco km 36.5, Texcoco, State of Mexico. CP. 56230. Tel. 01 (55) 58045900, ext. 1518.
2Department of Phytotechnics-Chapingo Autonomous University. Chapingo, Texcoco, State of Mexico.
This investigation was carried out with the objective of establishing the protocol to know the totipotencia of the petal for the generation of seedling massively by means of the addition, to the culture medium, of an auxin and a cytokinin and to induce the cloning of four varieties of Begonia elatior. Full petals were used that were grown in Murashige and Skoog culture medium in a completely randomized experimental design consisting of five combined levels of ANA and BA (0, 1, 0.5, 2 and 3 mg L-1). The highest regeneration of callus recorded, on average, 57.2%, in petals of the cv ‘Heidi’ and 46.4% in the variety ‘Gloriosa’. These results were given at 65 days of MS medium culture supplemented with 1 mg L-1 of ANA and 1 mg L-1 of BA. The best sprout formation and growth was obtained with MS medium added with 1 mg L-1 of ANA and 1 mg L-1 of BA with 13.2% in the variety ‘Heidi’ and ‘Gloriosa’. Occasionally, floral buds were present in the treatment 1 mg L-1 of ANA + 1 mg L-1 of BA in the cv ‘Heidi’. The root formation was obtained only in the ‘Heidi’ variety and 1 mg L-1 of ANA + 1 mg L-1 of BA was induced in the treatment. It is possible to obtain seedlings from petals of Begonia elatior.
Keywords: auxin; cytokinin; indirect organogenesis
Esta investigación se llevó a cabo con el objetivo de establecer el protocolo para conocer la totipotencia del pétalo para la generación de plántula masivamente mediante la adición, al medio de cultivo, de una auxina y una citocinina e inducir la clonación de cuatro variedades de Begonia elatior. Se emplearon pétalos completos que fueron cultivados en medio de cultivo Murashige y Skoog en un diseño experimental completamente al azar consistente en cinco niveles combinados de ANA y BA (0, 1, 0.5, 2 y 3 mg L-1). La mayor regeneración de callo registró, en promedio, de 57.2%, en pétalos de la cv ‘Heidi’ y de 46.4% en la variedad ‘Gloriosa’. Estos resultados se dieron a los 65 días de cultivo medio MS suplementado con 1 mg L-1 de ANA y 1 mg L-1 de BA. La mejor formación de brotes y crecimiento se obtuvo el medio MS adicionado con 1 mg L-1 de ANA y 1 mg L-1 de BA con 13.2% en la variedad ‘Heidi’ y ‘Gloriosa’. Ocasionalmente se presentaron yemas florales en el tratamiento 1 mg L-1 de ANA+ 1 mg L-1 de BA en la cv ‘Heidi’. La formación de raíz se obtuvo, únicamente, en la variedad ‘Heidi’ y se indujo en el tratamiento 1 mg L-1 de ANA+ 1 mg L-1 de BA. Es posible obtener plántulas a partir de pétalos de Begonia elatior.
Palabras clave: auxina; citocinina; organogénesis indirecta
Introduction
The begoniacea family comprises five genera and 920 species, most of them belonging to the genus Begonia (Mendi et al, 2009; Kabirnataj et al., 2012). The genus begonia comprises approximately 2000 varieties and around 200 species that have been introduced by commercial growers, including Begonia tuberhybrida, B. rex, B. semperflorens, Begonia × hiemalis, Begonia × elatior, Begonia × cheimantha and Begonia × socotrana (Takayama, 1982). The begonias are part of our most beautiful and varied adornment plants. The Begonia has been, for years, the first flower plant in Central Europe and although currently it is not so sold, it maintains a privileged position among the top 10 commercials (Rout et al., 2006).
The begonias are important as perennial ornamental plants that are distributed in the tropical and subtropical regions of the world. They are used in gardening for the implantation of massifs and borders and as pot plants. In general, they are propagated by seed or vegetatively by leaf cutting, rhizomes, tubers and mainly by leaf cutting with petiole (Mendi et al., 2009). However, it is possible that conventional methods of propagation are problematic due to the rapid onset of diseases. In addition, the production of large quantities of genetically homogeneous plants is also very difficult.
Begonia elatior, is the product of the cross of B. socotrana x B. tuberhybrida and is known as B. hiemalis and is considered a flower with great potential because of the great acceptance in local consumption, but with good management in marketing It can be considered for the international market. In the domestic market it is the predominant one since sales indicate that it is around 90% and possibly its demand increases. However, because it is an import product, the cost per cutting is 0.45 €, an approximate equivalent of $10.00 cost which is high. The propagation of Begonia elatior, is important from the commercial point of view. In this situation, a method is required to obtain vegetative material at a low production cost and with high quality.
For the development of this research, micropropagation was used as a tool for the massive propagation of Begonia elatior. For the development of the research, the petal was chosen as explant because although the begonias have been studied by several researchers (Takayama and Misawa,1982; Simmonds and Werry, 1987; Bowes and Curtis, 1991; Nakano et al., 1999; Bouman and Klerk, 2001; Kishimoto et al., 2002; Burritt and Leung, 2003; Espino et al., 2004; Nhut et al., 2005; Shimada et al., 2006, 2007; Mendi et al., 2009; Romocea et al., 2010; Kabirnataj et al., 2012). There is little or no information about the in vitro cultivation of petal of Begonia elatior as mother material, so we think that this type of explant can be interesting and cheap, especially because of the amount of flowers and petals that one has in a plant. In addition, through micropropagation, a high quality plant can be obtained, which is a necessary and critical condition for producers (Chebet et al., 2003). Therefore, the present work was oriented, as an alternative method of mass propagation of Begonia elatior, the establishment of a protocol to know, through hormonal manipulation, the organogenetic potential of the petal for the clonal generation of seedlings.
Materials and methods
The present investigation was carried out in the Biotechnology laboratory (tissue culture section) in the area of Agronomy of the Autonomous University Chapingo (UACH), Texcoco, State of Mexico and in the Embryogenesis laboratory belonging to the Area of Fruit Production, Institute of Genetic Resources and Productivity of the Postgraduate College-Campus Montecillo.
Four varieties of Begonia elatior plants (‘Heidi’, ‘Clara’, ‘Gloriosa’ and ‘Tacora gee’) were used as experimental material, which were purchased in the ‘Madre Selva’ market, Xochimilco delegation, Mexico City. The varieties were ‘Tacora gel’ (yellow petals), ‘Gloriosa’ (orange), ‘Heidi’ (red) and ‘Clara’ (white).
The petals used were extracted from male flowers (2 to 3 cm) located on the second and third floors of the plant structure. Full petals of the four varieties were washed, to remove the dust, with soapy water in running water for 30 min and immersed with 70% alcohol for 20 s, sterilized in 4% calcium hypochlorite with two drops of tween 20 for 15 min. The explants were rinsed three times with sterilized bidistilled water and seeded in culture medium Murashige and Skoog (1962) supplemented with glycine 0.2 mg L-1, nicotinic acid 0.5 mg L-1, myo-inositol 100 mg L-1, pyridoxine 0.5 mg L-1, thiamine-HCl 0.4 mg L-1, 30 g L-1 sucrose, 7.5 g L-1 agar (sigma) and the growth regulators naphthalene acetic acid (ANA) and benzylamino purine (BA) that were added in combination and the witness. The pH of the culture medium was adjusted to 5.7 or 5.8 with 0.1 N NaOH or 0.1 N HCl. The culture medium Murashige and Skoog (MS) was sterilized in an autoclave at 1 kg cm-2 at 120 °C for 15 min. and it was served in gerber-type bottles (20 ml/bottle) with plastic caps. The petals were incubated at a temperature of 26 °C ±2 °C with a photoperiod of 16 h of light and 8 h of darkness and a luminous intensity of 50 μM m-2 s-1 throughout the experiment.
Establishment of the inoculum
Petals of the four varieties were inoculated in the MS culture medium containing concentrations and combinations of BA and ANA in the following treatments: 1) 0 mg L-1 of ANA + 0 mg L-1 of BA; 2) 1 mg L-1 of ANA + 1 mg L-1 of BA; 3) 0.5 mg L-1 of ANA + 2 mg L-1 of BA; 4) 1 mg L-1 of ANA + 2 mg L-1 of BA; and 5) 0.5 mg L-1 of ANA) + 3 mg L-1 of BA.
Multiplication
For the micropropagation, the regenerated seedlings were used, which were subcultured in fresh establishment medium added with the same concentrations of ANA and BA used for the establishment of the explant.
Induction of the root
For the induction of the root, the seedlings were transferred to a medium with treatment 2 (1 mg L-1 ANA + 1 mg L-1 BA).
Experimental design
A completely randomized experimental design was used, with factorial 45 where the factors to be evaluated were: variety and concentration of growth regulators, generating 20 treatments with 10 repetitions resulting in 200 experimental units. The 2 petals were planted per bottle. The statistical analyzes for the variable number of outbreaks, was carried out with the Infostat program (Table 1).
Table 1 Factorial sample: begonia variety and concentration of ANA + BA.
| Factor 1 variety | Factor 2 concentration of ANA and BA |
|---|---|
| MI = 0 mg L-1 (ANA) + 0 mg L-1 (BA) | |
| C1= Clara | MII = 1 mg L-1 (ANA) + 1 mg L-1 (BA) |
| C2= Heidi | MIII = 0.5 mg L-1 (ANA) + 2 mg L-1 (BA) |
| C3= Gloriosa | MIV = 1 mg L-1 (ANA) + 2 mg L-1 (BA) |
| C4 = Tacora geel | MV = 0.5 mg L-1 (ANA) + 3 mg L-1 (BA) |
Letter C corresponds to the varieties of the flower and M to the concentration of the hormones ANA and BA.
Results and discussion
Callus formation
The callus formation occurred at the base of the petal 120 days after sowing. The treatments that tested positive for callus formation were the 2 (1 mg L-1 (ANA + 1 mg L-1 of BA) and treatment 4 (1 mg L-1 of ANA) + 2 mg L-1 of BA). In the variety ‘Heidi’ the value was, on average, 57.2% and in treatment 4 22%; in ‘Gloriosa’, in treatment 4 (1 mg L-1 (ANA) + 2 mg L-1 of BA), an average percentage of 46 was obtained (Figure 1). Treatment 2 agrees with Datta (2002) who reported, in studies carried out in Rosa sp., that the development of calluses from petal explants in seven cultivars (Contempo, America’s Junior Miss, Manasi, Mrinalini, Preyasi, Sylvia and Queen Elizabeth), was satisfactory, for callus formation, in MS medium with 1 mg L-1 of ANA and 1 mg L-1 of BA.
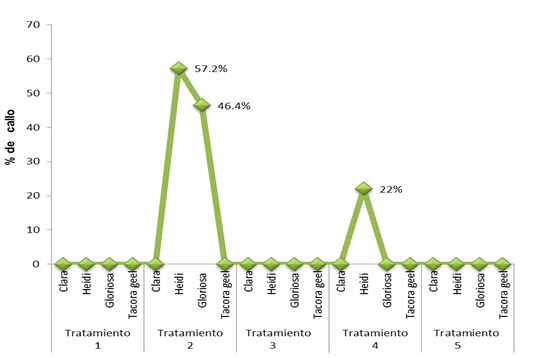
Figure 1 Formation of petal callus expressed as a percentage of four varieties of Begonia elatior grown in treatments 2 and 4 of BA/ANA in MS medium.
The results indicate that the ‘Tacora geel’ and ‘Clara’ varieties were not successful in any treatment. This variation of not callus formation, according to Bonga (1987), can be attributed to the different capacity of regeneration or sensitivity of the petals and that this is depending on the maturity of the explant (petal) in relation to the concentrations of ANA and BA.
Number of petals with bud
The Figure 2 shows the varieties that generated adventitious shoots. It was observed that in the varieties ‘Clara’ and ‘Tacora gel’ there was no type of organogenic response. In contrast, in the ‘Heidi’ variety in treatment 2 (1 mg L-1 of ANA + 1 mg L-1 of BA), 20 petals showed response, in addition, the same variety in treatment 4 (1 mg L-1 ANA + 2 mg L-1 BA) presented 12 petals with adventitious buds. The ‘Gloriosa’ variety, in treatment 2 (1 mg L-1 of ANA + 1 mg L-1 of BA), 16 petals showed an organogenic response.

Number of buds per petal
The obtaining of shoots, by means of indirect organogenesis, began in the area of insertion of the petals with the receptacle between 60 and 65 days after sowing. In the ‘Heidi’ variety, shoot formation started at 65 days in treatment 2 (1 mg L-1 ANA + 1 mg L-1 BA), obtaining an average of 13.2/petal and 105 days in treatment 4 (1 mg L-1 ANA + 2 mg L-1 BA) generating an average of 1.63/petal (Figure 3). Therefore, there is a significant difference between both treatments. In the case of the ‘Gloriosa’ variety, sprouting was observed at 60 days only in the treatment of 1 mg L-1 ANA + 1 mg L-1 BA with an average of 8.40/petal (Figure 3). According to the obtained results, the multiplication was achieved in treatment 2 with the concentration of 1:1 in the BA/ANA ratio.
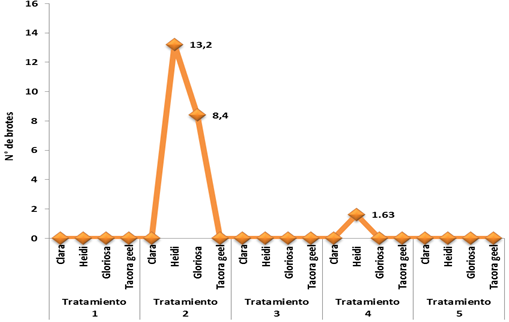
Figure 3 Combination effect of α-naphthalene-acetic acid (ANA) and benzyladenine (BA) on the number of buds/petal of the four varieties. The data are expressed as an average percentage of ten repetitions.
Theoretically, this ratio should be callus formation, but in this investigation, besides producing callus, shoot generation was induced, it can be assumed that the response is directly related to an endogenous concentration of the cytokinin in the explant. Usually a high concentration of cytokinins is required which, combined with an auxin, has more effective results in the case of the Begonia genus (Takayama and Misawa, 1982). Mendi (2009) considers that the combination 1 mg L-1 ANA + 1 mg L-1 BA produces a low regeneration and that it could be caused by an antagonistic effect of the ANA on BA since it considers that 1 mg L-1 of ANA is high for leaf and pedicel.
In contrast, Pierik and Tetteroo, (1987) induced sprouting in inflorescences (flower buds), not petals, using the concentration of 0.5 mg L-1 for both BA and ANA (1:1). Rosna et al. (2009) in petiole and Asmah Awal et al. (2013), using young inflorescences and peduncles as experimental material, promoted the formation of shoots with the treatment of 1 mg L-1 ANA + 1 mg L-1 BA with the addition to the MS medium of 40 mg L-1 of adenine. These results with sucrose confirm that the combination 1 mg L-1 ANA + 1 mg L-1 BA (1:1) is appropriate for the induction of the indirect organogenesis of adventitious shoots and consequently for the multiplication in petals of Begonia elatior.
It should be noted that during the development of the research an aspect considered convenient to mention was observed. It refers to the buds coming from petals of the varieties ‘Heidi’ and ‘Gloriosa’ planted in treatment 2, in which the formation of flower buds was obtained, which in some cases developed until the formation of the flower (Figure 4a, 4b). In this sense, it is necessary to point out that the petal is susceptible to generate floral buds through organogenesis. In this regard, Awal et al. (2013) obtained the formation of flowers by cultivating floral buds and peduncles Begonia elatior plants in MS medium supplemented ANA and BA in concentration with 1 mg L-1 ANA + 1 mg L-1 BA (1:1), but added adenine (40 mg L-1). Considers that it is possible that the concentration of sucrose may induce the process of floral bud formation, without defining the optimum concentration. Nguyen et al. (2006) determined that the process of formation of floral buds is due to the relationship sucrose and cytokinins which allows the induction of flowering in the Begonia system.

Figure 4 a) generation of bud floral bud of var. ‘Glorious’ in MS medium supplemented with ANA and BA (arrow); and b) development of the floral button of the var. ‘Heidi’ in vitro.
In general, sucrose is frequently used as a carbon source for in vitro studies with flowers. Takimoto (1960) stated that plants that produce small amounts of chlorophyll could have the ability to generate the formation of flower buds irrespective of light conditions and that exogenous sucrose can replace the requirement of high intensity of light. It is assumed that floral initiation in vitro may require a long incubation period of darkness and the presence of sucrose, in addition to the hormones. Jumin and Nito (1996) in Fortunella hindsii and Kachonpadungkitti et al. (2001) in Fagopyrum esculentum, concluded that sucrose has an effect on the induction of flower buds. Zhang et al. (2008) mentioned that, in Perilla frutescens, the presence of sucrose, from inflorescence cultures, induces the formation of floral buds. It is worth mentioning that this is the first time that it is mentioned that from petals of Begonia elatior one can obtain, on the one hand, seedlings and on the other, the possibility of inducing the formation of floral buds and in vitro flowering.
Root training
The rhizogenesis was induced only in the ‘Heidi’ variety (Figure 5) at 90 days after sowing, in the MS medium in treatment 2 (1 mg L-1 ANA + 1 mg L-1 BA). Seedlings of var. ‘Heidi’ developed roots of approximately, on average, 1.5 cm in length.

Conclusions
The main objective was fulfilled to know if the petals are able to de-differentiate and suffer a redifferentiation when obtaining the seedlings.
The organogenesis or obtaining adventitious shoots is feasible from callus derived from the petal in the two concentrations of BA and ANA used in the experiment (1 mg L-1 ANA + 1 mg L-1 BA and 1 mg L-1 ANA + 2 mg L-1 BA).
For the induction of callus, BA and NAA are required at a concentration of 1 mg L-1 ANA + 1 mg L-1 BA.
The cultivation of tissues through the cultivation of petals of Begonia elatior, can improve the ornamental propagation to achieve a low cost production.
The rooting of the seedlings is obtained in the multiplication medium with 1 mg L-1 ANA + 1 mg L-1 BA.
The petal color factor must be studied to know if it affects organogenesis since not all varieties produced seedlings. Apparently, organogenesis occurs in intense colors such as red and orange. The light colors (yellow and white) showed no sign of organogenesis.
Another factor that requires study is to determine if cytokinins and sucrose are or not induce the formation of floral buds in the in vitro culture of petals of Begonia elatior.
Literatura citada
Asmah, A.; Abdul, B. A. A.; Rosna, M. T. and Jamilah, S. Y. and Sadegh, M. 2013. Effect of adenine, sucrose and plant growth regulators on the indirect organogenesis and on in vitro flowering in Begonia x hiemalis Fotsch. AJCS. 7(5):691-698. [ Links ]
Bouman, H. and De Klerk, G. J. 2001. Measurement of the extent of somaclonal variation in Begonia plants regenerated under various conditions. Comparison of Three Assays, Theor. Appl. Genet. 102(1):111-117. [ Links ]
Bowes, B. G. and Curtis, E. W. 1991. Conservation of the British national Begonia collection by micropropagation. New Phytologist. 119(1):169-181. [ Links ]
Burritt, D. J. and Leung, D. W. M. 2003. Adventitious shoot regeneration from Begonia x Erythrophylla petiole sections is developmentally sensitive to light quality. Physiol. Plantarum. 118(2):289-296. [ Links ]
Chebet, D. K.; Okeno, J. A. and Mathenge, P. 2003. Biotechnological approaches to improve horticultural crop production. Acta Hortic. 625:473-477. [ Links ]
Datta, S. K. 2002. In vitro petal culture and callus formation in Rosa species. Indian J. Agric. Sci. 72(5):271-276. [ Links ]
Espino, F. J.; Linacero, R.; Rueda, J. and Vázquez, A. M. 2004. Shoot regeneration in four Begonia genotypes. Biol. Plant. 48(1):101-104. [ Links ]
Jumin, H. B. and Nito, N. 1996. In vitro flowering of Fortunella hindsii (Champ.). Plant Cell Rep. 15:484-488. [ Links ]
Kabirnataj, S.; Ghasemi, Y.; Nematzadeh, G.; Asgharzadeh, R.; Shahin, K. B. and Yazdani, M. 2012. Effect of explant type and growth regulators on in vitro micropropagation of Begonia rex. Inter. Res. J. Appl. Basic Sci. 3(4):896-901. [ Links ]
KachonpadungkittI, Y.; Romchatngoen, S.; Hasegawa, K. and Hisajima, S. 2001. Efficient flower induction from cultured buckwheat (Fagopyrum esculentum L.) node segments in vitro. Plant Growth Regul. 35:37-45. [ Links ]
Kishimoto, S.; Aida, R. and Shibata, M. 2002. Agrobacterium tumefaciensmediated transformation of elatior Begonia (Begonia x Hiemalis Fotsch). Plant Sci. 162(5): 697-703. [ Links ]
Mendi, Y. Y.; Curuk, P.; Kocaman, E.; Unek, C.; Eldogan, S.; Gencel, G. and Cetiner, S. 2009. Regeneration of begonia plantlets by direct organogenesis. Afr. J. Biotechnol. 8(9):1860-1863. [ Links ]
Murashige, T. and Skoog, F. 1962. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Plant Physiol. 15:473-497. [ Links ]
Nakano, M.; Niimi, Y.; Kobayashi, D. and Watanabe, A. 1999. Adventitious shoot regeneration and micropropagation of hybrid tuberous Begonia (Begonia x Tuberhybrida Voss). Scı. Hortic. 79(3-4):245-251. [ Links ]
Nguyen, H. V.; Phan, H. A. and Duong, T. N. 2006. The role of sucrose and different cytokinins in the in vitro floral morphogenesis of rose (Hybrid Tea) cv. ‘First Prize’. Plant Cell, Tissue and Organ Culture. 87(3):315-320. [ Links ]
Pierik, R. L. M. and Tetteroo, F. A. A. 1987. Vegetative propagation of Begonia venosa Skan in vitro from inflorescence explants. Plant Cell, Tissue and Organ Culture . 10:135-142. [ Links ]
Rosna, M. T.; Nor, A. H. and Asmah, A. 2009. In vitro flowering of selected ornamental plants. Acta Hortic . 881(881):141-146. [ Links ]
Rout, G. R.; Mohapatra, A. and S. Mohan, J. 2006. A Tissue culture of ornamental pot plant: A critical review on present scenario and future prospects. Biotechnol. Adv. 24:531-560. [ Links ]
Shimada, Y.; Mori, G.; Katahara, Y. and Oda, M. 2006. Formation of adventitious buds on leaf pieces cutting of Begonia Tuberhybrida Group. J. Jpn. Soc. Hortic. Sci. 75(4):318-322. [ Links ]
Shimada, Y.; Mori, G.; Oda, M. and Ishida, G. 2007. Effects of BA and leaf piece orientation on adventitious bud formation in leaf cutting of Begonia Tuberhybrida Group. J. Jpn. Soc. Hortic. Sci. 76(2):157-162. [ Links ]
Simmonds, J. and Werry, T. 1987. Liquid shake cultures for improved micropropagation of Begonia × hiemalis. Hortic Sci. 22:122-124. [ Links ]
Takayama, S. and Misawa, M. 1982. Factors affecting differentiation and growth in vitro and a mass-propagation scheme for Begonia x hiemalis. Sci. Hortic. 16:65-75. [ Links ]
Takimoto, A. 1960. Effect of sucrose on flower initiation of Pharbitis nil in aseptic culture. Plant Cell Physiol. 1:241-24. [ Links ]
Zhang, Y. W.; Yang, C. F.; Gituru, W. R. and Guo, Y. H. 2008. Within-season adjustment of sex expression in females and hermaphrodites of the clonal gynodioecious herb Glechoma longituba (Lamiaceae). Ecol Res. 23:873-881. [ Links ]
Received: June 2018; Accepted: August 2018
 Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
