











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
El tomate (Solanum lycopersicum L.) es uno de los cultivos de hortalizas más importantes y ampliamente cultivadas en el mundo y el ácaro de las dos manchas, Tetranychus urticae Koch (Acari: Tetranychidae) es una de las plagas importantes de este cultivo, (Saeidi y Mallik, 2012). El daño provocado por este ácaro consiste en la remoción del contenido celular de la planta, causando pequeños puntos pálidos o cicatrices donde las células epidérmicas han sido destruidas ocasionando clorosis, defoliación e incluso muerte de la planta, lo cual contribuye a la reducción del rendimiento (Tomczyk y Kropczynska, 1985).
Una de las herramientas más utilizadas para el manejo de este acaro es el control químico (Takematsu et al., 1994); sin embargo, el mayor problema que enfrenta este tipo de control es su rápida habilidad para desarrollar resistencia (Stumpf y Nauen, 2001, 2002). A nivel mundial se han reportado más de 200 casos de resistencia de T. urticae (Konanz y Nauen, 2004) por lo mismo, el uso de variedades de plantas resistentes es una herramienta que puede ayudar en el control de esta plaga (Flexner et al. 1995) y puede ser considerado como parte del manejo integrado de esta (Zehnder et al., 2007). Este tipo de control se da como respuesta a mecanismos tales como: antixenosis, tolerancia y antibiosis o una combinación de estos (Granados y Paliwal, 2001).
La antixenosis se presenta cuando las características físicas y químicas de la planta sirven como una barrera para alguna de las actividades del artrópodo, se observa tolerancia cuando una planta puede mantener elevadas poblaciones de una plaga sin que se disminuya su producción y por último, ocurre antibiosis cuando la planta afecta de alguna manera en forma negativa la fisiología del insecto (Álvarez et al., 2001).
Los cultivos resistentes a los artrópodos proporcionan importantes beneficios ecológicos y económicos para la agricultura global, las interacciones incompatibles entre plantas resistentes y artrópodos de plagas avirulentas están mediadas por proteínas vegetales inducidas constitutivamente y producidas por artrópodos y aleloquímicos de defensa sintetizados por productos de genes de resistencia, (Smith y Clement, 2012). Además, el uso de variedades resistentes disminuye parcialmente la necesidad de aplicaciones de plaguicidas y por tanto mejora la población de enemigos naturales (Desneux et al., 2007).
Sin duda la genómica de plantas y artrópodos ofrece muchas oportunidades para desarrollar más eficientemente las plantas resistentes a los artrópodos, aunque para la integración de cultivos resistentes en los programas de MIP se requiere además de otros estudios de carácter interdisciplinario. Debido a lo anterior, existe la necesidad de conocer en primera instancia el comportamiento de T. urticae en determinadas variedades de tomate, posteriormente conocer las características que le confieren resistencia contra esta plaga, por último, crear variedades con características de tolerancia que les permitan ser competitivas.
Por tal motivo, el objetivo de esta investigación fue dar inicio al primer punto; es decir, evaluar los niveles de tolerancia, repelencia y mortalidad de 10 variedades de tomate a T. urticae y el grado en que las variedades ‘Toro’, ‘Saladette’ ‘Cereza’ y ‘Tsan-101’ propician el desarrollo de esta plaga. Lo anterior bajo la hipótesis de que el comportamiento poblacional de T. urticae es diferente en las variedades en estudio, algunas de ellas con factores que impiden o limitan su desarrollo.
Materiales y métodos
El experimento se llevó a cabo en una cámara bioclimática del departamento de Parasitología Agrícola de la Universidad Autónoma Agraria Antonio Narro (UAAAN). Se utilizó una colonia de laboratorio de T. urticae previamente establecida bajo condiciones controladas de 25 ±2 ºC, humedad relativa de 60-70% y un fotoperiodo de 12 h sobre plántulas de frijol (Phaseolus vulgaris L.). Las plántulas de tomate se cultivaron en semilleros con 10 cm de profundidad, el sustrato utilizado fue una mezcla de tierra de monte, agrolita y turba (Peat Moss) en proporción 1:1:1.
Cuando las plántulas presentaron al menos cinco hojas verdaderas se trasladaron a una cama de invernadero de 60 cm x 9 m. con una distancia entre plantas de 20 cm. y entre variedades de 50 cm, una temperatura de 28 ±4 ºC y una HR de 60 ±15%. Se fertilizaron cada 20 días mediante el producto comercial Yaramila Complex® el cual contiene 12% de nitrógeno, 11% de fósforo y 8% de potasio y riego por goteo durante todo el desarrollo del experimento.
Para el manejo del material en laboratorio se utilizó la técnica de Abou-setta y Childers (1987) conocida como hoja arena, que consistió en la transferencia de ácaros mediante un pincel de pelo de camello 000 a hojas de tomate con al menos cinco hojas verdaderas donde se extrajeron 90 discos de las hojas con la ayuda de un sacabocados de 13 mm de diámetro, se colocaron por el envés en cajas de Petri provistas de algodón saturado con agua destilada y se colocaron 10 hembras adultas de T. urticae, por disco con 90 repeticiones por variedad.
Se registró el comportamiento de la arañita de dos manchas mediante los factores de supervivencia, mortalidad y repelencia, esta última en base al comportamiento de dejar los discos de la hoja durante 4 días. Para determinar los parámetros demográficos de T. urticae, se colocaron 25 hembras durante 24 h para que ovipositaran, posteriormente se removieron las hembras dejando solamente los huevos los cuales eclosionaron y la colonia de nuevos individuos continuaron su desarrollo hasta que alcanzaron el estado adulto.
Posteriormente se seleccionaron 100 hembras de un día de edad recién apareadas y se colocaron individualmente en los discos de hojas de tomate de las variedades en estudio. Estas hembras apareadas se mantuvieron bajo las mismas condiciones ambientales que la colonia madre de tal forma que cada unidad experimental consistió de una hembra por disco. Los huevos depositados por estas hembras se mantuvieron en el mismo disco hasta la aparición de las larvas las cuales fueron colocadas luego individualmente en otros discos foliares para que continuaran su desarrollo. A partir de este punto, se registró la supervivencia diaria y descendencia de las hembras adultas hasta que murió la última hembra. Se realizaron cálculos demográficos basados en el modelo de Birch (1948) y se utilizó el método de Jack nife para estimar las desviaciones estándar con un intervalo de confianza de 95%.
Los resultados registrados para la supervivencia, mortalidad y repelencia fueron sometidos a análisis de varianza (Anova) con un diseño completamente al azar con 90 repeticiones, cuando el Anova indico la existencia de diferencias significativas entre los tratamientos, se aplicó la prueba de Tukey (p≤ 0.05) para la separación de medias. Para cada uno de los análisis se utilizó el programa RStudio.
Resultados y discusión
En el Cuadro 1 se observan los resultados de las variables supervivencia, mortalidad y repelencia de T. urticae sobre las 10 variedades en estudio. Como se puede observar la variedad Cereza presentó el mayor porcentaje de supervivencia con 76.78% seguido por la variedad La-1959 y Paloma (70.92 y 70.03%) durante 4 días de observación. Mientras que las variedades que resultaron con mayor mortalidad fueron Toro, Pegaso y Monte Carlos con 18.28, 16.14 y 12.86% de mortalidad respectivamente. Como se observa la variedad Pegaso y Monte Carlos presentaron 11.70 y 29.64% menos mortalidad que la variedad Toro; lo anterior, los coloca como las variedades menos adecuadas para el desarrollo poblacional de esta especie. La variedad toro resultó ser 68.24% más resistente que la variedad Cereza.
Cuadro 1 supervivencia, repelencia y mortalidad de Tetranychus urticae en 10 variedades de tomate. Las medias fueron tomadas después de cuatro días de infestación.
| Variedades | Mortalidad | Repelencia | Supervivencia | |||
|---|---|---|---|---|---|---|
| Media ± D S | Dif* | Media ± D S | Dif | Media ± D S | Dif | |
| Toro | 18.28 ±20.26 a | 0 | 28.83 ±22.72 ab | -7.82 | 52.94 ±33.19 f | -31 |
| Pegaso | 16.14 ±17.94 ab | -11.7 | 24.25 ±20.63 bc | -22.47 | 59.58 ±29.26 de | -22.4 |
| Monte Carlos | 12.86 ±18.45 bc | -29.64 | 27.06 ±25.12 abc | -13.5 | 59.78 ±34.31 de | -22.1 |
| Tsan-101-su-9 | 12.31 ±17.68 cd | -32.68 | 31.28 ±24.9 a | 0 | 56.47 ±29.76 ef | -26.5 |
| Paloma | 11.06 ±14.89 cd | -39.51 | 18.75 ±17.82 de | -40.05 | 70.31 ±23.65 b | -8.42 |
| La-1959 | 10.36 ±16.51 cd | -43.31 | 18.19 ±16.72 e | -41.83 | 70.92 ±23.72 ab | -7.63 |
| Saladette | 9.25 ±13.36 cde | -49.4 | 23.64 ±20.2 cd | -24.42 | 67.08 ±25.44 bc | -12.7 |
| Tsan-103-su-n | 8.75 ±14.51 de | -52.13 | 28.31 ±24.26 abc | -9.5 | 62.92 ±28.45 cd | -18 |
| Bola | 6.53 ±10.31 e | -64.29 | 23.86 ±21.25 c | 69.64 ±24.53 b | -9.3 | |
| Cereza | 5.81 ±9.2 e | -68.24 | 17.42 ±16.6 e | -44.32 | 76.78 ±19.4 a | 0 |
*= diferencia en términos de porcentaje en relación a la mayor cifra registrada en cada columna.
Es importante conocer que factor o factores de la planta inciden en el comportamiento poblacional del acaro. La prueba de Tukey señala que la respuesta en relación al comportamiento poblacional de las variedades en estudio fue muy diferente en las 10 variedades en estudio colocando la variedad Cereza como la más susceptible con menor mortalidad, menor repelencia y mayor supervivencia, por su parte la variedad Toro registró una mayor mortalidad menor supervivencia y la segunda mayor en repelencia. Aunque recientemente se ha publicado el genoma completo de T. urticae el cual ha mostrado una proliferación de familias de genes asociados
Con la digestión y desintoxicación de compuestos secundarios de plantas que respalda su comportamiento polífago, existen un grupo de moléculas que repelen o disminuyen el desarrollo de esta especie (Santamaría et al., 2012). Son múltiples los factores que inciden para que una planta resulte adversa para el desarrollo de plagas, al respecto Smith (2005) menciona que la resistencia de las plantas a los artrópodos es la suma de los componentes constitutivos, cualidades genéticamente heredadas que resultan en una planta menos dañada que una planta susceptible que carece de estas cualidades.
Un estudio realizado por Alba et al. (2009) encontraron diferencias altamente significativas al desarrollo de T. urticae entre las más susceptibles y las más resistente de S. pimpinellifolium (TO-937), debido a un elevado contenido de acilsacarosa y una alta densidad de tricomas tipo IV que aumentaron la mortalidad y repelencia y también se registró una reducción de la oviposición de T. urticae. Kawazu et al. (2012) compararon los perfiles de expresión de los genes inducibles para ácido jasmónico (Pin2 y LapA1) y ácido salicílico (PRb-1b y GluB) en plantas de tomate en respuesta a la infestación de T. urticae, se activaron los genes para ácido jasmónico LapA1 y adicionalmente los genes para ácido salicílico (PRb-1b y GluB), lo cual representa una señal de que estas moléculas también intervienen de alguna forma en el proceso de resistencia en este cultivo.
Además, para el caso del tomate algunos estudios indican que las plantas que crecen en condiciones de invernadero a baja intensidad de luz durante días largos reducen sus defensas aleloquímicas y son más susceptibles a T. urticae (Jansen y Stamp, 1997). Otros factores como la inducción de metabolitos secundarios, la morfología de la superficie del folículo foliar y la presencia de enemigos naturales también juegan un papel importante en la aceptación de las plantas (Van den Boom et al., 2003).
Supervivencia y fecundidad
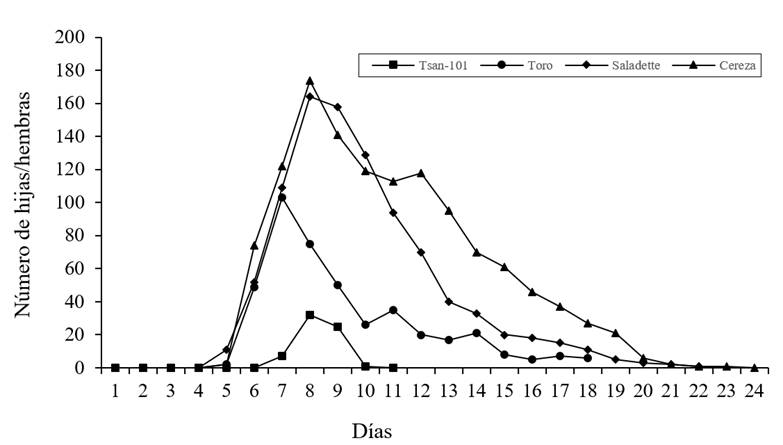
En relación con la tasa de supervivencia (Figura 1) se muestran diferencias significativas (prueba de Test Log-Rank p≤ 0.05,) donde la variedad Tsan-101 y Toro, mostraron mayor proporción de mortalidad en comparación a las demás variedades, el rango de hembras vivas en la variedad se redujo considerablemente durante los primeros seis días mientras que las demás variedades Cereza y Saladette mostraron una supervivencia similar. La tasa de fecundidad por edad especifica de T. urticae (Figura 2) e Tsan-101 fue menor que las demás variedades la variedad Cereza produjo un considerable mayor número de hembras hijas.

Figura 1 Curvas de supervivencia de Tetranychus urticae Koch. en cuatro variedades de tomate (S. lycopersicum) ‘Tsan-101’, ‘Toro’, ‘Saladette’ y ‘Cereza’.
Parámetros poblacionales
En relación a la tasa bruta de reproducción (GRR); es decir, el número de hijas hembras de T. urticae producidas por madre a través de todas las edades, el valor más alto (58.87) fue registrado en ‘Tsan-101’ seguido por ‘Toro’ (47.71) ‘Cereza’ (46.24) y ‘Saladette’ (32.96) (Cuadro 2). Los valores de GRR obtenidos en esta investigación indican claras diferencias en la capacidad reproductiva de T. urticae en las cuatro variedades de tomate; no obstante, estos valores fueron inferiores a lo reportado por Landeros et al. (2013) quienes registraron un GRR de 78.33 sobre rosales y Sáenz de Cabezón et al. (2006) reportaron un GRR de 85.88 para T. urticae en discos de hoja de frijol.
Cuadro 2 Parámetros poblacionales de Tetranychus urticae en discos de hojas de cuatro variedades de tomate ‘T-SAN101’, ‘Toro’, ‘Saladette’ y ‘Cereza’.
| Parámetros poblacionales | TSAN-101 | Toro | Saladette | Cereza |
|---|---|---|---|---|
| GRR | 58.87 | 47.71 | 32.96 | 46.24 |
| Ro | 2.32 | 6.05 | 11 | 14.14 |
| Tc | 6.307 | 8.202 | 8.914 | 9.898 |
| rc | 0.133 | 0.219 | 0.268 | 0.267 |
| rm | 0.1341 | 0.2428 | 0.3012 | 0.3123 |
| T2 | 5.16 | 2.85 | 2.3 | 2.21 |
| λ | 1.14 | 1.27 | 1.35 | 1.36 |
| TG | 6.28 | 7.41 | 7.96 | 8.48 |
Con respecto a la tasa de la reproducción neta (Ro) o (el promedio del número de hijas que una hembra produce durante su vida) la variedad ‘Cereza’ registró el mayor número de hijas de T. urticae o hembra en una generación (14.14) seguido de ‘Saladette’ (11), ’Toro’ (6.05) y ‘Tsan-101’ (2.32). Estos valores representar una considerable reducción de Ro de T. urticae en las diferentes variedades (Cuadro 2) así en la variedad ‘Tsan-101’ y ‘Toro’ se registró un desarrollo poblacional de T. urticae más lento en comparación a ‘Saladette’ y ‘Cereza’ donde esta última se mostró como la más susceptible. Estos datos son inferiores a lo reportado por Amala et al. (2016) que registraron un valor de Ro de 125.43 en discos de hojas de vid en la variedad Thompson Seedless.
En relación a la tasa intrínseca de crecimiento (rm); es decir, la tasa de incremento poblacional en ausencia de una fuerza denso-dependiente se muestra que la variedad ‘Cereza’ (0.3123) registró (rm) mayor de T. urticae que las demás y por lo mismo se consideró más susceptible a esta plaga seguida por ‘Saladette’ (0.3012), ‘Toro’ (0.2428) y ‘Tsan-101’ (0.1341). Estos resultados son similares a los valores que oscilan entre 0.254 y 0.313 en cinco cultivares de pepino (Cucumis sativus L.) (Kheradpir et al., 2007). Y mayores a los registrados por Hoque et al. (2008) quienes registran en rm de 0.1873 en discos de hojas de frijol (Lablab purpureus L.).
En cuanto al tiempo medio entre generaciones (TG), el valor más alto fue de 8.48 días, con un aumento diario de la población de 1.36 fue registrado para ‘Cereza’ seguido por los tiempos progresivamente más cortos en ‘Saladette’, ‘Toro’ y ‘Tsan-101’ (Cuadro 2), dichos resultados implican un mayor potencial de daño en la variedad ‘Cereza’ en comparación con las otras 3 variedades y son resultados inferiores a los reportados por Riahi et al. (2011) sobre variedades de Melocotón donde registraron un TG de 16.9 días y un factor de crecimiento diario de 1.07. Por último, el tiempo de duplicación (T2) de la población de T. urticae en ‛Tsan-101’, ‘Toro’, ‘Saladette’ y ‘Cereza’ fueron de 5.16, 2.85, 2.3 y 2.21 días, respectivamente, donde Tsan-101 presentó mayor resistencia para el desarrollo de la población seguido por ‘Toro’, ‘Saladette’ y ‘Cereza’.
Conclusión
De las variedades de tomate en estudio, Toro y Tsan 101 presentaron características de resistencia a T. urticae que cambiaron el comportamiento y respuesta hacia las plantas; lo anterior, coloca a estas dos variedades como adecuadas para el desarrollo de este cultivo en áreas con alta incidencia de esta plaga, además de que pueden ser utilizadas en programas de mejoramiento una vez que se conozcan cuáles son los factores que inciden en la defensa de ellas.

