










Services on Demand
Journal
Article
 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista mexicana de ciencias agrícolas
Print version ISSN 2007-0934
Rev. Mex. Cienc. Agríc vol.9 n.5 Texcoco Jun./Aug. 2018
https://doi.org/10.29312/remexca.v9i5.1505
Articles
Population behavior of Tetranychus urticae Koch. (Acari: Tetranychidae) in tomato varieties
1Department of Agricultural Parasitology-Autonomous Agrarian University Antonio Narro. Calzada Antonio Narro 1923, Saltillo, Coahuila, Mexico. CP. 25315.
Ten tomato varieties were examined for resistance to Tetranychus urticae by bioassays on leaf discs. A colony of T. urticae from different ornamental crops of Saltillo, Coahuila, Mexico, was established in bean seedlings (Phaseolus vulgaris L.) in a Biotronette chamber. At 25 ±2 °C, 60-70 HR and 12:12 h L: D. According to the design of the experiment, the females were placed in leaf discs of the 10 tomato varieties under study in a proportion of 10 disc with 90 repetitions, the behavior was evaluated based on survival, repellency and mortality. Subsequently, 100 freshly mated and fertilized one-day-old females were transferred to discs of tomato leaves (Solanum lycopersicum L.) of 1.3 cm in diameter of cultivars ‛Tsan-101’, ‛Toro’, ‛Saladette’ and ‛Cereza’ in such a way that each experimental unit included 1 female per disk. The demographic parameters in this experiment showed a lower growth potential of this pest in the cultivars ‛Tsan-101’ and ‛Toro’ than in ‛Saladette’ and ‛Cereza’.
Keywords: Tetranychus urticae; population parameters; resistance; tomato
Se examinaron diez variedades de tomate para detectar resistencia a Tetranychus urticae mediante bioensayos en discos de hojas. Se estableció una colonia de T. urticae de diferentes cultivos ornamentales de Saltillo, Coahuila, México, en plántulas de frijol (Phaseolus vulgaris L.) en una cámara Biotronette. A 25 ±2 °C, 60-70 HR y 12:12 h L: D. De acuerdo con el diseño del experimento, las hembras fueron colocadas en discos de hojas de las 10 variedades de tomate bajo estudio en una proporción de 10 por disco con 90 repeticiones, el comportamiento se evaluó en función de la supervivencia, repelencia y mortalidad. Posteriormente, se transfirieron 100 hembras recién apareadas y fecundadas de un día de edad a discos de hojas de tomate (Solanum lycopersicum L.) de 1.3 cm de diámetro de cultivares ‘Tsan-101’, ‘Toro’, ‘Saladette’ y ‘Cereza’ de tal manera que cada unidad experimental incluía 1 hembra por disco. Los parámetros demográficos en este experimento mostraron un menor potencial de crecimiento de esta plaga en los cultivares ‘Tsan-101’ y ‘Toro’ que en ‘Saladette’ y ‘Cereza’.
Palabras clave: Tetranychus urticae; parámetros poblacionales; resistencia; tomate
Introduction
The tomato (Solanum lycopersicum L.) is one of the most important and widely cultivated vegetable crops in the world and the two-spotted mite, Tetranychus urticae Koch (Acari: Tetranychidae) is one of the important pests of this crop (Saeidi and Mallik, 2012). The damage caused by this mite consists of the removal of the cellular content of the plant, causing small pale spots or scars where the epidermal cells have been destroyed causing chlorosis, defoliation and even death of the plant, which contributes to the reduction of the yield (Tomczyk and Kropczynska, 1985).
One of the most used tools for managing this mite is chemical control Takematsu et al. (1994); however, the biggest problem facing this type of control is its rapid ability to develop resistance (Stumpf and Nauen, 2001, 2002). Worldwide, more than 200 cases of T. urticae resistance have been reported (Konanz and Nauen, 2004) therefore, the use of resistant plant varieties is a tool that can help in the control of this pest (Flexner et al., 1995) and can be considered as part of integrated management (Zehnder et al., 2007). This type of control is given in response to mechanisms such as: antixenosis, tolerance and antibiosis or a combination of these (Granados and Paliwal, 2001).
The antixenosis occurs when the physical and chemical characteristics of the plant serve as a barrier to some of the activities of the arthropod, tolerance is observed when a plant can maintain high populations of a pest without decreasing its production and finally, antibiosis occurs when the plant affects in a negative way the physiology of the insect (Álvarez et al., 2001).
Arthropod resistant crops provide important ecological and economic benefits for global agriculture, the incompatible interactions between resistant plants and avirulent pest arthropods are mediated by plant proteins constitutively induced and produced by defense arthropods and allelochemicals synthesized by resistance gene products, (Smith and Clement, 2012). In addition, the use of resistant varieties partially decreases the need for pesticide applications and therefore improves the population of natural enemies (Desneux et al., 2007).
Undoubtedly, the genomics of plants and arthropods offer many opportunities to develop more resistant plants resistant to arthropods, although for the integration of resistant crops in MIP programs, other interdisciplinary studies are required. Due to the above, there is a need to know in the first instance the behavior of T. urticae in certain varieties of tomato, later know the characteristics that give resistance against this pest, finally, create varieties with characteristics of tolerance that allow them to be competitive.
For this reason, the objective of this investigation was to start the first point; that is, to evaluate the levels of tolerance, repellency and mortality of 10 varieties of tomato to T. urticae and the degree to which the varieties ‛Toro’, ‛Saladette’ ‛Cereza’ and ‛Tsan-101’ promote the development of this pest. The above under the hypothesis that the population behavior of T. urticae is different in the varieties under study, some of them with factors that impede or limit their development.
Materials and methods
The experiment was carried out in a bioclimatic chamber of the Department of Agricultural Parasitology of the Autonomous Agrarian University Antonio Narro (UAAAN). A previously established T. urticae laboratory colony was used under controlled conditions of 25 ±2 °C, relative humidity of 60-70% and a 12-hour photoperiod on bean seedlings (Phaseolus vulgaris L.). The tomato seedlings were grown in seedbeds with a depth of 10 cm, the substrate used was a mixture of forest soil, agrolita and peat (Peat moss) in a ratio of 1:1:1.
When the seedlings presented at least five true leaves they were moved to a greenhouse bed of 60 cm x 9 m. with a distance between plants of 20 cm. and between varieties of 50 cm, a temperature of 28 ±4 ºC and an HR of 60 ±15%. They were fertilized every 20 days by the commercial product Yaramila Complex® which contains 12% nitrogen, 11% phosphorus and 8% potassium and drip irrigation throughout the development of the experiment.
The Abou-Setta and Childers technique (1987), known as the sand sheet, was used to handle the material in the laboratory, which consisted in the transfer of mites using a camel hair brush 000 to tomato leaves with at least five true leaves where 90 discs were extracted from the leaves with the help of a 13 mm diameter punch, they were placed on the back in Petri dishes provided with cotton saturated with distilled water and 10 adult females of T. urticae were placed, per disc with 90 repetitions per variety.
The behavior of the spider of two spots was recorded by the factors of survival, mortality and repellency, the latter based on the behavior of leaving the discs of the leaf for 4 days. To determine the demographic parameters of T. urticae, 25 females were placed for 24 h to oviposit, then the females were removed leaving only the eggs which hatched and the colony of new individuals continued their development until they reached the adult stage.
Subsequently, 100 freshly matched one-day-old females were selected and placed individually on the discs of tomato leaves of the varieties under study. These mated females were kept under the same environmental conditions as the mother colony in such a way that each experimental unit consisted of one female per disk. The eggs deposited by these females were kept in the same disc until the appearance of the larvae which were then placed individually in other leaf discs to continue their development. From this point, the daily survival and offspring of the adult females was recorded until the last female died. Demographic calculations were performed based on the Birch model (1948) and the Jack nife method was used to estimate standard deviations with a confidence interval of 95%.
The results recorded for survival, mortality and repellency were subjected to analysis of variance (Anova) with a completely random design with 90 repetitions, when the Anova indicated the existence of significant differences between the treatments, the Tukey test was applied (p≤ 0.05) for the separation of means. The RStudio program was used for each of the analyzes.
Results and discussion
In the Table 1 shows the results of the variables survival, mortality and repellency of T. urticae on the 10 varieties under study. As you can see the variety Charry presented the highest percentage of survival with 76.78% followed by the variety La-1959 and Paloma (70.92 and 70.03%) during 4 days of observation. While the varieties with the highest mortality were Toro, Pegaso and Monte Carlos with 18.28, 16.14 and 12.86% mortality respectively. As it is observed the variety Pegaso and Monte Carlos presented 11.7 and 29.64% less mortality than the Toro variety; the above, places them as the less suitable varieties for the population development of this species. The Toro variety turned out to be 68.24% more resistant than the variety Cereza.
Table 1 Survival, repellency and mortality of Tetranychus urticae in 10 tomato varieties. The stockings were taken after four days of infestation.
| Varieties | Mortality | Repellency | Survival | |||
|---|---|---|---|---|---|---|
| Mean ± DS | Dif* | Mean ± DS | Dif | Mean ± DS | Dif | |
| Toro | 18.28 ±20.26 a | 0 | 28.83 ±22.72 ab | -7.82 | 52.94 ±33.19 f | -31 |
| Pegaso | 16.14 ±17.94 ab | -11.7 | 24.25 ±20.63 bc | -22.47 | 59.58 ±29.26 de | -22.4 |
| Monte Carlos | 12.86 ±18.45 bc | -29.64 | 27.06 ±25.12 abc | -13.5 | 59.78 ±34.31 de | -22.1 |
| Tsan-101-su-9 | 12.31 ±17.68 cd | -32.68 | 31.28 ±24.9 a | 0 | 56.47 ±29.76 ef | -26.5 |
| Paloma | 11.06 ±14.89 cd | -39.51 | 18.75 ±17.82 de | -40.05 | 70.31 ±23.65 b | -8.42 |
| La-1959 | 10.36 ±16.51 cd | -43.31 | 18.19 ±16.72 e | -41.83 | 70.92 ±23.72 ab | -7.63 |
| Saladette | 9.25 ±13.36 cde | -49.4 | 23.64 ±20.2 cd | -24.42 | 67.08 ±25.44 bc | -12.7 |
| Tsan-103-su-n | 8.75 ±14.51 de | -52.13 | 28.31 ±24.26 abc | -9.5 | 62.92 ±28.45 cd | -18 |
| Bola | 6.53 ±10.31 e | -64.29 | 23.86 ±21.25 c | 69.64 ±24.53 b | -9.3 | |
| Cereza | 5.81 ±9.2 e | -68.24 | 17.42 ±16.6 e | -44.32 | 76.78 ±19.4 a | 0 |
*= difference in terms of percentage in relation to the highest figure registered in each column.
It is important to know what factor or factors of the plant affect the population behavior of the mite. The Tukey test indicates that the response in relation to the population behavior of the varieties under study was very different in the 10 varieties under study, placing the Cereza variety as the most susceptible with lower mortality, lower repellency and greater survival, on the other hand the variety Toro recorded a higher mortality, lower survival and the second greater in repellency. Although the complete genome of T. urticae has recently been published, which has shown a proliferation of associated gene families
With the digestion and detoxification of plant secondary compounds that supports its polyphagous behavior, there is a group of molecules that repel or diminish the development of this species (Santamaria et al., 2012). There are multiple factors that affect a plant to be adverse for the development of pests, in this regard (Smith, 2005) mentions that the resistance of plants to arthropods is the sum of the constituent components, genetically inherited qualities that result in a plant less damaged than a susceptible plant that lacks these qualities.
A study conducted by Alba et al. (2009) found highly significant differences in the development of T. urticae between the most susceptible and the most resistant of S. pimpinellifolium (TO-937), due to a high content of acylsacarosa and a high density of type IV trichomes that increased mortality and repellency and a reduction in the oviposition of T. urticae was also recorded. Kawazu et al. (2012) compared the expression profiles of the inducible genes for jasmonic acid (Pin2 and LapA1) and salicylic acid (PRb-1b and GluB) in tomato plants in response to the T. urticae infestation, genes for jasmonic acid were activated LapA1 and additionally the genes for salicylic acid (PRb-1b and GluB), which represents a signal that these molecules also intervene in some way in the resistance process in this culture.
In addition, in the case of tomato some studies indicate that plants that grow under greenhouse conditions at low light intensity for long days reduce their allelochemical defenses and are more susceptible to T. urticae (Jansen and Stamp, 1997). Other factors such as the induction of secondary metabolites, the morphology of the leaf follicle surface and the presence of natural enemies also play an important role in the acceptance of plants (Van den Boom et al., 2003).
Survival and fertility
In relation to the survival rate (Figure 1) significant differences are shown (test of Test Log-Rank p≤ 0.05,) where the variety Tsan-101 and Toro, showed a greater proportion of mortality compared to the other varieties, the range of live females in the variety was considerably reduced during the first six days while the rest Cereza and Saladette varieties showed a similar survival. The specific fertility rate by age of T. urticae (Figure 2) and Tsan-101 was lower than the other varieties. The variety Cereza produced a considerable number of female daughters.
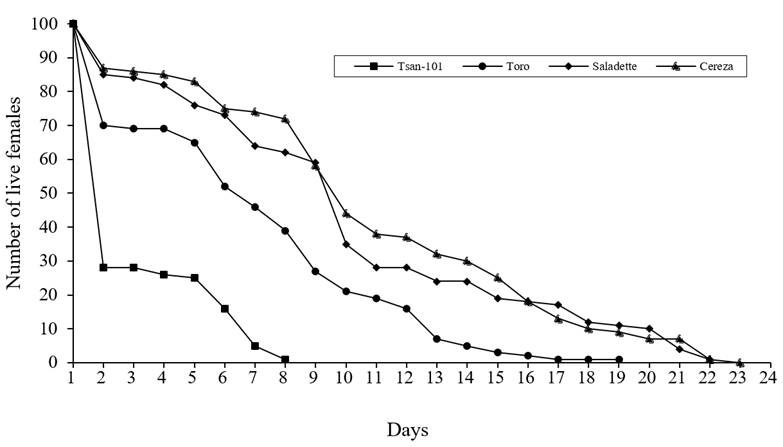
Figure 1 Survival curves of Tetranychus urticae Koch. in four varieties of tomato (S. lycopersicum) ‛Tsan-101’, ‛Toro’, ‛Saladette’ and ‛Cereza’.

Population parameters
In relation to the crude reproduction rate (GRR); that is, the number of female daughters of T. urticae produced by mother through all ages, the highest value (58.87) was recorded in ‛Tsan-101’ followed by ‛Toro’ (47.71) ‛Cereza’ (46.24) and ‛Saladette’ (32.96) (Table 2). The GRR values obtained in this investigation indicate clear differences in the reproductive capacity of T. urticae in the four tomato varieties, although these values were lower than that reported by Landeros et al. (2013) who registered a GRR of 78.33 on rose bushes and Saenz de Cabezon et al. (2006) reported a GRR of 85.88 for T. urticae on bean leaf discs.
Table 2 Population parameters of Tetranychus urticae in leaf discs of four tomato varieties ‛T-SAN101’, ‛Toro’, ‛Saladette’ and ‛Cereza’.
| Population parameters | TSAN-101 | Toro | Saladette | Cereza |
|---|---|---|---|---|
| GRR | 58.87 | 47.71 | 32.96 | 46.24 |
| Ro | 2.32 | 6.05 | 11 | 14.14 |
| Tc | 6.307 | 8.202 | 8.914 | 9.898 |
| rc | 0.133 | 0.219 | 0.268 | 0.267 |
| rm | 0.1341 | 0.2428 | 0.3012 | 0.3123 |
| T2 | 5.16 | 2.85 | 2.30 | 2.21 |
| λ | 1.14 | 1.27 | 1.35 | 1.36 |
| TG | 6.28 | 7.41 | 7.96 | 8.48 |
With respect to the rate of net reproduction (Ro) or (the average number of daughters that a female produces during her life) the ‛Cereza’ variety recorded the highest number of daughters of T. urticae or female in a generation (14.14) followed by ‛Saladette’ (11), ‛Toro’ (6.05) and ‛Tsan-101’ (2.32) These values represent a considerable reduction of Ro of T. urticae in the different varieties (Table 2) as well as in the variety ‛Tsan-101’ and ‛Toro’ recorded a population development of T. urticae slower compared to ‛Saladette’ and ‛Cereza’ where the latter was shown as the most susceptible. These data are lower than that reported by Amala et al. (2016) that recorded a Ro value of 125.43 in vine leaf disks in the Thompson Seedless variety.
In relation to the intrinsic rate of growth (rm); that is, the rate of population increase in the absence of a dense-dependent force shows that the variety ‛Cereza’ (0.3123) record (rm) greater than T. urticae than the others and therefore was considered more susceptible to this plague followed by ‛Saladette’ (0.3012), ‛Toro’ (0.2428) and ‛Tsan-101’ (0.1341). These results are similar to the values that oscillate between 0.254 and 0.313 in 5 cultivars of cucumber (Cucumis sativus L.) (Kheradpir et al., 2007). And greater than those recorded by Hoque et al. (2008) who registered in rm of 0.1873 in discs of bean leaves (Lablab purpureus L.).
Regarding the average time between generations (TG), the highest value was 8.48 days, with a daily increase of the population of 1.36 was recorded for ‛Cereza’ followed by the progressively shorter times in ‛Saladette’, ‛Toro’ and ‛Tsan-101’ (Table 2), these results imply a greater potential for damage in the variety ‛Cereza’ in comparison with the other three varieties and are inferior results to those reported by Riahi et al. (2011) on peach varieties where they registered a TG of 16.9 days and a daily growth factor of 1.07. Finally, the doubling time (T2) of the population of T. urticae in ‛Tsan-101’, ‛Toro’, ‛Saladette’ and ‛Cereza’ were 5.16, 2.85, 2.3 and 2.21 days, respectively, where Tsan -101 showed greater resistance for the development of the population followed by ‛Toro’, ‛Saladette’ and ‛Cereza’.
Conclusion
Of the varieties of tomato under study, Toro and Tsan 101 showed characteristics of resistance to T. urticae that changed the behavior and response towards the plants; the previous thing, places to these two varieties like suitable for the development of this crop in areas with high incidence of this plague, besides that they can be used in improvement programs once they are known which are the factors that affect in the defense of them.
Literatura citada
Abou, S. M. M. and Childers, C. C. 1987. A modified leaf arena technique for rearing phytoseiid or tetranychid mite for biological studies. Florida Entomol. 70(2):245-248. [ Links ]
Alba, J. M.; Montserrat M. and Fernández, M. R. 2009. Resistance to the two-spotted spider mite (Tetranychus urticae) by acylsucroses of wild tomato (Solanum pimpinellifolium) trichomes studied in a recombinant inbred line population. Exp. Appl. Acarol. 47(1):35-47. [ Links ]
Álvarez, S. J. H.; Cortina, G. H. A. y Villegas, M. J. F. 2001. Métodos para evaluar antibiosis a Hypothenemus hampei en café bajo condiciones controladas. Cenicafé. 52(3):2025-2034. [ Links ]
Amala, U.; Chinniah C.; Sawant I. S.; Yadav D. S. and Phad, D. M. 2016. Comparative biology and fertility parameters of two spotted spider mite, Tetranychus urticae Koch. On different grapevine varieties. VITIS-J. Grapevine Res. 55(1):31-36. [ Links ]
Birch, L. C. 1948. The intrinsic rate of natural increase of an insect population. J. Anim. Ecol. 17:15-26. [ Links ]
Desneux, N.; Decourtye, A. and Delpuech, J. M. 2007. The sublethal effects of pesticides on beneficial arthropods. Ann. Rev. Entomol. 52:81-106. [ Links ]
Flexner, j.; Westigar, P. H.; Hilton, R. and Croft, B. A. 1995. Experimental evaluation of resistance management for two-spotted spider mite (Acari: Tetranychidae) on Southern Oregon pear: 1987-1993. J. Econ. Entomol. 88:1517-1524. [ Links ]
Granados, G. y Paliwall, R. L. 2001. Mejoramiento para la resistencia a los insectos. In: maíz en los trópicos, mejoramiento y producción. Paliwall, R. L.; Granados, G.; Lafitte, H. R.; Violic, A. D. y Marathee, J. P. (Eds.). Organización de las naciones unidas para la agricultura y la alimentación (FAO). Colección FAO: producción y protección vegetal No. 28. Roma, Italia. 219-232 pp. [ Links ]
Hoque, M. F.; Islam, F. W. and Khalequzzaman, M. 2008. Life tables of two-spotted spider mite Tetranychus urticae Koch (Acari: Tetranychidae) and its predator Phytoseiulus persimilis Athias-Henriot (Acari: Phytoseiidae). J. Bio- Sci. 16:1-10. [ Links ]
Jansen, M. P. T. and Stamp, N. E. 1997. Effects of light availability on host plant chemistry and the consequences for behavior and growth of an insect herbivore. Entomol. Exp. Appl. 82:319-333. [ Links ]
Kawazu, K.; Mochizuki, A.; Sato, Y.; Sugeno, W.; Murat, M.; Seo, S. and Mitsuhara, I. 2012. Different expression profiles of jasmonic acid inducible genes in the tomato plant against herbivores with various feeding modes. Arthropod-Plant Interactions. 6:221-230. [ Links ]
Kheradpir, N.; Khalghani, J.; Ostovan, H. and Rezapanah, M. R. 2007. The comparison of demographic traits in Tetranychus urticae Koch (Acari: Tetranychidae) on five different greenhouse cucumber hybrids (Cucumis sativus). Acta Hort. 747:425-429. [ Links ]
Konanz, S. and Nauen, R. 2004. Purification and partial characterization of a glutathione S-transferase from the two-spotted spider mite, Tetranychus urticae. Pesticide Biochem. Physiol. 79:49-57. [ Links ]
Landeros, F. J.; Chávez, E. C.; Aguirre, L. A.; Canales, F. R. and Ochoa, Y. M. 2013. Demographic parameters of Tetranychus urticae (Acari: Tetranychidae) on four Rosa sp. Cultivars. Florida Entomologist. 96(4):1508-1512. [ Links ]
Riahi, E.; Nemati, A.; Shishehbor, P. and Saeidi, Z. 2011. Population growth parameters of the two-spotted spider mite, Tetranychus urticae, on three peach varieties in Irán. Acarologia. 51(4):473-480. [ Links ]
Saeidi, Z. and Mallik, B. 2012. Entrapment of two-spotted spider mite, Tetranychus urticae (Acari: Prostigmata: Tetranychidae), by type IV glandular trichomes of Lycopersicon species. J. Entomol. Soc. Irán. 31(2):15-27. [ Links ]
Sáenz de cabezón, F. J.; Martínez-Villar, E.; Moreno, F.; Marco, V. and Pérez, M. I. 2006. Influence of sublethal exposure to triflumuron on the biological performance of Tetranychus urticae Koch (Acari: Tetranychidae). Spanish J. Agric. Res. 4(2):167-172. [ Links ]
Smith, C. M. and Clement, S. L. 2012. Molecular bases of plant resistance to arthropods. Annu. Rev. Entomol. 57:309-328. [ Links ]
Santamaria, M. E.; Cambra, I.; Martinez, M.; Pozanco, C.; González, M. P.; Grbic, V.; Castañera, P.; Ortego, F. y Diaz, I. 2012. Gene pyramiding of peptidase inhibitors enhaces plant resistance to the pider mite Tetranychus urticae.Plos ONE 7(8): e43011. [ Links ]
Stumpf, N. and Nauen, R. 2001. Resistance mechanisms to mitocondrial electron transport inhibitors in a field-collected strain of Tetranychus urticae Koch (Acari: Tetranychidae). J. Econ. Entomol . 94:1577-1583. [ Links ]
Stumpf, N. and Nauen, R. 2002. Biochemical markers linked to abamectin resistance in Tetranychus urticae (Acari: Tetranychidae). Pestic. Biochem. Physiol. 72(2):111-121. [ Links ]
Takematsu, A. P.; Sulplicy, F. N.; Souza, F. M. F. e Sato M. E. 1994. Sensibilidade de Tetranychus urticae (Koch, 1836) proveniente de roseira (Rosa sp.) de Holambra-SP a alguns acaricidas. Rev. Agric. (Piracicaba) 69:129-137. [ Links ]
Tomczyk, A. and Kropczynska, D. 1985. Effects on the host plant. In: spider mites their biology, natural enemies and control. Word crop pest Helle, W. and Sabelis, M. W. (Eds.). Elsevier, Amsterdam. 317-329 pp. [ Links ]
Van den Boom, C.; Beek, T. V. and Dicke, M. 2003. Differences among plant species in acceptance by the spider mite Tetranychus urticae Koch. J. Appl. Entomol. 127(3):177-183. [ Links ]
Zehnder, G.; Gurr, G. M.; Kuhne, S.; Wade, M. R.; Wratten, S. D. and Wyss, E. 2007. Arthropod pest management in organic crops. Ann. Rev. Entomol . 52:57-80. [ Links ]
Received: January 2018; Accepted: April 2018
 Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
