











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
El tomate (Solanum lycopersicum L.), actualmente es una de las hortalizas de mayor importancia en muchos países del mundo por el gran número de subproductos que se obtienen de él y las divisas que aporta a su economía. En México se destinan aproximadamente 80 000 hectáreas de invernadero para su cultivo (Betancourt y Pierre, 2013) por lo que es obligado desarrollar o adaptar tecnologías que permitan mejorar su rendimiento y la calidad del fruto sin provocar efectos adversos al medio ambiente.
El uso de biorreguladores en la agricultura se ha intensificado en años recientes para mejorar la producción y calidad de frutos a través de diversas acciones que ofrece esta tecnología. En varias investigaciones se ha descubierto que la bioactividad de los biorreguladores está directamente relacionada con un mejor enlace del biorregulador al sitio de recepción en la célula y una mayor capacidad reactiva en el punto de inducción al estimular o inhibir un proceso fisiológico. Esta característica los ubica como una excelente tecnología para incrementar la calidad y producción en términos de maduración temprana o tardía de frutos, mayor contenido de antioxidantes, vida de anaquel, traslocación de nutrientes e incluso el combate de plagas y enfermedades (Nickell, 1988).
La prohexadiona de calcio (P-Ca) es un retardante de crecimiento, que actúa a través de la inhibición de la síntesis de giberelinas biológicamente activas (A1, A4, y A7) lo cual reduce el crecimiento vegetativo (Ramírez et al., 2016a). Se ha reportado en chile pimiento y cereza que P-Ca aumenta el contenido de citocininas en el meristemo apical, la cual está relacionada con la formación de flores y cuajado de los frutos (Ramírez et al., 2010). Efectos similares han sido reportados en especies frutales de clima templado como pera, durazno y manzana (Ramírez et al., 2003; Costa et al., 2004a; Costa et al., 2004b); sin embargo, poco se conoce sobre el efecto P-Ca, giberelinas AG4/7 y de la citocinina 6-bencil amino purina (6BAP) en el cultivo de tomate.
Por lo anterior, en este estudio se planteó conocer los efectos que producen la prohexadiona de calcio giberelinas AG4/7 y 6-bencilaminopurina en tomate (Solanum lycopersicum L.) bajo condiciones de invernadero.
Materiales y métodos
El estudio se realizó en la Universidad Autónoma Agraria Antonio Narro, Saltillo, Coahuila, México, en un invernadero con una estructura superior metálica cubierta con plástico blanco (calibre 720) en el techo y placas laterales de policarbonato. Como material vegetal se usaron plántulas de dos meses de tomate saladette de crecimiento indeterminado híbrido “Raptor-F1” las cuales fueron trasplantadas el 15 de abril de 2015 a bolsas de plástico negro con capacidad de 12 litros usando como sustrato: suelo, tezontle y perlita (2:1:2 v/v). Las bolsas fueron distribuidas a una distancia de 50 cm entre plantas y 75 cm entre filas, las condiciones climáticas dentro del invernadero se mantuvieron a 25 °C y 65% de humedad relativa durante el experimento. Se usó un sistema de riego por goteo de alto flujo en cada maceta, realizándose tres riegos diarios en diferentes horarios al día (9:00, 13:00 y 18:00 h), donde se aplicaron 900 ml de agua en cada maceta en cada riego efectuado, siendo esta cantidad el gasto por maceta al haberse efectuado una previa calibración. El cultivo se manejó a un tallo con las labores culturales tradicionales del departamento de horticultura en la UAAAN.
Los tratamientos con biorreguladores fueron los siguientes: control (agua), P -Ca (50 mg L-1), AG4/7 (50 mg L-1), AG4/7 (100 mg L-1), 6-BAP (50 mg L-1), 6-BAP (100 mg L-1), AG4/7 (50 mg L-1) + 6-BAP (50 mg L-1) y AG4/7 (100 mg L-1) + 6-BAP (100 mg L-1). Cuando las plantas mostraron los primeros primordios florales el 18 de mayo de 2015 se realizó la primera aplicación foliar a punto de rocío utilizando un atomizador y 15 días después se realizó la segunda aplicación con los mismos tratamientos. El experimento se estableció en un diseño completamente al azar con 10 repeticiones por tratamiento.
Las variables evaluadas fueron: temperatura de la hoja, contenido de clorofila, transpiración foliar, fotosíntesis, eficiencia intrínseca, rendimiento, contenido de vitamina C y licopeno en frutos, contenido de minerales en hojas y frutos (N, P, K, Ca, y Mg) y biomasa fresca y seca total. Los datos de resultados se evaluaron con el análisis de varianza (Anova) y una prueba de medias DMS (p≤ 0.05) mediante el uso del programa estadístico R versión 2.14.2 para Windows 8.1.
Parámetros foliares
La temperatura (°C) y clorofila (Unidades SPAD) en hojas fueron medidas de manera simultánea con un termómetro digital Thermometer IR marca Radioshack y un Medidor de Clorofila SPAD 502 marca Konica Minolta respectivamente. Se tomó como referencia la hoja más joven y mejor desarrollada en cada planta. Se realizaron tres mediciones después de cada aplicación cada 5 días. La transpiración foliar (mmol H2O m-2 s-1) y fotosíntesis (µmol CO2 m-2 s-1) fueron obtenidas con un Medidor Portátil LI-COR Modelo LI-6400 TX, en hojas descritas anteriormente.
Estas mediciones se realizaron en dos ocasiones, 5 días después de la primera aplicación y 37 días después de la segunda aspersión. Finalmente, la eficiencia intrínseca del uso de agua ((µmolCO2) (mmolH2O)-1) se obtuvo con la relación de fotosíntesis y transpiración foliar.
Rendimiento
El rendimiento total por planta resultó de la suma del peso de frutos cosechados en 10 cortes, utilizando una báscula marca Scout® Pro con una capacidad de 1 000 g.
Antioxidantes
El contenido de vitamina C en frutos se determinó usando el método reportado por Padayatt et al. (2001). Se maceró 10 g de pericarpio del fruto con 10 ml de ácido clorhídrico al 2% (v/v), posteriormente se homogeneizó la mezcla en 40 ml de agua destilada, se filtró a través de gasa y se colectó en un matraz Erlenmeyer. Se tomaron 10 ml de la solución y se titularon con 2.6- diclorofenolindofenol (1x10-3 N), hasta que la solución alcanzó un color rosa. El contenido de vitamina C se determinó utilizando la siguiente fórmula:
El contenido de licopeno se obtuvo de 3 g de peso fresco de pericarpio del fruto. Las muestras se colocaron en un mortero congelado que contenía 3 ml de amortiguador de fosfatos (pH 7) y se molió, de la mezcla obtenida se tomaron 2ml y se colocaron en tubos de centrífuga, se agregaron 4 ml de la mezcla hexano - acetona (3:2), se agitó la mezcla para separar y disolver los pigmentos de las membranas (Davis et al., 2003), se centrifugó a 3 000 rpm durante 10 min para la separación de fases, se extrajo la fase coloreada y se cuantifico a una longitud de onda de 450 nm en un equipo de HPLC marca Varian, modelo 500-MS. Para cuantificar el contenido de licopeno en las muestras, se construyó una curva de calibración de licopeno estándar (Sigma, Co) con un rango de 0-40 mg ml-1 previamente disuelto en la solución mencionada. Las muestras se compararon con la curva de calibración y el contenido de licopeno se determinó usando una ecuación de regresión lineal.
Contenido de minerales
Para el análisis de minerales (N, P, K, Mg y Ca) en frutos y hojas, se seleccionaron tres plantas al azar por cada tratamiento. El contenido de minerales en hojas se determinó al realizar tres muestreos en un intervalo de 20 días partiendo del inicio de la floración. La determinación de minerales en frutos se realizó en el primer corte cuando se tomaron 3 muestras al azar de las 10 repeticiones por tratamiento. El proceso de análisis de minerales correspondientes a Ca, Mg, K y P se realizó en dos etapas, la extracción y cuantificación. Para la extracción, las muestras fueron secadas en una estufa Modelo Felisa FE-291 a 70 °C durante 72 h, estas fueron molidas y un gramo de la muestra fue sometida al proceso de calcinación a 600 °C durante dos horas, las cenizas obtenidas se recuperaron con ácido clorhídrico 1:1 y se aforo a 100 m con agua destilada. En la segunda fase la concentración de los minerales Ca, Mg, y K (mg kg-1) se leyeron en un espectrofotómetro de absorción atómica Varían Spectrc AA5, mientras que la concentración de fosforo (mg kg-1). Se realizó obteniendo la absorbancia por fotocolorimetria con una longitud de onda de 560 nm. El nitrógeno (%) se determinó utilizando el procedimiento modificado de la digestión del micro-Kjeldahl (Jones, 1991).
Biomasa total
Para determinar la biomasa (g) se realizó un muestreo destructivo de tres plantas seleccionadas al azar en cada tratamiento. Se pesaron las muestras en una balanza Scout® Pro con una escala 0 - 1 000 g para obtener peso fresco, posteriormente las plantas fueron colocadas en bosas de papel estraza y puestas dentro de una estufa de secado marca Felisa FE 291 a 70 °C por cuatro días y luego se obtuvo el peso seco.
Resultados y discusión
Parámetros foliares
El efecto obtenido al aplicar biorreguladores sobre la temperatura de la hoja se muestra en la Figura 1. Los tratamientos individuales con giberelinas AG4/7 al compararse con el control, incrementaron significativamente la temperatura de la hoja (DMS p≤ 0.05), mientras que las aplicaciones con P-Ca y 6-BAP a 50 mg L-1 mostraron la menor temperatura en el tejido. Ferreyra et al. (2002) han relacionado la temperatura de la planta con la evolución del estado hídrico de la misma; efecto que ellos relacionan con las variaciones de aporte de agua a los tejidos. Goldhamer et al. (1999); Matthews et al. (1987) indican que la temperatura de la planta está relacionada con el estrés hídrico de la atmosfera adyacente a la misma.
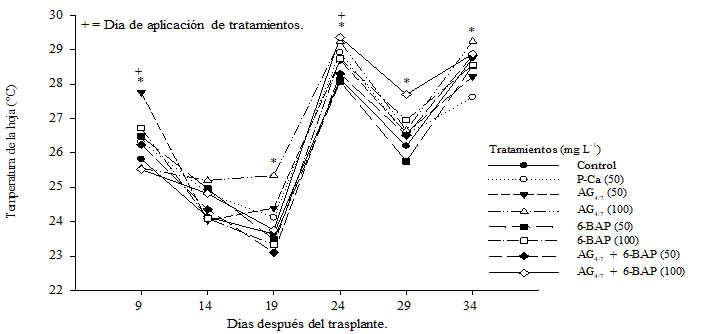
Figura 1 Efecto de biorreguladores en la temperatura de la hoja de plantas de tomate saladette híbrido “Raptor-F1”. Cada punto representa la media de diez repeticiones. *= diferencias estadísticas. DMS (p≤ 0.05).
Las giberelinas son hormonas que incrementan elongación celular originando un mayor flujo de agua hacia el tejido, condición que provocaría una mayor fluctuación de la temperatura de la hoja y manifestarse un déficit hídrico más rápido (Ramírez et al., 2016a) tal y como se puede apreciar en el Cuadro 1 donde la eficiencia hídrica ((µmolCO2) (mmolH2O)-1) que mostraron las plantas sometidas bajo tratamiento fueron menores al testigo en el primer muestreo. La P-Ca es un retardante vegetal que ocasiona en plantas un cierre parcial de estomas y por lo tanto una reducción en la pérdida de agua a través de la hoja (Rademacher, 2004) lo que ocasiona que esta hormona incremente ligeramente la temperatura de la hoja teniendo una eficiencia intrínseca del uso del agua ((µmolCO2) (mmolH2O)-1) (Cuadro 1) menor que el testigo.
Cuadro 1 Efecto de biorreguladores en la fotosíntesis, transpiración foliar y eficiencia hídrica en plantas de tomate saladette híbrido “Raptor-F1”.
| Tratamientos (mg L-1) | Fotosíntesis (µmol CO2 m-2 s-1) |
Transpiración foliar (mmolH2Om-2 s-1) |
Eficiencia intrínseca (µmolCO2) (mmolH2O)-1 |
|||||
| 5 DD1A | 37 DD2A | 5 DD1A | 37 DD2A | 5 DD1A | 37 DD2A | |||
| Control | 24.1006 a | 11.0341 ab | 9.4949 c | 5.3013 a | 2.5279 a | 2.184 b | ||
| P-Ca(50) | 23.9093 a | 8.22962 b | 10.1735 abc | 3.9729 ab | 2.3569 ab | 2.5308 b | ||
| AG4/7(50) | 21.6628 ab | 10.3904 ab | 10.0333 abc | 2.274 b | 2.1703 bc | 5.2571 a | ||
| AG4/7(100) | 20.006 ab | 11.5683 ab | 10.1711 abc | 4.0577 ab | 1.9596 cd | 2.94 b | ||
| 6-BAP(50) | 21.9897 ab | 10.8346 ab | 10.7223 ab | 4.9765 a | 2.0441 c | 2.1835 b | ||
| 6-BAP(100) | 16.7876 b | 12.5993 a | 9.589 bc | 4.0472 ab | 1.7597 d | 3.149 b | ||
| AG4/7+6-BAP(50) | 23.7023 a | 10.9025 ab | 11.1604 a | 3.7085 ab | 2.135 bc | 3.4587 b | ||
| AG4/7+6-BAP(100) | 19.3155 ab | 10.1388 ab | 9.9116 a | 3.8445 ab | 1.9542 cd | 3.1022 b | ||
| CV (%) | 14.89 | 21.68 | 9.71 | 38.64 | 11.5 | 48.67 | ||
| SE | * | * | * | * | * | * | ||
Valores con la misma letra en cada columna son estadísticamente iguales de acuerdo con la prueba DMS; *= diferencias significativas a p≤ 0.05; SE= significancia estadística; CV= coeficiente de variación; DD1A= días después de la primera aplicación. DD2A= días después de la segunda aplicación. Cada valor es el promedio de seis plantas.
El contenido de clorofila en las hojas de todos los tratamientos con biorreguladores fué menor que el control (Figura 2). Los niveles de clorofila en las hojas tratadas con P-Ca estuvieron cercanas al control. Bekheta et al. (2009) demostraron en plantas de haba que P-Ca a 10, 20 y 30 mg L-1 aumentaron el contenido de pigmentos. Fletcher y Hofestra (1985) propusieron que la aplicación óptima de biorreguladores causan mejorias en diversas plantas vegetales como el aumento de niveles de clorofila y una amplia gama de cloroplastos. Los autores realizarón sus estudios con plantas a campo abierto. El presente estudió fue realizado en invernadero y con diferentes concentraciones de biorreguladores. La menor cantidad de clorofila observada entre ellos pudiera reflejar el concepto de concentración supraóptima reportada en otros cultivos (Rademacher, 2000).
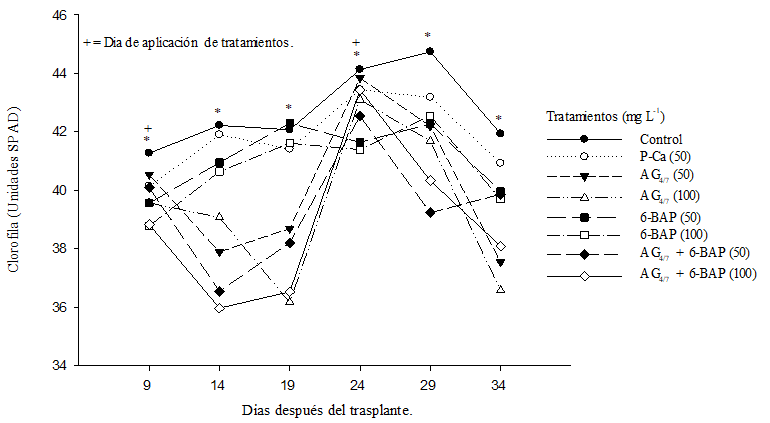
Figura 2 Efecto de biorreguladores en el contenido de clorofila de plantas de tomate saladette híbrido “Raptor-F1”. Cada punto representa la media de diez repeticiones; *= diferencias estadísticas (DMS p≤ 0.05).
Es importante señalar, que la clorofila es un factor de crecimiento exponencial en la planta que a menudo adopta una curva que crece a inicios del cultivo pero que suele disminuir al finalizar el ciclo de produccion (Zermeño et al., 2015). Este comportamiento natural se modificó al aplicar AG4/7 a 50 y 100 mg L-1 y con la combinacion de AG4/7 y 6-BAP a 50 y 100 mg L-1. Se puede observar que días despues de las aplicaciones, el verdor de las hojas (representadas en unidades SPAD) se reduce de forma imediata y logró recuperarse 15 días despues de la aplicación (Figura 2). P-Ca sigue un patron natural de produccion de clorofila, lo cual podría explicarse por su capacidad para inhibir la formación de giberelinas biologicamente activas (Ramírez et al., 2003; Ramírez et al., 2016a). Las plantas asperjadas con 6-BAP a concentraciones de 50 y 100 mg L-1 mostraron valores menores a P-Ca; sin embargo, el comportamiento de la curva de clorofila no se vió afectada, esto concuerda con lo reportado por Ros et al. (2004) quienes observaron estas mismas tendencias al realizar aplicaciones de 6-BAP en un cultivo de algodón.
En el Cuadro 1 se presentan los efectos de biorreguladores sobre la fotosintesis (µmol CO2 m-2 s-1), transpiración foliar (mmol H2O m-2 s-1) y la eficiencia intrínseca del uso del agua ((µmolCO2) (mmolH2O)-1) en hojas. Se observó, en la mayoría de los tratamientos con biorreguladores que no modificaron adversamente esos procesos fisiológicos y por lo tanto pueden ser considerados muy positivos para futuros estudios que permitan ampliar más el conocimiento sobre fotosíntesis y conductancia estomática en tomate bajo condiciones de invernadero.
Biomasa total
En la Figura 3 se presenta la biomasa fresca y seca total de los tratamientos. P-Ca y 6-BAP a 50 mg L-1, fueron similares (p≤ 0.05) al control en la variable peso fresco total (PFT). En previas investigaciones P-Ca en dósis superiores a 100 mg L-1 incrementó el numero de hojas, diametro del tallo y numero de entrenudos asi como la biomasa fresca y seca de la planta (Ramírez et al., 2016a). En manzano Golden Delicious también fue observada esta relación (Ramírez et al., 2003). Esto evidencia el hecho de que P-Ca es un bloqueador de sintesis de giberelinas biologicamente activas (Rademacher, 2004) que actua como retardante de crecimiento apical y estimula la síntesis de citocininas originando el incremento vegetativo de otros organos de la planta (Costa, et al., 2004a). Esa experiencia llevaría a considerar en el presente estudio el no efecto debido a baja dósis de esos biorreguladores empleados en tomate.
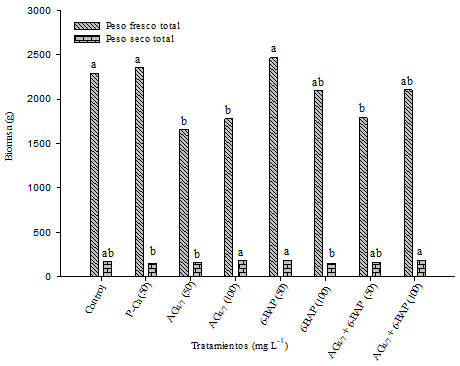
Figura 3 Influencia de biorreguladores en la biomasa fresca y seca total en plantas de tomate saladette híbrido “Raptor-F1”. Cada barra representa el promedio de diez repeticiones. Barras con la misma letra son estadísticamente iguales (DMS p≤ 0.05).
En peso seco total (PST) (Figura 3) se observo una difereciencia estadisticamente significativa (p≤ 0.05) entre los tratamientos donde AG4/7 + 6-BAP mostraron ser efectivos al aumentar la materia seca de las plantas. Estos efectos han sido observados en cacao por Cárdenas-Hernández et al. (2010) quienes evaluaron el efecto del ácido giberélico y la 6-bencilaminopurina sobre el desarrollo de yemas en injertos de esa especie. A nivel general estos resultados apoyan el postulado de que las giberelinas y 6-BAP promueven la elongación celular, aumentando el crecimiento de la planta (El Fouly et al., 1988; Wilson-García et al., 2007).
La aplicación de cualquier biorregulador no alteró el contenido de N, Mg y Ca en hojas (Cuadro 2). De acuerdo a Gutiérrez (1997) el flujo de minerales en plantas es variado ya que depende de la disponibilidad y de la demanda entre órganos, así como la etapa fenológica de la misma; sin embargo, estos elementos son requeridos para el metabolismo de la planta ya que cumplen funciones estructurales en las moléculas orgánicas, en la reserva energética e iónica y reacciones redox (Aven et al., 1992). En base a esas experiencias, los efectos en la fisiología del tomate observados en este estudio (Cuadro 2) son interesantes al no observar detrimento en los niveles de nutrientes.
Cuadro 2 Efecto de biorreguladores en el contenido de minerales en hojas de plantas de tomate saladette híbrido “Raptor-F1”.
| Tratamientos (mg L-1) | N (%) | P (mg kg-1) | K (mg kg-1) | Mg (mg kg-1) | Ca (mg kg-1) |
| Control | 3.052 a | 2562.7451 a | 24466.66 ab | 4383.33 a | 36933.33 a |
| P-Ca(50) | 3.2351 a | 2669.4117 a | 21800 b | 4370 a | 39800 a |
| AG4/7(50) | 3.3266 a | 2563.5294 a | 21233.33 b | 4316.66 a | 38000 a |
| AG4/7(100) | 3.2656 a | 2447.4509 ab | 22400 b | 4366.66 a | 36066.66 a |
| 6-BAP(50) | 3.0825 a | 2561.1764 a | 25100 ab | 4340 a | 35733.33 a |
| 6-BAP(100) | 3.0825 a | 2463.9215 ab | 23600 ab | 4363.33 a | 36566.66 a |
| AG4/7+6-BAP(50) | 3.113 a | 2502.3529 ab | 27800 a | 4420 a | 35266.66 a |
| AG4/7+6-BAP(100) | 3.1435 a | 1960.3921 a | 26800 a | 4406.66 a | 34500 a |
| CV | 8.5004 | 20.7393 | 15.4744 | 2.2418 | 14.1219 |
| SE | ns | * | * | ns | ns |
Valores con la misma letra en cada columna son estadísticamente iguales de acuerdo con la prueba DMS; *= diferencias significativas a una p≤ 0.05; NS= diferencias no significativas a una P≤ 0.05; SE= significancia estadística; CV= coeficiente de variación. Cada valor representa el promedio de 6 plantas.
En la mayoría de los tratamientos, el contenido de fosforo fue similar al control; mientras que, el contenido de potasio fue mayor en los tratamientos AG4/7 + 6-BAP a cualquier dosis (Cuadro 2). La eficiencia de los biorreguladores en la nutrición vegetal ha sido relacionada con la translocación de nutrientes hacia determinados tejidos (Medjdoub et al., 2002; Rademacher, 2004) en donde se puede causar un estímulo o inhibición de su aparición o desarrollo (Ramírez et al., 2010).
El Cuadro 3 muestra los niveles de nitrógeno acumulados en el fruto. Los tratamientos AG4/7 a 50mg L-1 y AG4/7+ 6-BAP a 100 mg L-1 presentaron una mayor cantidad de este elemento superando al control en 2.1 y 8% respectivamente. Se ha demostrado que el nitrógeno está relacionado directamente con el contenido proteico y es también un promotor de la síntesis y producción de azúcar en los frutos (Kjellbom y Larsson, 1984). Un estudio realizado por Ramírez et al. (2016a) se demostró que AG4/7 a 100 mg L-1 incremento el contenido de grados brix y firmeza y prolongó la vida de anaquel en los frutos. Lo anterior es relevante puesto que a mayor contenido de azúcar en el fruto existe la posibilidad de una menor vida de anaquel (Siller-Cepeda et al., 2004).
Cuadro 3 Efecto de biorreguladores en el contenido de minerales en frutos de tomate saladette híbrido “Raptor-F1”.
| Tratamientos (mg L-1) | N (%) | P (mg kg-1) | K (mg kg-1) | Mg (mg kg-1) | Ca (mg kg-1) |
| Control | 2.7468 ab | 2169.0196 abc | 41233.33 a | 1966.66 a | 2200 abc |
| P-Ca (50) | 2.6857 ab | 2273.3333 ab | 38133.33 a | 1860 ab | 2300 ab |
| AG4/7 (50) | 2.8078 a | 2053.7254 bc | 35166.66 a | 1686.66 b | 1933.33 bc |
| AG4/7 (100) | 2.5636 ab | 2374.5098 a | 37733.33 a | 1770 ab | 2233.33 ab |
| 6-BAP (50) | 2.5942 ab | 1986.2745 c | 36000 a | 1720 ab | 2366.66 a |
| 6-BAP (100) | 2.3805 b | 2380 a | 38300 a | 1856.66 ab | 2100 abc |
| AG4/7+6-BAP (50) | 2.7468 ab | 2237.2549 ab | 39800 a | 1803.33 ab | 1833.33 c |
| AG4/7+6-BAP (100) | 2.9604 a | 2283.5294 ab | 38600 a | 1790 ab | 2066.66 abc |
| CV (%) | 12.8088 | 9.4362 | 14.1087 | 9.6691 | 15.7478 |
| SE | * | * | ns | * | * |
Valores con la misma letra en cada columna son estadísticamente iguales de acuerdo con la prueba DMS; *= diferencias significativas a una p≤ 0.05; SE= significancia estadística; ns= diferencias no significativas a una p≤ 0.05; CV= coeficiente de variación, cada valor representa el promedio de seis plantas.
El contenido de fósforo en frutos varió entre tratamientos (Cuadro 3). Los biorreguladores AG4/7 y 6-BAP a 100 mg L-1 mostraron incrementos de 9.4 y 9.7% respectivamente con relación al control. Gutiérrez (1997) demostró que el fósforo es traslocado en mayor cantidad a los frutos cuando estos inician su desarrollo. El contenido de potasio no se vio afectado por los biorreguladores y mantuvieron niveles similares al control.
Ramírez et al. (2010) demostraron que P-Ca no afecta los niveles de K en frutos de tomate de cascara. Barrera et al. (2008) reportó que el contenido de potasio en la planta oscila entre 2.7% y 4.5%, influye en la fotosíntesis, transporte de los carbohidratos y desempeña un papel importante como elemento antagónico del nitrógeno en frutos, además de que regula la entrada y el metabolismo de los nutrimentos. En medicina humana el potasio ocupa el tercer puesto dentro de los minerales que más actúan en nuestro organismo y desempeña un papel importante en la mayoría de las funciones vitales (Hopkinson et al., 1998); sin embargo, Hobson y Davies (1980) indican que una alta concentración de potasio en fruto se asocia a una baja calidad del mismo.
Un ligero y menor contenido de magnesio en frutos resultó en la mayoría de los tratamientos con biorreguladores (Cuadro 3). Es posible que esta variación sea provocada por la concentración de hormonas utilizada y por el estado fisiológico del momento en el desarrollo-maduración del fruto. Gutiérrez (1997), menciona que el flujo de Mg en plantas es variado y depende de la disponibilidad y demanda entre órganos de la planta y la etapa de desarrollo de la misma. Por otro lado, Betancourt y Pierre (2013) señalan que el orden de extracción del Mg en tomate es de hoja > tallo > fruto > raíz, haciendo posible demostrar que la cantidad de Mg que se acumula en frutos depende de la cantidad almacenada en tallos y hojas.
Esto se refleja al comparar el Mg hallado en hojas (Cuadro 2) donde las plantas asperjadas con los biorreguladores fueron similares al control, condición que pudo provocar un envío desequilibrado de Mg hacia los frutos. Lo anterior, reflejaría la mayor cantidad de Mg observada en hojas al compararse con frutos. Estos datos coinciden con los publicados por Fayad et al. (2002) quienes reportaron una mayor acumulación de Mg en la parte vegetativa del tomate con relación al fruto.
Se puede observar que el contenido de calcio fue mayor en el tratamiento 6-BAP a 50 mg L-1 (Cuadro 3) mientras que P-Ca a 50 mg L-1 también mostró una tendencia mayor al control. Betancourt y Pierre (2013) observó que el nivel de Ca acumulado en frutos de tomate fue gradual partiendo de hojas > tallos y que generalmente la acumulación en frutos suele ser muy baja, esto presumiblemente debido a que el Ca tiene poca movilidad en el floema, transportándose en la planta básicamente a través del xilema (Malone et al., 2002). Es importante considerar entonces, que el alto nivel de Ca con 6-BAP y P-Ca al 50 mg L-1 resalta el potencial de estos biorregulares como buenas alternativas para incrementar la calidad en el fruto de tomate.
Rendimiento
La Figura 4 muestra que el rendimiento en los tratamientos 6-BAP y P-Ca en 50 mg L-1 fue similar al testigo. Este comportamiento podría deberse al efecto concentración. Previas investigaciones (Ramírez et al., 2010) reportan que la P-Ca a 200 mg L-1 incremento el rendimiento 83% en tomate de cascara. Esta influencia que ha tenido P-Ca en aumentar el rendimiento, también se obtuvo en pera (Costa et al., 2004a) y manzano (Greene, 1996; Unrath, 1999; Basak y Rademacher, 2000). El resto de los tratamientos tuvieron una tendencia a tener un rendimiento menor que el control. Este efecto puede reflejar mayor estímulo a desarrollo vegetativo el cual compitió con la formación floral y por lo tanto menos rendimiento (Rademacher, 2004).

Antioxidantes
El efecto de los tratamientos sobre el contenido de licopeno y vitamina C en frutos de tomate se muestra en el Cuadro 4. La combinación de AG4/7 + 6-BAP a 100 mg L-1 originó aumentos significativos en ambos antioxidantes; estos superaron al control en 55.4% y 47.8% en licopeno y vitamina C respectivamente. Las concentraciones individuales de AG4/7 mostraron mayor contenido de los dos antioxidantes; mientras que P-Ca también reflejo ese efecto para vitamina C.
Cuadro 4 Efecto de biorreguladores en el contenido de licopeno y vitamina C en frutos de tomate saladette híbrido “Raptor-F1”.
| Tratamientos (mg L-1) | Licopeno (mg L-1) | Vitamina C (mg 100 g-1) |
| Control | 3.2343 abc | 14.485 b |
| P-Ca(50) | 3.834 abc | 15.625 ab |
| AG4/7(50) | 3.9233 ab | 19.8 ab |
| AG4/7(100) | 2.618 bc | 20.26 ab |
| 6-BAP(50) | 2.6376 bc | 14.465 b |
| 6-BAP(100) | 1.7523 c | 14.555 b |
| AG4/7+6-BAP(50) | 4.118 ab | 20.61 ab |
| AG4/7+6-BAP(100) | 5.0263 a | 21.41 a |
| CV (%) | 66.4166 | 29.8391 |
| SE | * | * |
Valores con la misma letra en cada columna son estadísticamente iguales de acuerdo con la prueba DMS; *= diferencias significativas a una p≤ 0.05; CV= coeficiente de variación; SE= significancia estadística, cada valor representa el promedio de 10 plantas.
Previas investigaciones demuestran que P-Ca incrementa el contenido de vitamina C y licopeno en tomate rojo (Ramírez et al., 2003) y chile habanero (Ramírez et al., 2016b). Estos hallazgos permitieron catalogar a P-Ca como un retardante que incrementa el contenido de antioxidantes lo cual repercute en una buena salud en los seres humanos ya que fortalece el sistema inmunológico que da protección contra enfermedades como la diabetes, el cáncer y la presión arterial (Ramírez et al., 2010). Poca información existe sobre el estímulo de giberelinas y citocininas en la síntesis de licopeno y vitamina C en tomate (Ramírez et al., 2010). Por lo tanto, más investigación es necesaria sobre este tema.
El mayor contenido en licopeno y vitamina C observado en varios tratamientos con biorreguladores otorga un valor agregado al fruto. Esta característica permite un precio mayor en el mercado internacional que el de un tomate con niveles normales de esos antioxidantes (Ramírez et al., 2016a), lo que compensa un rendimiento igual o quizás menor al del control.
Conclusiones
En tomate saladette híbrido “Raptor-F1” cultivado en invernadero, los biorreguladores P-Ca y 6-BAP a 50 mg L-1 no alteran la fisiología foliar y rendimiento, incrementan el nivel de potasio en hojas y de nitrógeno y calcio en frutos. 6-BAP a 50 mg L-1 incrementa la materia fresca y seca de la planta; al combinarse con las giberelinas AG4/7 a 100 mg L-1 aumentan el contenido de vitamina C y licopeno en frutos.

